Species Composition and Toxigenic Potential of Fusarium Isolates Causing Fruit Rot of Sweet Pepper in China

 ,
, 

Abstract
:1. Introduction
2. Results and Discussion
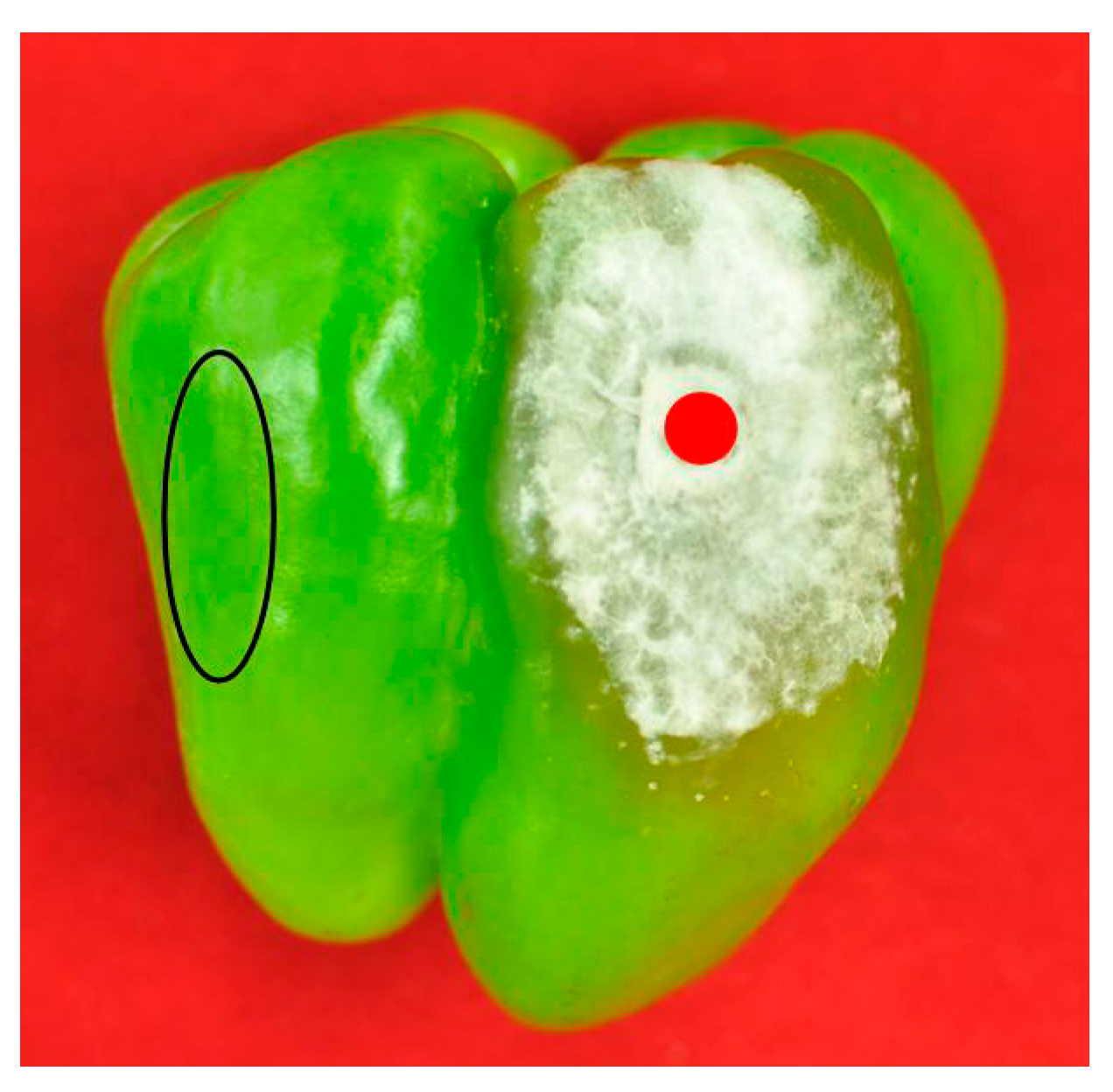
2.1. Isolation and Identification of Fusarium Isolates
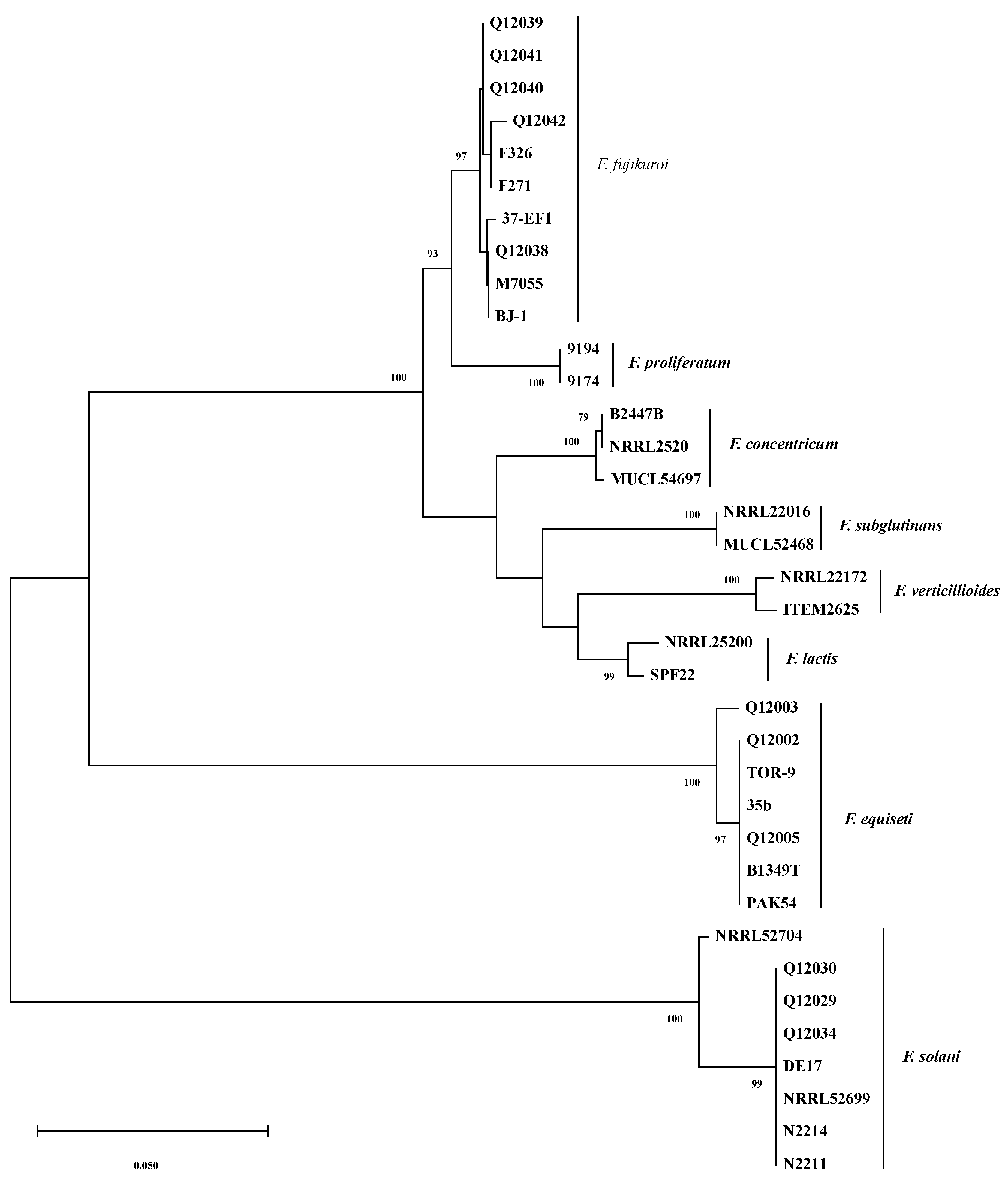
2.2. Molecular Phylogenetics
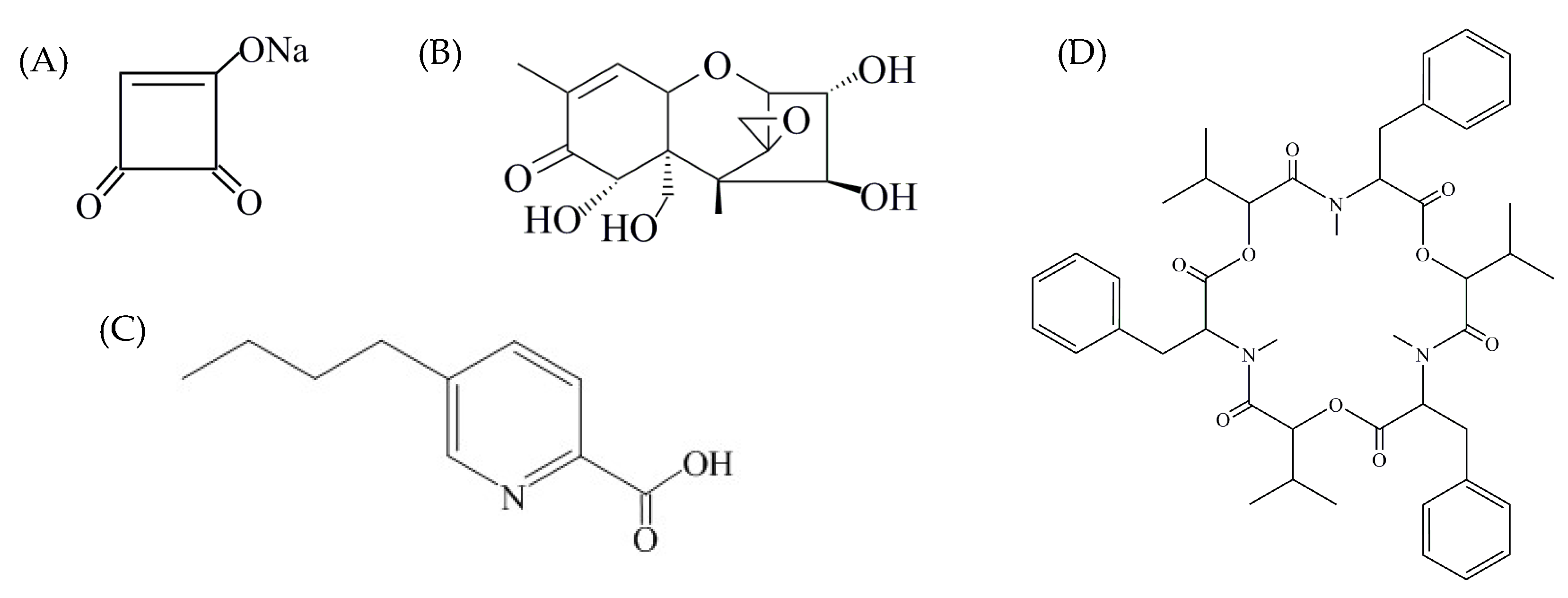
2.3. Multi-Mycotoxin Analysis in Pepper Fruit
3. Conclusions
4. Materials and Methods
4.1. Isolation and Purification of Fusarium Strains
4.2. Nucleotide Maniplation
4.3. Fusarium Strain Identification
4.4. Phylogenetic Analysis
4.5. Mycotoxin Production Analysis in Pepper Fruits via LC-MS/MS
Author Contributions
Funding
Conflicts of Interest
References
- Vanderslice, J.T.; Higgs, D.J.; Hayes, J.M.; Block, G. Ascorbic acid and dehydroascorbic acid content of foods-as-eaten. J. Food Compos. Anal. 1990, 3, 105–118. [Google Scholar] [CrossRef]
- Speranza, G.; Lo Scalzo, R.; Morelli, C.F.; Rabuffetti, M.; Bianchi, G. Influence of drying techniques and growing location on the chemical composition of sweet pepper (Capsicum annuum L., var. Senise). J. Food Biochem. 2019, e13031. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Yu, C.S.; Shen, Y.; Fang, X.D.; Chen, L.; Min, J.M.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed]
- Van Poucke, K.; Monbaliu, S.; Munaut, F.; Heungens, K.; De Saeger, S.; Van Hove, F. Genetic diversity and mycotoxin production of Fusarium lactis species complex isolates from sweet pepper. Int. J. Food Microbiol. 2012, 153, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.W.; Hong, S.K.; Kim, W.G.; Lee, Y.K. First report of internal fruit rot of sweet pepper in Korea caused by Fusarium lactis. Plant Dis. 2011, 95, 1476. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Bouras, N.; Yang, J.; Howard, R.J.; Strelkov, S.E. Mycotoxin production by isolates of Fusarium lactis from greenhouse sweet pepper (Capsicum annuum). Int. J. Food Microbiol. 2011, 151, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.T. Fusarium stem and fruit rot of sweet peppers in the glasshouse. Plant Pathol. 1994, 43, 225–227. [Google Scholar] [CrossRef]
