Transcriptional Regulation of Aflatoxin Biosynthesis and Conidiation in Aspergillus flavus by Wickerhamomyces anomalus WRL-076 for Reduction of Aflatoxin Contamination


Abstract
:1. Introduction
2. Results and Discussion
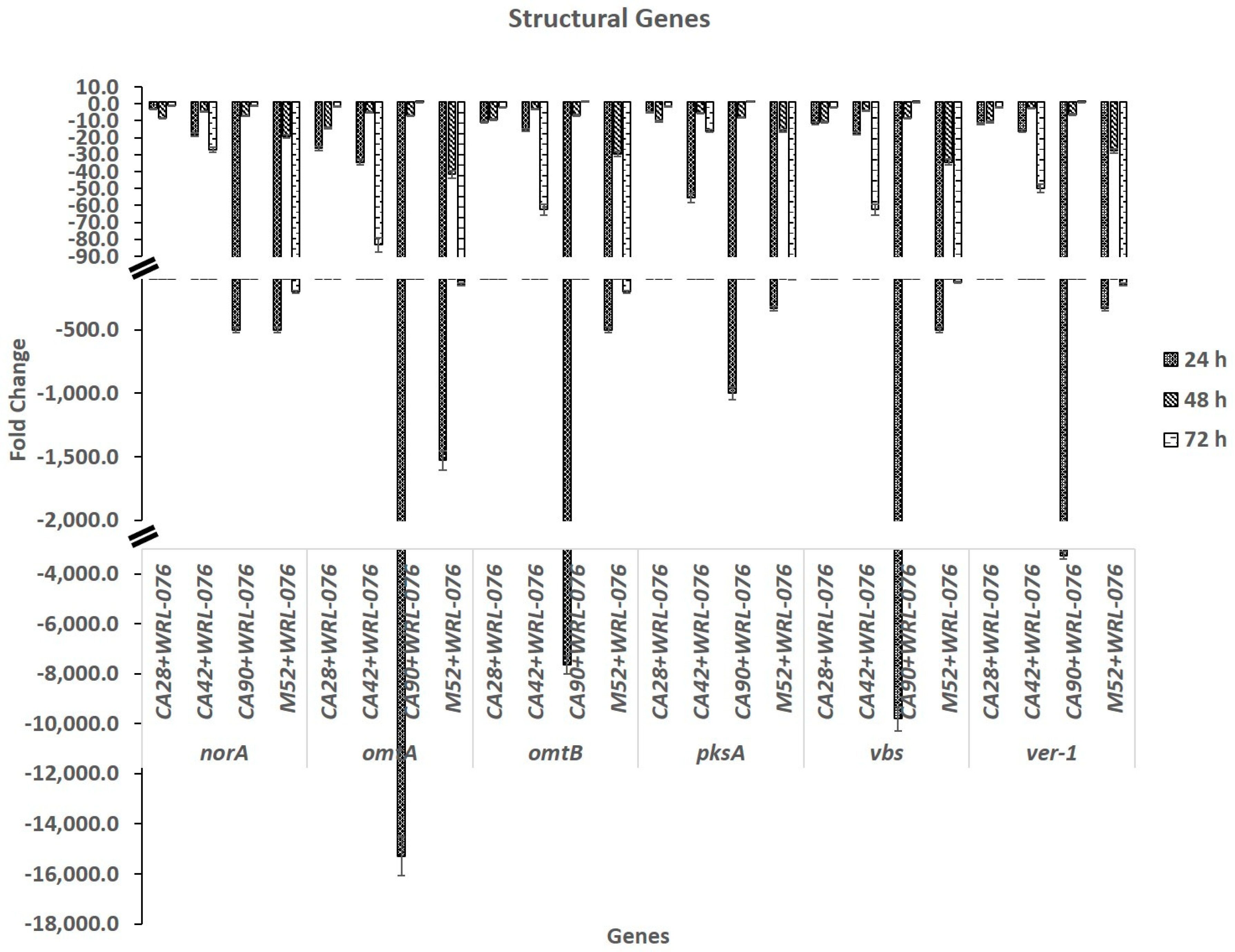
2.1. Down Regulation of Expression of AF Biosynthetic Cluster Genes by W. anomalus
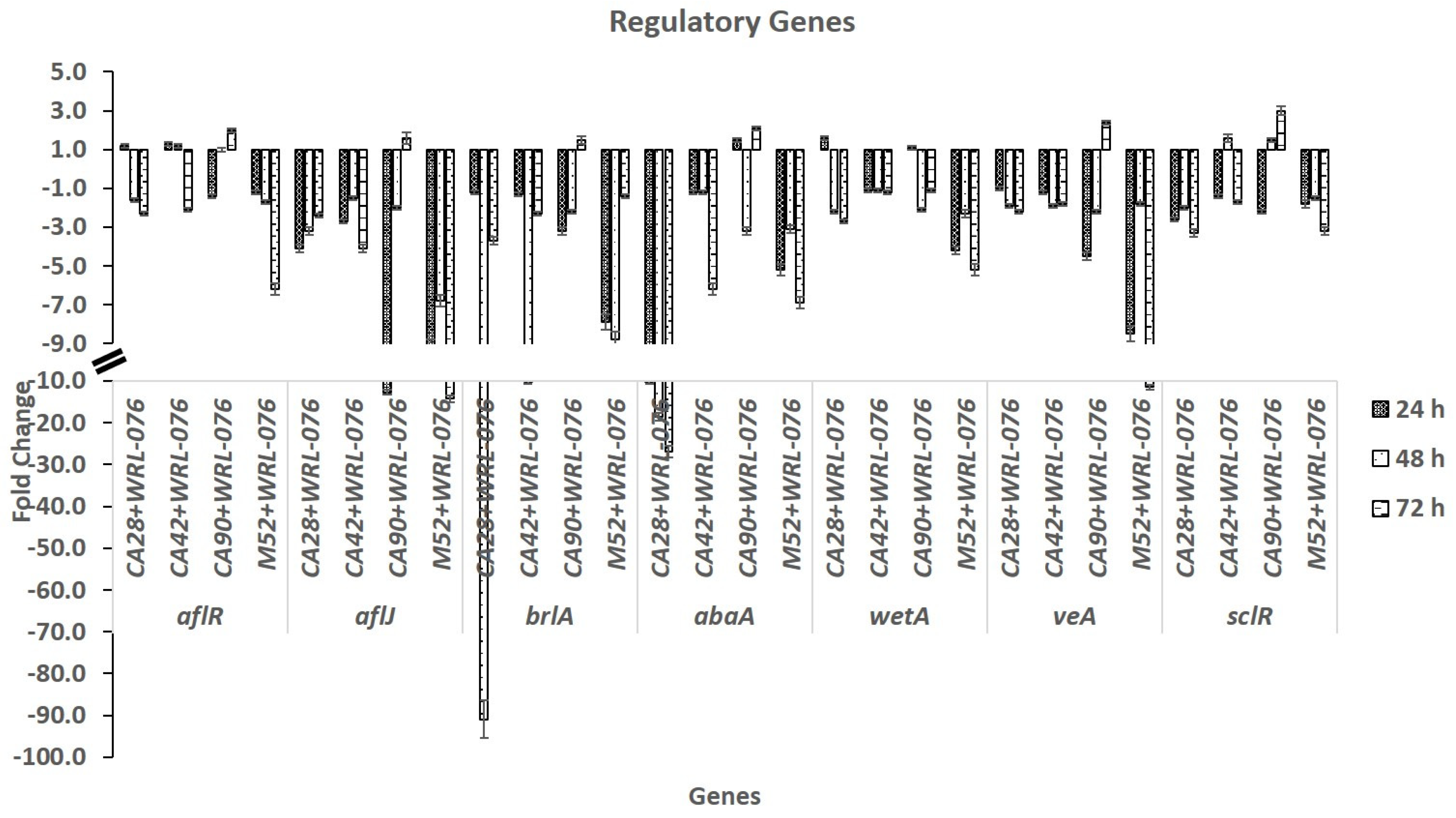
2.2. Effect of W. anomalus on Transcription of Genes of Conidiation and Sclerotial Formation
2.3. Inhibition of Aflatoxin Production in Yeast and A. flavus Dual Cultures
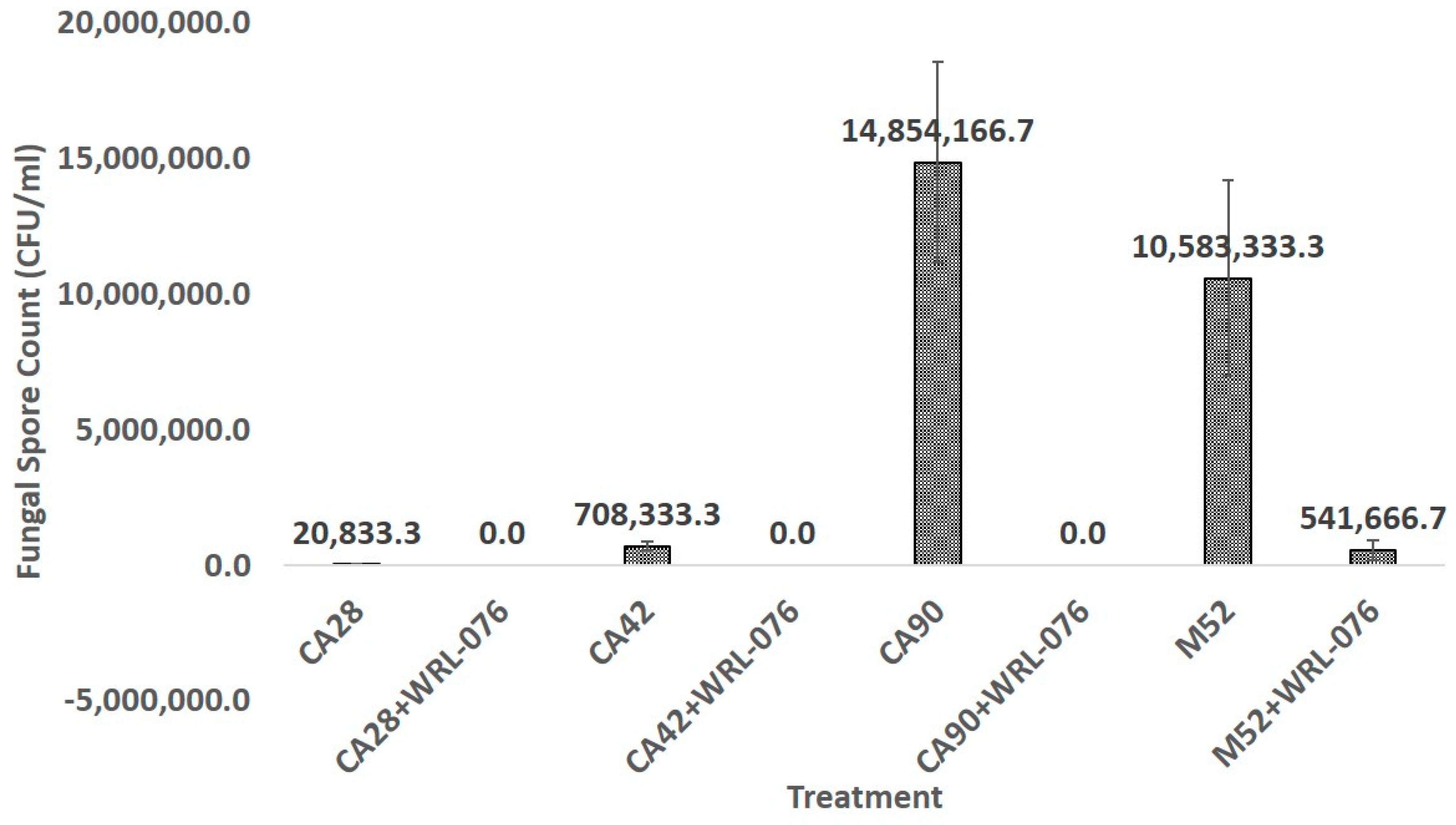
2.4. Reduction of Fungal Biomass and Number of Conidial Formation
3. Conclusions
4. Material and Methods
4.1. Microbial Strains and Media
4.2. Experimental Design
4.3. Real Time RT-PCR Analysis of AF Biosynthetic Genes and Conidia Regulatory Genes
4.4. High Performance Liquid Chromatography (HPLC) Analysis of AFB1
4.5. Determination of Fungal Biomass and Conidia Numbers
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Henry, S.H.; Bosch, F.X.; Bowers, J.C. Aflatoxin, hepatitis and worldwide liver cancer risks. Adv. Exp. Med. Biol. 2002, 504, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Hedayati, M.T.; Pasqualotto, A.C.; Warn, P.A.; Bowyer, P.; Denning, D.W. Aspergillus flavus: Human pathogen, allergen and mycotoxin producer. Microbiology 2007, 153, 1677–1692. [Google Scholar] [CrossRef] [PubMed]
