Malaysian Cobra Venom: A Potential Source of Anti-Cancer Therapeutic Agents


Abstract
:1. Introduction
2. Malaysian Common Cobras
3. Proteomic Composition of the Venom from N. kaouthia, N. sumatrana, and O. hannah
4. Potential AntiCancer Activity of Malaysian Cobra Venom
4.1. Ophiophagus Hannah
4.2. Naja Kaouthia
4.3. Naja Sumatrana
5. Future Directions and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Omar, Z.A.; Ibrahim Tamin, N. National Cancer Registry Report: Malaysia Cancer Statistics-Data and Figure; National Cancer Registry: Putrajaya, Malaysia, 2011; pp. 85–87. [Google Scholar]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.-W. Cancer and radiation therapy: Current advances and future directions. Int. J. Med. Sci. 2012, 9, 193. [Google Scholar] [CrossRef] [PubMed]
- Jain, D.; Kumar, S. Snake venom: A potent anticancer agent. Asian Pac. J. Cancer Prev. 2012, 13, 4855–4860. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Dong, C.; Li, X.; Han, W.; Su, X. Anticancer potential of bioactive peptides from animal sources (review). Oncol. Rep. 2017, 38, 637–651. [Google Scholar] [CrossRef] [PubMed]
- Liberio, M.S.; Joanitti, G.A.; Fontes, W.; Castro, M.S. Anticancer peptides and proteins: A panoramic view. Protein Pept. Lett. 2013, 20, 380–391. [Google Scholar] [PubMed]
- Mustaffa, F.; Indurkar, J.; Ali, N.; Hanapi, A.; Shah, M.; Ismail, S.; Mansor, S. A review of malaysian medicinal plants with potential antidiabetic activity. J. Pharm. Res. 2011, 4, 4217–4224. [Google Scholar]
- Tan, K.Y.; Tan, C.H.; Fung, S.Y.; Tan, N.H. Venomics, lethality and neutralization of Naja kaouthia (monocled cobra) venoms from three different geographical regions of Southeast Asia. J. Proteome 2015, 120, 105–125. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.-J.; Rim, G.S.; Yang, H.I.; Yin, C.S.; Koh, H.G.; Jang, M.-H.; Kim, C.-J.; Choe, B.-K.; Chung, J.-H. Bee venom induces apoptosis through caspase-3 activation in synovial fibroblasts of patients with rheumatoid arthritis. Toxicon 2005, 46, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-D.; Kim, S.-Y.; Kim, T.-W.; Lee, S.-H.; Yang, H.-I.; Lee, D.-I.; Lee, Y.-H. Anti-inflammatory effect of bee venom on type II collagen-induced arthritis. Am. J. Chin. Med. 2004, 32, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-Y.; Shim, S.-R.; Rhee, H.Y.; Park, H.-J.; Jung, W.-S.; Moon, S.-K.; Park, J.-M.; Ko, C.-N.; Cho, K.-H.; Park, S.-U. Effectiveness of acupuncture and bee venom acupuncture in idiopathic Parkinson’s Disease. Parkinsonism Relat. Disord. 2012, 18, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-I.; Yang, E.J.; Lee, M.S.; Kim, Y.-S.; Huh, Y.; Cho, I.-H.; Kang, S.; Koh, H.-K. Bee venom reduces neuroinflammation in the MPTP-induced model of Parkinson’s Disease. Int. J. Neurosci. 