- Yang, J.; Kharbanda, P.D.; Howard, R.J.; Mirza, M. Identification and pathogenicity of Fusarium lactis, causal agent of internal fruit rot of greenhouse sweet pepper in Alberta. Can. J. Plant Pathol. 2009, 31, 47–56. [Google Scholar] [CrossRef]
- Howard, R.J.; Garland, J.A.; Seaman, W.L. Disease and Pests of Vegetable Crops in Canada, 1st ed.; The Canadian Phytopathological Society and Entomological Society of Canada: Ottawa, ON, Canada, 1994; pp. 333–336. [Google Scholar]
- Yang, Y.; Cao, T.; Yang, J.; Howard, R.J.; Kharbanda, P.D.; Strelkov, S.E. Histopathology of internal fruit rot of sweet pepper caused by Fusarium lactis. Can. J. Plant Pathol. 2010, 32, 86–97. [Google Scholar] [CrossRef]
- Utkhede, R.; Mathur, S. Fusarium fruit rot of greenhouse sweet peppers in Canada. Plant Dis. 2003, 87, 100. [Google Scholar] [CrossRef]
- Utkhede, R.; Mathur, S. Internal fruit rot caused by Fusarium subglutinans in greenhouse sweet peppers. Can. J. Plant Pathol. 2004, 26, 386–390. [Google Scholar] [CrossRef]
- Jarvis, W.R.; Khosla, S.K.; Barrie, S.D. Fusarium stem and fruit rot of sweet pepper in Ontario greenhouses. Can. Plant Dis. Surv. 1994, 74, 131–134. [Google Scholar]
- Frans, M.; Aerts, R.; Van Laethem, S.; Ceusters, J. Environmental effects on growth and sporulation of Fusarium spp. causing internal fruit rot in bell pepper. Eur. J. Plant Pathol. 2017, 149, 875–883. [Google Scholar] [CrossRef]
- Kharbanda, P.D.; Yang, J.; Howard, R.J.; Mirza, M. Internal fruit rot of greenhouse peppers caused by Fusarium lactis-a new disease. Greenh. Bus. 2006, 5, 11–16. [Google Scholar]
- Chu, F.S.; Liu, G.Y. Simultaneous occurrence of fumonisin B1 and other mycotoxins in moldy corn collected from People’s Republic of China in regions with high incidences of esophageal cancer. Appl. Environ. Microb. 1994, 60, 847–852. [Google Scholar]
- Peraica, M.; Radic, B.; Lucic, A.; Pavlovic, M. Toxic effects of mycotoxins in humans. Bull. World Health Organ. 1999, 77, 754–766. [Google Scholar] [PubMed]
- Yazar, S.; Omurtag, G.Z. Fumonisins, trichothecenes and zearalenone in cereals. Int. J. Mol. Sci. 2008, 9, 2062–2090. [Google Scholar] [CrossRef] [PubMed]