- Molyneux, R.J.; Mahoney, N.; Kim, J.H.; Campbell, B.C. Mycotoxins in edible tree nuts. Int. J. Food Microbiol. 2007, 119, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Trucksess, M.W.; Scott, P.M. Mycotoxins in botanicals and dried fruits: A review. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2008, 25, 181–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Roze, L.V.; Hong, S.-Y.; Linz, J.E. Aflatoxin Biosynthesis: Current Frontiers. Annu. Rev. Food Sci. Technol. 2013, 4, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Hua, S.S.T.; Chang, P.K.; Palumbo, J.D. Mycotoxins. In Toxins and Other Harmful Compounds in Foods; Witczak, A., Sikorski, Z.E., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2017; pp. 153–168. [Google Scholar]
- Van Egmond, H.P.; Schothorst, R.C.; Jonker, M.A. Regulations relating to mycotoxins in food: Perspectives in a global and European context. Anal. Bioanal. Chem. 2007, 389, 147–157. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). Declaration of the World Summit on Food Security; WFSFS 2009/2; FAO: Rome, Italy, November 2009; pp. 16–18. [Google Scholar]
- Commission of the European Community. Commission Directive 98/53/EC of July 1998 laying down the sampling methods and the methods of analysis for the official control of the levels of certain contaminants in food stuffs. Off. Eur. Commun. Legis. 1998, L201, 93–101. [Google Scholar]
- Kurtzman, C.P. Recognition of Yeast Species from Gene Sequence Comparisons. Open Appl. Inform. J. 2011, 5, 20–29. [Google Scholar] [CrossRef]