2011, 121, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.D.; Debnath, A.; Saha, A.; Giri, B.; Tripathi, G.; Vedasiromoni, J.R.; Gomes, A.; Gomes, A. Indian black scorpion (Heterometrus bengalensis Koch.) venom induced antiproliferative and apoptogenic activity against human leukemic cell lines U937 and K562. Leuk. Res. 2007, 31, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lyu, P.; Xi, X.; Ge, L.; Mahadevappa, R.; Shaw, C.; Kwok, H.F. Triggering of cancer cell cycle arrest by a novel scorpion venom-derived peptide—Gonearrestide. J. Cell Mol. Med. 2018, 22, 4460–4473. [Google Scholar] [CrossRef] [PubMed]
- Cushman, D.W.; Ondetti, M.A. History of the design of captopril and related inhibitors of angiotensin converting enzyme. Hypertension 1991, 17, 589–592. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S. A bradykinin-potentiating factor (BPF) present in the venom of Bothrops jararaca. Br. J. Pharmacol. 1965, 24, 163–169. [Google Scholar] [CrossRef]
- Pope, J.E.; Deer, T.R. Ziconotide: A clinical update and pharmacologic review. Expert Opin. Pharmacother. 2013, 14, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Koh, C.Y.; Kini, R.M. From snake venom toxins to therapeutics—Cardiovascular examples. Toxicon 2012, 59, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.L.; Wu, B.J.; Kao, P.H.; Fu, Y.S.; Chang, L.S. Antibacterial and membrane-damaging activities of β-bungarotoxin b chain. J. Pept. Sci. 2013, 19, 1–8. [Google Scholar] [CrossRef]
- Muller, V.D.; Russo, R.R.; Cintra, A.C.; Sartim, M.A.; Alves-Paiva Rde, M.; Figueiredo, L.T.; Sampaio, S.V.; Aquino, V.H. Crotoxin and phospholipases A2 from Crotalus durissus terrificus showed antiviral activity against dengue and yellow fever viruses. Toxicon 2012, 59, 507–515. [Google Scholar] [CrossRef]
- Castillo, J.C.Q.; Vargas, L.J.; Segura, C.; Gutiérrez, J.M.; Pérez, J.C.A. In vitro antiplasmodial activity of phospholipases A2 and a phospholipase homologue isolated from the venom of the snake Bothrops asper. Toxins 2012, 4, 1500–1516. [Google Scholar] [CrossRef]
- Yamane, E.S.; Bizerra, F.C.; Oliveira, E.B.; Moreira, J.T.; Rajabi, M.; Nunes, G.L.; de Souza, A.O.; da Silva, I.D.; Yamane, T.; Karpel, R.L. Unraveling the antifungal activity of a South American rattlesnake toxin crotamine. Biochimie 2013, 95, 231–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderon, L.A.; Sobrinho, J.C.; Zaqueo, K.D.; de Moura, A.A.; Grabner, A.N.; Mazzi, M.V.; Marcussi, S.; Nomizo, A.; Fernandes, C.F.; Zuliani, J.P.; et al. Antitumoral activity of snake venom proteins: New trends in cancer therapy. BioMed Res. Int. 2014, 2014, 203639. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, J.; Lin, Y. Snake venoms in cancer therapy: Past, present and future. Toxins 2018, 10, 346. [Google Scholar] [CrossRef] [PubMed]
- Vyas, V.K.; Brahmbhatt, K.; Bhatt, H.; Parmar, U. Therapeutic potential of snake venom in cancer therapy: Current perspectives. Asian Pac. J. Trop. Biomed. 2013, 3, 156–162. [Google Scholar] [CrossRef]