- Monbaliu, S.; Van Poucke, K.; Heungens, K.; Van Peteghem, C.; De Saeger, S. Production and migration of mycotoxins in sweet pepper analyzed by multimycotoxin LC-MS/MS. J. Agric. Food Chem. 2010, 58, 10475–10479. [Google Scholar] [CrossRef]
- Restani, P. Diffusion of mycotoxins in fruits and vegetables. In Mycotoxins in Fruits and Vegetables, 1st ed.; Barkai-Golan, R., Paster, N., Eds.; Academic Press: San Diego, CA, USA, 2008; pp. 105–114. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Lab Manual, 1st ed.; Blackwell Publishing: Ames, IA, USA, 2006; pp. 3–274. [Google Scholar]
- Wang, J.H.; Feng, Z.H.; Han, Z.; Song, S.Q.; Lin, S.H.; Wu, A.B. First report of pepper fruit rot caused by Fusarium concentricum in China. Plant Dis. 2013, 97, 1657. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from unclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- O’Donnell, K.; Humber, R.A.; Geiser, D.M.; Kang, S.; Park, B.; Robert, V.A.; Crous, P.W.; Johnston, P.R.; Aoki, T.; Rooney, A.P.; et al. Phylogenetic diversity of insecticolous fusaria inferred from multilocus DNA sequence data and their molecular identification via FUSARIUM-ID and Fusarium MLST. Mycologia 2012, 104, 427–445. [Google Scholar] [CrossRef] [PubMed]
- Scauflaire, J.; Gourgue, M.; Munaut, F. Fusarium temperatum sp. nov. from maize, an emergent species closely related to Fusarium subglutinans. Mycologia 2011, 103, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Ward, T.J.; Bielawski, J.P.; Kistler, H.C.; Sullivan, E.; O’Donnell, K. Ancestral polymorphism and adaptive evolution in the trichothecene mycotoxin gene cluster of phytopathogenic Fusarium. Proc. Natl. Acad. Sci. USA 2002, 9, 9278–9283. [Google Scholar] [CrossRef] [PubMed]
- Jennings, P.; Coates, M.E.; Turner, J.A.; Chandler, E.A.; Nicholson, P. Determination of deoxynivalenol and nivalenol chemotypes of Fusarium culmorum isolates from England and Wales by PCR assay. Plant Pathol. 2004, 53, 182–190. [Google Scholar] [CrossRef]