- Petersson, S.; Schnurer, J. Biocontrol of mold growth in high-moisture wheat stored under airtight conditions by Pichia anomala, Pichia guilliermondii, and Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1995, 61, 1027–1032. [Google Scholar]
- Petersson, S.; Schnurer, J. Pichia anomala as a biocontrol agent of Penicillium roqueforti in high-moisture wheat, rye, barley, and oats stored under airtight conditions. Can. J. Microbiol. 1998, 44, 471–476. [Google Scholar] [CrossRef]
- Schnürer, J.; Jonsson, A. Pichia anomala J121: A 30-year overnight near success biopreservation story. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2011, 99, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Jijakli, M.H.; Lepoivre, P. Characterization of an Exo-beta-1,3-Glucanase Produced by Pichia anomala Strain K, Antagonist of Botrytis cinerea on Apples. Phytopathology 1998, 88, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Haïssam, J.M. Pichia anomala in biocontrol for apples: 20 years of fundamental research and practical applications. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2011, 99, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Petersson, S.; Hansen, M.W.; Axberg, K.; Hult, K.; Schnürer, J. Ochratoxin a accumulation in cultures of Penicillium verrucosum with the antagonistic yeast Pichia anomala and Saccharomyces cerevisiae. Mycol. Res. 1998, 102, 1003–1008. [Google Scholar] [CrossRef]