- Das, I.; Ahmed, N.; Liat, L.B. Venomous terrestrial snakes of Malaysia: Their identity and biology. Clin. Toxicol. 2013, 1–15. [Google Scholar] [CrossRef]
- Tweedie, M.W.F. The Snakes of Malaya, 3rd ed.; Singapore National Printers: Singapore, 1983; 167p. [Google Scholar]
- Lee, M.L.; Tan, N.H.; Fung, S.Y.; Sekaran, S.D. Antibacterial action of a heat-stable form of L-amino acid oxidase isolated from king cobra (Ophiophagus hannah) venom. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Phua, C.; Vejayan, J.; Ambu, S.; Ponnudurai, G.; Gorajana, A. Purification and antibacterial activities of an L-amino acid oxidase from king cobra (Ophiophagus hannah) venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 198–207. [Google Scholar] [CrossRef]
- Ahn, M.Y.; Lee, B.M.; Kim, Y.S. Characterization and cytotoxicity of L-amino acid oxidase from the venom of king cobra (Ophiophagus hannah). Int. J. Biochem. Cell Biol. 1997, 29, 911–919. [Google Scholar] [CrossRef]
- Fung, S.Y.; Lee, M.L.; Tan, N.H. Molecular mechanism of cell death induced by king cobra (Ophiophagus hannah) venom L-amino acid oxidase. Toxicon 2015, 96, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.L.; Fung, S.Y.; Chung, I.; Pailoor, J.; Cheah, S.H.; Tan, N.H. King cobra (Ophiophagus hannah) venom L-amino acid oxidase induces apoptosis in PC-3 cells and suppresses PC-3 solid tumor growth in a tumor xenograft mouse model. Int. J. Med. Sci. 2014, 11, 593. [Google Scholar] [CrossRef] [PubMed]
- Li Lee, M.; Chung, I.; Yee Fung, S.; Kanthimathi, M.S.; Hong Tan, N. Antiproliferative activity of king cobra (Ophiophagus hannah) venom L-amino acid oxidase. Basic Clin. Pharmacol. Toxicol. 2014, 114, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Gomes, A.; Chakravarty, A.; Biswas, A.; Giri, B.; Dasgupta, S. CNS and anticonvulsant activity of a non-protein toxin (KC-MMTX) isolated from king cobra (Ophiophagus hannah) venom. Toxicon 2006, 47, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Du, X.-Y.; Clemetson, J.M.; Navdaev, A.; Magnenat, E.M.; Wells, T.N.; Clemetson, K.J. Ophioluxin, a convulxin-like C-type lectin from Ophiophagus hannah (king cobra) is a powerful platelet activator via glycoprotein VI. J. Biol. Chem. 2002, 277, 35124–35132. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Zhao, H.Y.; Yin, Y.; Shen, S.S.; Shan, L.L.; Chen, C.X.; Zhang, Y.X.; Gao, J.F.; Ji, X. Combined venomics, antivenomics and venom gland transcriptome analysis of the monocoled cobra (Naja kaouthia) from China. J. Proteom. 2017, 159, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Kulkeaw, K.; Chaicumpa, W.; Sakolvaree, Y.; Tongtawe, P.; Tapchaisri, P. Proteome and immunome of the venom of the Thai cobra, Naja kaouthia. Toxicon 2007, 49, 1026–1041. [Google Scholar] [CrossRef] [PubMed]