- Yang, L.; van der Lee, T.; Yang, X.; Yu, D.; Waalwijk, C. Fusarium populations on Chinese barley show a dramatic gradient in mycotoxin profiles. Phytopathology 2008, 98, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Al-Hatmi, A.M.S.; Mirabolfathy, M.; Hagen, F.; Normand, A.; Stielow, J.B.; Karami-Osbo, R.; Van Diepeningen, A.D.; Meis, J.F.; De Hoog, G.S. DNA barcoding, MALDI-TOF, and AFLP data support Fusarium ficicrescens as a distinct species within the Fusarium fujikuroi species complex. Fungal Biol. 2016, 120, 265–278. [Google Scholar] [CrossRef]
- Wulff, E.G.; Sorense, J.L.; Lubeck, M.; Nielsen, K.F.; Thrane, U.; Torp, J. Fusarium spp. associated with rice bakanae: Ecology, genetic diversity, pathogenicity and toxigenicity. Environ. Microbiol. 2010, 12, 649–657. [Google Scholar] [CrossRef]
- Stepien, L.; Koczyk, G.; Waskiewicz, A. FUM cluster divergence in fumonisins-producing Fusarium species. Fungal Biol. 2011, 115, 112–123. [Google Scholar] [CrossRef]
- Hestbjerg, H.; Nielsen, K.F.; Thrane, U.; Elmholt, S. Production of trichothecenes and other secondary metabolites by Fusarium culmorum and Fusarium equiseti on common laboratory media and a soil organic matter agar: An ecological interpretation. J. Agric. Food Chem. 2002, 50, 7593–7599. [Google Scholar] [CrossRef]
- Stankovic, S.; Levic, J.; Petrovic, T.; Logrieco, A.; Moretti, A. Pathogenicity and mycotoxin production by Fusarium proliferatum isolated from onion and garlic in Serbia. Eur. J. Plant Pathol. 2007, 118, 165–172. [Google Scholar] [CrossRef]
- Wang, Q.G.; Xu, L.J. Beauvericin, a bioactive compound produced by fungi: A short review. Molecules 2012, 17, 2367–2377. [Google Scholar] [CrossRef] [PubMed]
- Qu, B.; Li, H.P.; Zhang, J.B.; Xu, Y.B.; Huang, T.; Wu, A.B.; Zhao, C.S.; Carter, J.; Nicholson, P.; Liao, Y.C. Geographic distribution and genetic diversity of Fusarium graminearum and F. asiaticum on wheat spikes throughout China. Plant Pathol. 2008, 57, 15–24. [Google Scholar] [CrossRef]
- Wang, J.H.; Li, H.P.; Qu, B.; Zhang, J.B.; Huang, T.; Chen, F.F.; Liao, Y.C. Development of a generic PCR detection of 3-acetyldeoxynivalenol, 15-acetyldeoxynivalenol- and nivalenol-chemotypes of Fusarium graminearum clade. Int. J. Mol. Sci. 2008, 9, 2495–2504. [Google Scholar] [CrossRef]