- Hua, S.S.T.; Baker, J.L.; Flores-Espiritu, M. Interactions of saprophytic yeasts with a nor mutant of Aspergillus flavus. Appl. Environ. Microbiol. 1999, 65, 2738–2740. [Google Scholar] [PubMed]
- Hua, S.S.T. Potential use of saprophytic yeast to reduce populations of Aspergillus flavus in almond and pistachio orchards. In Proceedings of the Third International Symposium of Pistachio and Almond, Zaragoza, Spain, 20–24 May 2001; Battle, I., Hormaza, I., Espiau, M.T., Eds.; 2002; Volume 591, pp. 527–530. [Google Scholar]
- Hua, S.S.T. Application of a yeast, Pichia anomala strain WRL-076 to control Aspergillus flavus for reducing aflatoxin in pistachio and almond. IOBC Bulletin 2004, 27, 291–294. [Google Scholar]
- Hua, S.S.T.; Parfitt, D.E.; Holtz, B.A. Evaluation of a biopesticide, Pichia anomala WRL-076 to control Aspergillus flavus in a commercial orchard. In Proceedings of the California Conference of Biological Control V, Riverside, CA, USA, 25–27 July 2006; pp. 152–155. [Google Scholar]
- Hua, S.S.T. Biocontrol of Aspergillus flavus by Pichia anomala. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Mendez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2013; pp. 1067–1074. [Google Scholar]
- Isakeit, T.; Bétran, F.J.; Odvody, G.; Hua, S.S.T. Efficacy of Pichia anomala WLR-076 to control aflatoxin on corn in Texas. 2005; Plant Dis Rep 1. 2007, FC021. Available online: https://www.ars.usda.gov/research/publications/publication/?seqNo115=223081 (accessed on 28 January 2019).