- Yap, M.K.K.; Fung, S.Y.; Tan, K.Y.; Tan, N.H. Proteomic characterization of venom of the medically important southeast asian Naja sumatrana (equatorial spitting cobra). Acta Trop. 2014, 133, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Petras, D.; Heiss, P.; Sussmuth, R.D.; Calvete, J.J. Venom proteomics of Indonesian king cobra, Ophiophagus hannah: Integrating top-down and bottom-up approaches. J. Proteome Res. 2015, 14, 2539–2556. [Google Scholar] [CrossRef] [PubMed]
- Izidoro, L.F.M.; Sobrinho, J.C.; Mendes, M.M.; Costa, T.R.; Grabner, A.N.; Rodrigues, V.M.; da Silva, S.L.; Zanchi, F.B.; Zuliani, J.P.; Fernandes, C.F. Snake venom L-amino acid oxidases: Trends in pharmacology and biochemistry. BioMed Res. Int. 2014, 2014, 196754. [Google Scholar] [CrossRef] [PubMed]
- Augusto-de-Oliveira, C.S.; Stuginski, D.R.; Kitano, E.S.; Andrade-Silva, D.b.; Liberato, T.; Fukushima, I.; Serrano, S.M.; Zelanis, A. Dynamic rearrangement in snake venom gland proteome: Insights into Bothrops jararaca intraspecific venom variation. J. Proteome Res. 2016, 15, 3752–3762. [Google Scholar] [CrossRef]
- Shanbhag, V.K.L. Applications of snake venoms in treatment of cancer. Asian Pac. J. Trop. Biomed. 2015, 5, 275–276. [Google Scholar] [CrossRef] [Green Version]
- Essex, H.E.; Priestley, J.T. Effect of rattlesnake venom on Flexner-Jobling’s carcinoma in the white rat (Mus norvegicus Albinus.). Proc. Soc. Exp. Biol. Med. 1931, 28, 550–551. [Google Scholar] [CrossRef]
- Kurotchkin, T.; Spies, J. Effects of cobra venom on the Fujinami rat sarcoma. Proc. Soc. Exp. Biol. Med. 1935, 32, 1408–1410. [Google Scholar] [CrossRef]
- Kerkkamp, H.; Bagowski, C.; Kool, J.; van Soolingen, B.; Vonk, F.J.; Vlecken, D. Whole snake venoms: Cytotoxic, anti-metastatic and antiangiogenic properties. Toxicon 2018, 150, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Fung, S.Y.; Lee, M.L.; Tan, N.H. Proteomic investigation of the molecular mechanism of king cobra venom L-amino acid oxidase induced apoptosis of human breast cancer (MCF-7) cell line. Indian J. Exp. Biol. 2018, 56, 101–111. [Google Scholar]
- Debnath, A.; Chatterjee, U.; Das, M.; Vedasiromoni, J.R.; Gomes, A. Venom of Indian monocellate cobra and Russell’s viper show anticancer activity in experimental models. J. Ethnopharmacol. 2007, 111, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Feofanov, A.V.; Sharonov, G.V.; Astapova, M.V.; Rodionov, D.I.; Utkin, Y.N.; Arseniev, A.S. Cancer cell injury by cytotoxins from cobra venom is mediated through lysosomal damage. Biochem. J. 2005, 390, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Debnath, A.; Saha, A.; Gomes, A.; Biswas, S.; Chakrabarti, P.; Giri, B.; Biswas, A.K.; Gupta, S.D.; Gomes, A. A lethal cardiotoxic-cytotoxic protein from the Indian monocellate cobra (Naja kaouthia) venom. Toxicon 2010, 56, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Lipps, B.V. Novel snake venom proteins cytolytic to cancer cells in vitro and in vivo systems. J. Venom. Anim. Toxins Incl. Trop. Dis. 1999, 5, 172–183. [Google Scholar] [CrossRef]