- Abd Murad, N.B.; Mohamed Nor, N.N.; Shohaimi, S.; Mohd Zainudin, N.A.I. Genetic diversity and pathogenicity of Fusarium species associated with fruit rot disease in banana across Peninsular Malaysia. J. Appl. Microbiol. 2017, 123, 1533–1546. [Google Scholar] [CrossRef]
- Pastrana, A.M.; Basallote-Ureba, M.J.; Aquado, A.; Capote, N. Potential inoculum sources and incidence of strawberry soilborne pathogens in Spain. Plant Dis. 2017, 101, 751–760. [Google Scholar] [CrossRef]
- Wang, J.H.; Zhang, J.B.; Li, H.P.; Gong, A.D.; Xue, S.; Agboola, R.S.; Liao, Y.C. Molecular identification, mycotoxin production and comparative pathogenictiy of Fusarium temperatum isolated from maize in China. J. Phytopathol. 2013. [Google Scholar] [CrossRef]
- Lima, C.S.; Pfenning, L.H.; Costa, S.S.; Abreu, L.M.; Leslie, J.F. Fusarium tupiense sp. nov., a member of the Gibberella fujikuroi complex that causes mango malformation in Brazil. Mycologia 2012, 104, 1408–1419. [Google Scholar] [CrossRef]
- Masratul Hawa, M.; Salleh, B.; Latiffah, Z. Characterization and pathogenicity of Fusarium proliferatum causing stem rot of Hylocereus polyrhizus in Malaysia. Ann. Appl. Biol. 2013, 163, 269–280. [Google Scholar] [CrossRef]
- Skovgaard, K.; Rosendahl, S.; O’Donnell, K.; Nirenberg, H.I. Fusarium commune is a new species identified by morphological and molecular phylogenetic data. Mycologia 2003, 95, 630–636. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Monbaliu, S.; Van Poucke, C.; Van Peteghem, C.; Van Poucke, K.; Heungens, K.; De Saeger, S. Development of a multi-mycotoxin liquid chromatography/tandem mass spectrometry method for sweet pepper analysis. Rapid Commun. Mass Spectrom. 2009, 23, 3–11. [Google Scholar] [CrossRef]
- Zhao, Z.Y.; Yang, X.L.; Zhao, X.Y.; Chen, L.; Bai, B.; Zhou, C.Y.; Wang, J.H. Method development and validation for the analysis of emerging and traditional fusarium mycotoxins in pepper, potato, tomato, and cucumber by UPLC-MS/MS. Food Anal. Methods 2018, 11, 1780–1788. [Google Scholar] [CrossRef]

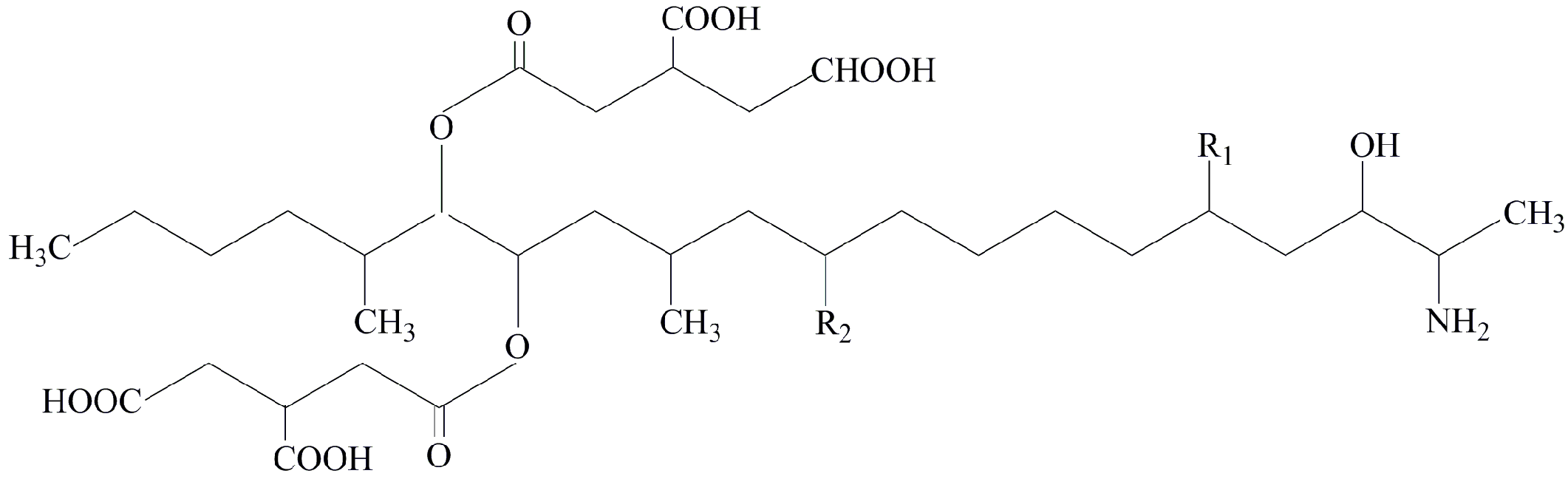
| Figure | R1 | R2 |
|---|---|---|
| Fumonisin B1 | OH | OH |
| Fumonisin B2 | OH | H |
| Fumonisin B3 | H | OH |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain Code | Origin | Symptom (External/Internal) | GenBank Accession No. |