- Hua, S.S.T.; Hernlem, B.J.; Yokoyama, W.; Sarreal, S.B.L. Intracellular trehalose and sorbitol synergistically promoting cell viability of a biocontrol yeast, Pichia anomala, for aflatoxin reduction. World J. Microbiol. Biotechnol. 2015, 31, 729–734. [Google Scholar] [CrossRef]
- Hua, S.S.T.; Brandl, M.; Hernlem, B.; Eng, J.G.; Sarreal, S.B.L. Fluorescent viability stains to probe the metabolic status of aflatoxigenic fungus in dual culture of Aspergillus flavus and Pichia anomala. Mycopathologia 2011, 171, 133–138. [Google Scholar] [CrossRef]
- Hua, S.S.T.; Beck, J.J.; Sarreal, S.B.L.; Gee, W. The major volatile compound 2-phenylethanol from the biocontrol yeast, Pichia anomala, inhibits growth and expression of aflatoxin biosynthetic genes of Aspergillus flavus. Mycotoxin Res. 2014, 30. [Google Scholar] [CrossRef]
- Chang, P.K.; Hua, S.S.T.; Sarreal, S.B.L.; Li, R.W. Suppression of aflatoxin biosynthesis in Aspergillus flavus by 2-phenylethanol is associated with stimulated growth and decreased degradation of branched-chain amino acids. Toxins 2015, 7, 3887–3902. [Google Scholar] [CrossRef] [PubMed]
- Bayman, P.; Cotty, P.J. Genetic diversity in Aspergillus flavus: Association with aflatoxin production and morphology. Can. J. Bot. 1993, 71, 23–31. [Google Scholar] [CrossRef]
- Ehrlich, K.C.; Montalbano, B.G.; Cotty, P.J. Divergent regulation of aflatoxin production at acidic pH by two Aspergillus strains. Mycopathologia 2005, 159, 579–581. [Google Scholar] [CrossRef] [PubMed]
- Ohkura, M.; Cotty, P.J.; Orbach, M.J. Comparative genomics of Aspergillus flavus S and L morphotypes yield Insights into niche adaptation. G3 (Bethesda) 2018, 8, 3915–3930. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chang, P.K.; Ehrlich, K.C.; Cary, J.W.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E.; Woloshuk, C.P.; Bennett, J.W. Clustered Pathway Genes in Aflatoxin Biosynthesis. Appl. Environ. Microbiol. 2004, 70, 1253–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlich, K.C.; Yu, J.; Cotty, P.J. Aflatoxin biosynthesis gene clusters and flanking regions. J. Appl. Microbiol. 2005, 99, 518–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins (Basel) 2012, 41, 24. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, K.C.; Montalbano, B.G.; Cotty, P.J. Sequence comparison of aflR from different Aspergillus species provides evidence for variability in regulation of aflatoxin production. Fungal Genet. Biol. 2003, 38, 63–74. [Google Scholar] [CrossRef]
- Calvo, A.M.; Bok, J.; Brooks, W.; Keller, N.P. veA is required for toxin and sclerotial production in Aspergillus parasiticus. Appl. Environ. Microbiol. 2004, 70, 4733–4739. [Google Scholar] [CrossRef]
- Calvo, A.M. The VeA regulatory system and its role in morphological and chemical development in fungi. Fungal Genet. Biol. 2008, 45, 1053–1061. [Google Scholar] [CrossRef]