- Zainal Abidin, S.A.; Rajadurai, P.; Hoque Chowdhury, M.E.; Othman, I.; Naidu, R. Cytotoxic, Anti-Proliferative and Apoptosis Activity of L-Amino Acid Oxidase from Malaysian Cryptelytrops purpureomaculatus (CP-LAAO) Venom on Human Colon Cancer Cells. Molecules 2018, 23, 1388. [Google Scholar] [CrossRef] [PubMed]
- Zainal Abidin, S.A.; Rajadurai, P.; Chowdhury, M.E.H.; Ahmad Rusmili, M.R.; Othman, I.; Naidu, R. Cytotoxic, Antiproliferative and Apoptosis-inducing Activity of L-Amino Acid Oxidase from Malaysian Calloselasma rhodostoma on Human Colon Cancer Cells. Basic Clin. Pharmacol. Toxicol. 2018, 123, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Mauro, V.P.; Adriana, S.P.; Andreimar, M.S.; Juliana, P.Z. An Update on Potential Molecular Mechanisms Underlying the Actions of Snake Venom L-amino Acid Oxidases (LAAOs). Curr. Med. Chem. 2018, 25, 2520–2530. [Google Scholar]
- Zhang, L.; Wei, L.J. ACTX-8, a cytotoxic L-amino acid oxidase isolated from Agkistrodon acutus snake venom, induces apoptosis in Hela cervical cancer cells. Life Sci. 2007, 80, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, W.T. Isolation and characterization of ACTX-6: A cytotoxic L-amino acid oxidase from Agkistrodon acutus snake venom. Nat. Prod. Res. 2008, 22, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Wang, J.H.; Lee, W.H.; Wang, Q.; Liu, H.; Zheng, Y.T.; Zhang, Y. Molecular characterization of Trimeresurus stejnegeri venom L-amino acid oxidase with potential anti-HIV activity. Biochem. Biophys. Res. Commun. 2003, 309, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Liu, S.; Dong, P.; Zhao, D.; Wang, C.; Tao, Z.; Sun, M.Z. Akbu-LAAO exhibits potent anti-tumor activity to HepG2 cells partially through produced H2O2 via TGF-beta signal pathway. Sci. Rep. 2015, 5, 18215. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, P.H.; Zuliani, J.P.; Fernandes, C.F.; Calderon, L.A.; Stabeli, R.G.; Nomizo, A.; Soares, A.M. Mechanism of the cytotoxic effect of L-amino acid oxidase isolated from Bothrops alternatus snake venom. Int. J. Biol. Macromol. 2016, 92, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Suhr, S.M.; Kim, D.S. Identification of the snake venom substance that induces apoptosis. Biochem. Biophys. Res. Commun. 1996, 224, 134–139. [Google Scholar] [CrossRef]
- Rodrigues, R.S.; Izidoro, L.F.; de Oliveira, R.J., Jr.; Sampaio, S.V.; Soares, A.M.; Rodrigues, V.M. Snake venom phospholipases A2: A new class of antitumor agents. Protein Pept. Lett. 2009, 16, 894–898. [Google Scholar] [CrossRef]
- Zouari-Kessentini, R.; Luis, J.; Karray, A.; Kallech-Ziri, O.; Srairi-Abid, N.; Bazaa, A.; Loret, E.; Bezzine, S.; El Ayeb, M.; Marrakchi, N. Two purified and characterized phospholipases A2 from Cerastes cerastes venom, that inhibit cancerous cell adhesion and migration. Toxicon 2009, 53, 444–453. [Google Scholar] [CrossRef]