|---|---|---|---|---|
| F. equiseti | Q12001 | Shanghai | external | - |
| Q12002 | Shanghai | internal | KF208617 | |
| Q12003 | Shanghai | external | KF208618 | |
| Q12004 | Shanghai | external | - | |
| Q12005 | Shanghai | internal | KF208619 | |
| Q12006 | Shanghai | external | - | |
| Q12007 | Shanghai | external | - | |
| Q12008 | Shanghai | internal | - | |
| Q12009 | Shanghai | external | - | |
| Q12010 | Hainan | external | - | |
| Q12011 | Shandong | external | - | |
| Q12012 | Shandong | external | - | |
| Q12013 | Shanghai | external | - | |
| Q12014 | Shanghai | external | - | |
| Q12015 | Shandong | external | - | |
| Q12016 | Helongjiang | external | - | |
| Q12017 | Zhejiang | external | - | |
| Q12018 | Zhejiang | internal | - | |
| Q12019 | Zhejiang | external | - | |
| Q12020 | Zhejiang | external | - | |
| Q12021 | Zhejiang | external | - | |
| Q12022 | Zhejiang | internal | - | |
| Q12023 | Zhejiang | external | - | |
| Q12024 | Zhejiang | external | - | |
| Q12025 | Hunan | external | - | |
| Q12026 | Hunan | external | - | |
| Q12027 | Hunan | external | - | |
| F. solani | Q12028 | Shanghai | external | - |
| Q12029 | Shanghai | external | KF208620 | |
| Q12030 | Shanghai | external | KF208621 | |
| Q12031 | Shanghai | external | - | |
| Q12032 | Shanghai | external | - | |
| Q12033 | Shanghai | external | - | |
| Q12034 | Shanghai | internal | KF208622 | |
| Q12035 | Shanghai | internal | - | |
| Q12036 | Shanghai | external | - | |
| Q12037 | Shanghai | external | - | |
| F. fujikuroi | Q12038 | Hainan | external | KF208623 |
| Q12039 | Zhejiang | external | KF208624 | |
| Q12040 | Zhejiang | external | KF208625 | |
| Q12041 | Hunan | internal | KF208626 | |
| Q12042 | Hunan | external | KF208627 |
| Fusarium Species | Strain Code | Position 1 | Mycotoxins Detected (μg/kg) | |||||
|---|---|---|---|---|---|---|---|---|
| BEA | FA | FB1 | FB2 | MON | NIV | |||
| F. equiseti | Q12002 | IS | 5.20 | 411.64 | - | - | - | 184.16 |
| HS | - | 13.80 | - | - | - | - | ||
| F. equiseti | Q12004 | IS | 9.12 | 160.32 | - | - | - | - |
| HS | - | - | - | - | - | - | ||
| F. solani | Q12029 | IS | - | 41.44 | - | - | - | - |