- Georgianna, D.R.; Payne, G.A. Genetic regulation of aflatoxin biosynthesis: From gene to genome. Fungal Genet. Biol. 2009, 46, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Amare, M.G.; Keller, N.P. Molecular mechanisms of Aspergillus flavus secondary metabolism and development. Fungal Genet. Biol. 2014, 66, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Adams, T.H.; Boylan, M.T.; Timberlake, W.E. brlA is necessary and sufficient to direct conidiophore development in Aspergillus nidulans. Cell 1988, 54, 353–362. [Google Scholar] [CrossRef]
- Adams, T.H.; Wieser, J.K.; Yu, J.H. Asexual sporulation in Aspergillus nidulans. Microbiol. Mol. Biol. Rev. 1998, 62, 35–54. [Google Scholar] [PubMed]
- Andrianopoulos, A.; Timberlake, W.E. The Aspergillus nidulans abaA gene encodes a transcriptional activator that acts as a genetic switch to control development. Mol. Cell. Biol. 1994, 14, 2503–2515. [Google Scholar] [CrossRef] [PubMed]
- Marshall, M.A.; Timberlake, W.E. Aspergillus nidulans wetA activates spore-specific gene expression. Mol. Cell. Biol. 1991, 11, 55–62. [Google Scholar] [CrossRef]
- Etxebeste, O.; Garzia, A.; Espeso, E.A.; Ugalde, U. Aspergillus nidulans asexual development: Making the most of cellular modules. Trends Microbiol. 2010, 18, 569–576. [Google Scholar] [CrossRef]
- Wu, M.Y.; Mead, M.E.; Kim, S.C.; Rokas, A.; Yu, J.H. WetA bridges cellular and chemical development in Aspergillus flavus. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Jin, F.J.; Takahashi, T.; Matsushima, K.I.; Hara, S.; Shinohara, Y.; Maruyama, J.I.; Kitamoto, K.; Koyama, Y. SclR, a basic helix-loop-helix transcription factor, regulates hyphal morphology and promotes sclerotial formation in Aspergillus oryzae. Eukaryot. Cell 2011, 10, 945–955. [Google Scholar] [CrossRef]
- Wicklow, D.T. Survival of Aspergillus flavus sclerotia in soil. Trans. Br. Mycol. Soc. 1987, 89, 131–134. [Google Scholar] [CrossRef]
- Wicklow, D.T.; Wilson, D.M.; Nelsen, T.C. Survival of Aspergillus-Flavus Sclerotia and Conidia Buried in Soil in Illinois or Georgia. Phytopathology 1993, 83, 1141–1147. [Google Scholar] [CrossRef]
- Cary, J.W.; OBrian, G.R.; Nielsen, D.M.; Nierman, W.; Harris-Coward, P.; Yu, J.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Calvo, A.M. Elucidation of veA-dependent genes associated with aflatoxin and sclerotial production in Aspergillus flavus by functional genomics. Appl. Microbiol. Biotechnol. 2007, 76, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Duran, R.M.; Cary, J.W.; Calvo, A.M. Production of cyclopiazonic acid, aflatrem, and aflatoxin by Aspergillus flavus is regulated by veA, a gene necessary for sclerotial formation. Appl. Microbiol. Biotechnol. 2007, 73, 1158–1168. [Google Scholar] [CrossRef] [PubMed]