- Khunsap, S.; Pakmanee, N.; Khow, O.; Chanhome, L.; Sitprija, V.; Suntravat, M.; Lucena, S.E.; Perez, J.C.; Sanchez, E.E. Purification of a phospholipase A2 from Daboia russelii siamensis venom with anticancer effects. J. Venom. Res. 2011, 2, 42–51. [Google Scholar]
- Liang, Y.J.; Yang, X.P.; Wei, J.W.; Fu, L.W.; Jiang, X.Y.; Chen, S.W.; Yang, W.L. correlation of antitumor effect of recombinant sea snake basic phospholipase A2 to its enzymatic activity. Ai Zheng 2005, 24, 1474–1478. [Google Scholar] [PubMed]
- Chwetzoff, S.; Tsunasawa, S.; Sakiyama, F.; Menez, A. Nigexine, a phospholipase A2 from cobra venom with cytotoxic properties not related to esterase activity. Purification, amino acid sequence, and biological properties. J. Biol. Chem. 1989, 264, 13289–13297. [Google Scholar] [PubMed]
- Rudrammaji, L.M.; Gowda, T.V. Purification and characterization of three acidic, cytotoxic phospholipases A2 from Indian cobra (Naja naja naja) venom. Toxicon 1998, 36, 921–932. [Google Scholar] [CrossRef]
- Premzl, A.; Kovacic, L.; Halassy, B.; Krizaj, I. Generation of ammodytoxin-anti-cathepsin B immuno-conjugate as a model for delivery of secretory phospholipase A2 into cancer cells. Toxicon 2008, 51, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Maity, G.; Mandal, S.; Chatterjee, A.; Bhattacharyya, D. Purification and characterization of a low molecular weight multifunctional cytotoxic phospholipase A2 from Russell’s viper venom. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2007, 845, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Sales, T.A.; Marcussi, S.; Da Cunha, E.F.F.; Kuca, K.; Ramalho, T.C. Can inhibitors of snake venom phospholipases A2 lead to new insights into anti-inflammatory therapy in humans? A theoretical study. Toxins 2017, 9, 341. [Google Scholar] [CrossRef] [PubMed]
- Quach, N.D.; Arnold, R.D.; Cummings, B.S. Secretory phospholipase A2 enzymes as pharmacological targets for treatment of disease. Biochem. Pharmacol. 2014, 90, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.M.; Chang, S.Y.; Lin, K.L.; Chiu, C.C.; Chang, L.S.; Lin, S.R. Taiwan cobra cardiotoxin III inhibits SRC kinase leading to apoptosis and cell cycle arrest of oral squamous cell carcinoma CA9-22 cells. Toxicon 2010, 56, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.L.; Su, J.C.; Chien, C.M.; Chuang, P.W.; Chang, L.S.; Lin, S.R. Down-regulation of the JAK2/PI3k-mediated signaling activation is involved in taiwan cobra cardiotoxin III-induced apoptosis of human breast MDA-MB-231 cancer cells. Toxicon 2010, 55, 1263–1273. [Google Scholar] [CrossRef]
- Chen, K.C.; Lin, S.R.; Chang, L.S. Involvement of mitochondrial alteration and reactive oxygen species generation in Taiwan cobra cardiotoxin-induced apoptotic death of human neuroblastoma SK-N-SH cells. Toxicon 2008, 52, 361–368. [Google Scholar] [CrossRef]