| HS | - | - | - | - | - | - | ||
| F. solani | Q12034 | IS | 98.48 | 78.98 | - | - | - | - |
| HS | - | - | - | - | - | - | ||
| F. solani | Q12037 | IS | - | - | - | - | - | - |
| HS | - | - | - | - | - | - | ||
| F. fujikuroi | Q12038 | IS | 153.84 | 10662.36 | 1512.48 | 1190.12 | 2439.48 | - |
| HS | - | 592.00 | 45.36 | 31.67 | 193.75 | - | ||
| F. fujikuroi | Q12039 | IS | 318.00 | 4870.56 | 1559.04 | 851.88 | 1429.76 | - |
| HS | 130.40 | 41.08 | 24.30 | 172.13 | - | |||
| F. fujikuroi | Q12040 | IS | 44.44 | 6522.72 | 43.64 | 26.96 | 151.72 | - |
| HS | - | 874.04 | - | - | - | - | ||
| F. fujikuroi | Q12041 | IS | 23.80 | 2940.72 | 3926.60 | 3734.16 | 1925.00 | - |
| HS | - | 530.48 | 183.04 | 144.92 | 253.12 | - | ||
| F. concentricum | MUCL54697 | IS | 1019.60 | 2886.08 | - | - | 35.80 | - |
| HS | - | 55.64 | - | - | - | - | ||
| Speices | Strain code | Origin | Host/Substrate | Accession number | Reference |
|---|---|---|---|---|---|
| F. concentricum | B2447B | Malaysia | Banana | KY379181 | [38] |
| F. concentricum | MUCL54697 | China | Sweet pepper | KC816735 | [22] |
| F. concentricum | NRRL25202 | Korea | Delphacidae | JF740760 | [24] |
| F. equiseti | 35b | China | Wheat | KY466715 | - |
| F. equiseti | B1349T | China | Tomato | KM886212 | - |
| F. equiseti | PAK54 | USA | Loquat | KY523101 | - |
| F. equiseti | Q12002 | China | Sweet pepper | KF208617 | This study |
| F. equiseti | Q12003 | China | Sweet pepper | KF208618 | This study |
| F. equiseti | Q12005 | China | Sweet pepper | KF208619 | This study |
| F. equiseti | TOR-9 | Spain | Strawberry | KX215087 | [39] |
| F. fujikuroi | 37-EF1 | Malaysia | Pineapple | KC584844 | - |
| F. fujikuroi | BJ-1 | China | Bletilla striata | MH263736 | - |
| F. fujikuroi | F271 | India | Rice | KM586385 | - |
| F. fujikuroi | F326 | India | Rice | KP009955 | - |
| F. fujikuroi | M7055 | China | Maize | KC964126 | [40] |
| F. fujikuroi | Q12038 | China | Sweet pepper | KF208623 | This study |
| F. fujikuroi | Q12039 | China | Sweet pepper | KF208624 | This study |
| F. fujikuroi | Q12040 | China | Sweet pepper | KF208625 | This study |
| F. fujikuroi | Q12041 | China | Sweet pepper | KF208626 | This study |