- Cotty, P.J. Aflatoxin-producing potential of communities of Aspergillus section Flavi from cotton producing areas in the United States. Mycol. Res. 1997, 101, 698–704. [Google Scholar] [CrossRef]
- Chang, P.K.; Scharfenstein, L.L.; Mack, B.; Ehrlich, K.C. Deletion of the Aspergillus flavus orthologue of A. Nidulans fluG reduces conidiation and promotes production of sclerotia but does not abolish aflatoxin biosynthesis. Appl. Environ. Microbiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Pfaffl, M. Quantification strategies in real-time PCR. In A–Z of Quantitative PCR; International University Line: San Diego, CA, USA, 2004; pp. 87–112. ISBN 0963681788. [Google Scholar]
- Hua, S.S.T.; Palumbo, J.D.; Parfitt, D.; Sarreal, S.L.; O’Keeffe, T. Development of a droplet digital PCR assay for population analysis of aflatoxigenic and atoxigenic Aspergillus flavus mixtures in soil. Mycotoxin Res. 2018, 34, 187–194. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Enzyme/Product | Functions |
|---|---|---|
| aflR | transcription activator AflR | pathway-specific regulator |
| aflJ | transcription enhancer | pathway co-regulator |
| norA | norsolorinic acid reductase | NOR → AVN |
| omtA | O-methyltransferase A | ST → OMST |
| omtB | O-methyltransferase B | DHDMST → DHST |
| pksA | polyketide synthase (PKS) | acetate → polyketide |
| vbs | versicolorin B synthase | VAL → VERB |
| ver-1 | dehydrogenase/ketoreductase | VERA → DMST |
| brlA | C2H2 zinc finger transcription factor | control of conidiophore development |
| abaA | conidial formation (abacus) | conidiation regulator activated by BrlA |
| wetA | conidial formation (wet-white conidia) | conidiation regulator activated by AbaA |
| veA | velvet nuclear complex component | global regulator |
| sclR | sclerotial formation | sclerotium regulator |
| Gene | CA28+WRL-076 | CA42+WRL-076 | CA90+WRL-076 | M52+WRL-076 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | |
| aflR | 1.2 ± 0.1 | −1.6 ± 0.1 | −2.3 ± 0.1 | 1.3 ± 0.1 | 1.2 ± 0.1 | −2.1 ± 0.1 | −1.4 ± 0.1 | 1.0 ± 0.1 | 2.0 ± 0.1 | −1.2 ± 0.1 | −1.7 ± 0.1 | −6.2 ± 0.3 |
| aflJ | −4.1 ± 0.2 | −3.2 ± 0.2 | −2.4 ± 0.1 | −2.7 ± 0.1 | −1.5 ± 0.1 | −4.1 ± 0.2 | −12.7 ± 0.6 | −2.0 ± 0.1 | 1.6 ± 0.3 | −9.3 ± 0.5 | −6.8 ± 0.3 | −14.3 ± 0.7 |
| norA | −2.8 ± 0.1 | −8.5 ± 0.4 | −1.1 ± 0.1 | −18.2 ± 0.9 | −4.5 ± 0.2 | −27.0 ± 1.4 | −500.0 ± 25.0 | −7.1 ± 0.4 | −1.0 ± 0.1 | −500.0 ± 25.0 | −19.2 ± 1.0 | −200.0 ± 10.0 |
| omtA | −26.3 ± 1.3 | −14.3 ± 0.7 | −1.7 ± 0.1 | −34.5 ± 1.7 | −5.2 ± 0.3 | −83.3 ± 4.2 | −15,290.5 ± 764.5 | −6.9 ± 0.3 | 1.2 ± 0.1 | −1,526.7 ± 76.3 | −41.7 ± 2.1 | −142.9 ± 7.1 |
| omtB | −10.8 ± 0.5 | −9.3 ± 0.5 | −1.8 ± 0.1 | −15.4 ± 0.8 | −3.1 ± 0.2 | −62.5 ± 3.1 | −7,633.6 ± 381.7 | −6.8 ± 0.3 | 1.3 ± 0.1 | −500.0 ± 24.9 | −29.4 ± 1.5 | −200.0 ± 9.8 |