- Paleari, L.; Negri, E.; Catassi, A.; Cilli, M.; Servent, D.; D’Angelillo, R.; Cesario, A.; Russo, P.; Fini, M. Inhibition of non-neuronal α7-nicotinic receptor for lung cancer treatment. Am. J. Respir. Crit. Care Med. 2009, 179, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, E.; Gurrola, G.B.; Schwartz, E.F.; Possani, L.D. Scorpion venom components as potential candidates for drug development. Toxicon 2015, 93, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Mahadevappa, R.; Kwok, H.F. Venom-based peptide therapy: Insights into anti-cancer mechanism. Oncotarget 2017, 8, 100908–100930. [Google Scholar] [CrossRef] [PubMed]
- Mahadevappa, R.; Ma, R.; Kwok, H.F. Venom peptides: Improving specificity in cancer therapy. Trends Cancer 2017, 3, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadoon, M.K.; Abdel-Maksoud, M.A.; Rabah, D.M.; Badr, G. Induction of apoptosis and growth arrest in human breast carcinoma cells by a snake (Walterinnesia aegyptia) venom combined with silica nanoparticles: Crosstalk between Bcl2 and caspase 3. Cell Physiol. Biochem. 2012, 30, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadoon, M.K.; Rabah, D.M.; Badr, G. Enhanced anticancer efficacy of snake venom combined with silica nanoparticles in a murine model of human multiple myeloma: Molecular targets for cell cycle arrest and apoptosis induction. Cell. Immunol. 2013, 284, 129–138. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Naja kaouthia | Naja sumatrana | Ophiophagus hannah | |
|---|---|---|---|
| Common name | Monocled cobra | Equatorial spitting cobra | King cobra |
| Characteristics | Absence of occipitals, brown to greyish-brown body, with white circle hood mark | Absence of occipitals, black body, without hood mark, white marking on throat | Large head; small hood; adult has yellow, green, brown, or black body; presence of a pair of occipitals behind parietals |
| Length | Usually 4–5 feet, occasionally can reach up to 7.5 feet | Usually 3–3.9 feet, occasionally can reach up to 4.9 feet | Usually 8–18 feet |
| Distribution in Malaysia | Peninsular Malaysia, mainly in the northern part of peninsular Malaysia | Peninsular Malaysia, Sabah, and Sarawak | Peninsular Malaysia, Sabah, and Sarawak |
| Habitat | Not habitat-specific, can adapt to a wide range of habitats such as grassland and paddy fields | Not habitat-specific, can adapt to a wide range of habitats such as primary and secondary forests and human-surrounding environments | Habitat-specific, mainly inhabits forests |
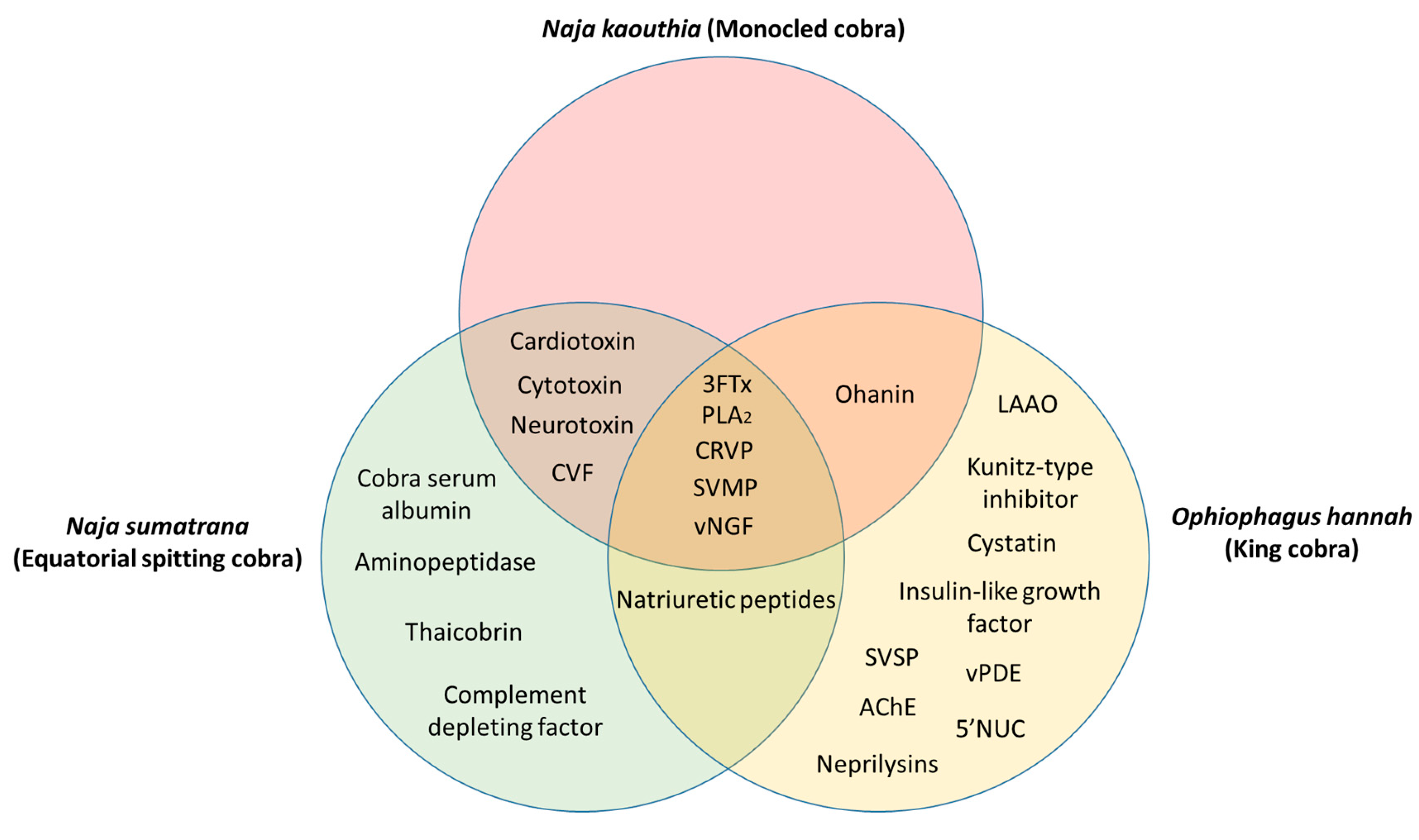
| Proteomic composition of the venom | 3FTx, PLA2, ohanin, CRVP, SVMP, vNGF, cardiotoxin, CVF, cytotoxin, and neurotoxin [36,37] | PLA2, neurotoxins, cardiotoxin, cytotoxin, 3FTX, CVF, SVMP, CRVP, natriuretic peptide, aminopeptidase, thaicobrin, complement-depleting factor, vNGF, and cobra serum albumin [38] | Natriuretic peptides, 3FTx, Kunitz-type inhibitor, PLA2, ohanin, CRVP, cystatin, insulin-like growth factor, SVMP, LAAO, SVSP, vNGF, vPDE, PLB, AChE, 5’NUC, and neprilysins [31,39] |