| F. fujikuroi | Q12042 | China | Sweet pepper | KF208627 | This study |
| F. lactisi | NRRL25200 | USA | Ficus carica | AF160272 | [41] |
| F. lactisi | SPF22 | Korea | Sweet pepper | JF411959 | [5] |
| F. proliferatum | 9174 | Malaysia | Pitaya | JX869021 | [42] |
| F. proliferatum | 9194 | Malaysia | Pitaya | JX869025 | [42] |
| F. solani | DE17 | Malaysia | Mangrove soil | KM096385 | - |
| F. solani | N2211 | Malaysia | Cucurbits | KT211623 | - |
| F. solani | N2214 | Malaysia | Cucurbits | KT211624 | - |
| F. solani | NRRL52699 | Colombia | Cercopidae | JF740782 | [24] |
| F. solani | NRRL52704 | USA | Tetranychidae | JF740786 | [24] |
| F. solani | Q12029 | China | Sweet pepper | KF208620 | This study |
| F. solani | Q12030 | China | Sweet pepper | KF208621 | This study |
| F. solani | Q12034 | China | Sweet pepper | KF208622 | This study |
| F. subglutinans | MUCL52468 | Belgium | Maize | HM067691 | [25] |
| F. subglutinans | NRRL22016 | USA | Maize | AF160289 | [25,41,43] |
| F. verticillioides | ITEM2625 | Slovakia | Maize | KF715264 | - |
| F. verticillioides | NRRL22172 | Germany | Maize | AF160262 | [25,41,43] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Wang, S.; Zhao, Z.; Lin, S.; Van Hove, F.; Wu, A. Species Composition and Toxigenic Potential of Fusarium Isolates Causing Fruit Rot of Sweet Pepper in China. Toxins 2019, 11, 690. https://doi.org/10.3390/toxins11120690
Wang J, Wang S, Zhao Z, Lin S, Van Hove F, Wu A. Species Composition and Toxigenic Potential of Fusarium Isolates Causing Fruit Rot of Sweet Pepper in China. Toxins. 2019; 11(12):690. https://doi.org/10.3390/toxins11120690
Chicago/Turabian StyleWang, Jianhua, Shuangxia Wang, Zhiyong Zhao, Shanhai Lin, François Van Hove, and Aibo Wu. 2019. "Species Composition and Toxigenic Potential of Fusarium Isolates Causing Fruit Rot of Sweet Pepper in China" Toxins 11, no. 12: 690. https://doi.org/10.3390/toxins11120690
APA StyleWang, J., Wang, S., Zhao, Z., Lin, S., Van Hove, F., & Wu, A. (2019). Species Composition and Toxigenic Potential of Fusarium Isolates Causing Fruit Rot of Sweet Pepper in China. Toxins, 11(12), 690. https://doi.org/10.3390/toxins11120690