| Species | Venom/Protein Component | Mechanism | Cancer Cell Type/Tissue | Reference |
|---|---|---|---|---|
| Ophiophagus hannah | Crude venom | Cytotoxic activity on pancreatic cancer cells (EC50; 1.39 ng/mL), reduced migration activity, and induction of apoptosis in PaTu 8988t cells | Patu 8988t | [45] |
| Crude venom | Reduced tumor-cell-induced angiogenesis in vivo | Zebrafish embryos | [45] | |
| L-amino acid oxidase (OH-LAAO) | Antiproliferative activity on murine melanoma, human fibrosarcoma, and murine epithelial cells | B16/F10, HT1080, and Balb/3T3 | [30] | |
| OH-LAAO | Cytotoxic activity on human breast adenocarcinoma cells (EC50 0.05 µg/mL) and apoptosis induction | MCF-7 | [33] | |
| OH-LAAO | Apoptosis induction and inhibition of prostate tumor growth | PC-3 xenograft in nude mice | [32] | |
| OH-LAAO | Induced alteration of gene expression involved in cytotoxicity and apoptotic effects | MCF-7 | [31] | |
| OH-LAAO | Modulation of proteins involved in stress response, ubiquitination, proteolysis, cell proliferation, and apoptosis | MCF-7 | [33] | |
| Naja kaouthia | Crude venom | Cytotoxic activity on pancreatic cancer cells (EC50 1.42 ng/mL) | PaTu 8988t | [45] |
| Crude venom | Venom at a nonlethal dose inhibited tumor-cell proliferation and showed cytotoxic activity and apoptosis induction in human lung cancer cells and leukemic cells | Ehrlich-ascites cells (EAC), U937, K562 | [47] | |
| Cytotoxin CT3 | Histopathological changes in leukemia cells treated with CT3 | A549 and HL60 | [48] | |
| Cardiotoxic–cytotoxic protein | Antiproliferative activity and apoptosis induction in human leukemic cells | U947 and K562 | [49] | |
| kaotree (N. kouthia) and atroporin (Crotalus atrox) combination | Elevated cytotoxic activity in various human cancer cells | HBL-100, BT-20, ZR-75-1, HT-29, and Diji | [50] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zainal Abidin, S.A.; Lee, Y.Q.; Othman, I.; Naidu, R. Malaysian Cobra Venom: A Potential Source of Anti-Cancer Therapeutic Agents. Toxins 2019, 11, 75. https://doi.org/10.3390/toxins11020075
Zainal Abidin SA, Lee YQ, Othman I, Naidu R. Malaysian Cobra Venom: A Potential Source of Anti-Cancer Therapeutic Agents. Toxins. 2019; 11(2):75. https://doi.org/10.3390/toxins11020075
Chicago/Turabian StyleZainal Abidin, Syafiq Asnawi, Yee Qian Lee, Iekhsan Othman, and Rakesh Naidu. 2019. "Malaysian Cobra Venom: A Potential Source of Anti-Cancer Therapeutic Agents" Toxins 11, no. 2: 75. https://doi.org/10.3390/toxins11020075
APA StyleZainal Abidin, S. A., Lee, Y. Q., Othman, I., & Naidu, R. (2019). Malaysian Cobra Venom: A Potential Source of Anti-Cancer Therapeutic Agents. Toxins, 11(2), 75. https://doi.org/10.3390/toxins11020075