| pksA | −5.2 ± 0.3 | −10.4 ± 0.5 | −1.6 ± 0.1 | −55.6 ± 2.8 | −5.5 ± 0.3 | −16.1 ± 0.8 | −1,000.0 ± 50.0 | −8.1 ± 0.4 | 1.4 ± 0.1 | −333.3 ± 16.7 | −15.9 ± 0.8 | −100.0 ± 5.0 |
| vbs | −11.5 ± 0.6 | −11.0 ± 0.5 | −1.8 ± 0.1 | −17.5 ± 0.9 | −3.9 ± 0.2 | −62.5 ± 3.1 | −9,803.9 ± 490.2 | −8.5 ± 0.4 | 1.2 ± 0.1 | −500.0 ± 24.8 | −34.5 ± 1.7 | −125.0 ± 6.3 |
| ver−1 | −11.9 ± 0.6 | −10.9 ± 0.5 | −2.1 ± 0.1 | −16.1 ± 0.8 | −2.5 ± 0.1 | −50.0 ± 2.5 | −3,257.3 ± 162.9 | −6.3 ± 0.3 | 1.2 ± 0.1 | −333.3 ± 16.4 | −27.8 ± 1.4 | −142.9 ± 7.3 |
| brlA | −1.2 ± 0.1 | −90.9 ± 4.5 | −3.7 ± 0.2 | −1.3 ± 0.1 | −10.3 ± 0.5 | −2.3 ± 0.1 | −3.2 ± 0.2 | −2.2 ± 0.1 | 1.5 ± 0.2 | −7.9 ± 0.4 | −8.8 ± 0.4 | −1.4 ± 0.1 |
| abaA | −10.2 ± 0.5 | −18.5 ± 0.9 | −27.0 ± 1.4 | −1.2 ± 0.1 | −1.2 ± 0.1 | −6.2 ± 0.3 | 1.5 ± 0.1 | −3.2 ± 0.2 | 2.1 ± 0.1 | −5.2 ± 0.3 | −3.1 ± 0.2 | −6.9 ± 0.3 |
| wetA | 1.6 ± 0.1 | −2.2 ± 0.1 | −2.7 ± 0.1 | −1.1 ± 0.1 | −1.1 ± 0.1 | −1.2 ± 0.1 | 1.1 ± 0.1 | −2.1 ± 0.1 | −1.1 ± 0.1 | −4.2 ± 0.2 | −2.3 ± 0.2 | −5.2 ± 0.3 |
| veA | −1.0 ± 0.1 | −1.9 ± 0.1 | −2.2 ± 0.1 | −1.2 ± 0.1 | −1.9 ± 0.1 | −1.8 ± 0.1 | −4.5 ± 0.2 | −2.2 ± 0.1 | 2.4 ± 0.1 | −8.5 ± 0.4 | −1.8 ± 0.1 | −11.5 ± 0.6 |
| sclR | −2.6 ± 0.1 | −2.0 ± 0.1 | −3.3 ± 0.2 | −1.4 ± 0.1 | 1.6 ± 0.2 | −1.7 ± 0.1 | −2.2 ± 0.1 | 1.5 ± 0.1 | 3.0 ± 0.2 | −1.8 ± 0.2 | −1.5 ± 0.1 | −3.2 ± 0.2 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| 18S | TTCCTAGCGAGCCCAACCT | GCCCGCCGAAGCAACT |
| aflR | GCCGCGCCCGAAA | GCACTTTTGAGCTGGCACAA |
| aflJ | CCGAAGATTCCGCTTGGA | TGAAGACATGCAGCAAAAGGA |
| norA | TCTAGCGCCGGTGTTCGT | CATTGCCGAAGCTCATCGTT |
| omtA | CGGGTTTCGCAAAAGCAT | GCAGGCAGGTCCTGTACGA |
| omtB | TGCTGTGGCATCCATTCAAA | GGACTGCGTCTTCCAAAAGG |
| pksA | TCACAAGCGATGCACAGTTG | AACTGACGAATGTGGGTCTTGTACT |
| vbs | GAGTCTACCGCCGCCGATA | GAAAAGGTCGGCCAGTCATC |
| ver-1 | GGTCCCCAAGCATGCTGTA | GCAGCGAACAAAGGTGTCAAT |
| brlA | TCAAGACGCACAGCCCTACA | GACGCGGTGCCGATAGAG |
| abaA | GAGTGGCAGACCGAATGTATGTTG | TAGTGGTAGGCATTGGGTGAGTTG |
| wetA | CCACAGCAGCCGATCCA | CCCCTTGCAGGATGTCATG |
| veA | TGGACCGCCCATCTCAAG | ATGCCGCACGGAAAGATC |
| sclR | TGCCGCACACAACATCATT | TTCTCCAAGGCCACGAACTT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hua, S.S.T.; Sarreal, S.B.L.; Chang, P.-K.; Yu, J. Transcriptional Regulation of Aflatoxin Biosynthesis and Conidiation in Aspergillus flavus by Wickerhamomyces anomalus WRL-076 for Reduction of Aflatoxin Contamination. Toxins 2019, 11, 81. https://doi.org/10.3390/toxins11020081
Hua SST, Sarreal SBL, Chang P-K, Yu J. Transcriptional Regulation of Aflatoxin Biosynthesis and Conidiation in Aspergillus flavus by Wickerhamomyces anomalus WRL-076 for Reduction of Aflatoxin Contamination. Toxins. 2019; 11(2):81. https://doi.org/10.3390/toxins11020081
Chicago/Turabian StyleHua, Sui Sheng T., Siov Bouy L. Sarreal, Perng-Kuang Chang, and Jiujiang Yu. 2019. "Transcriptional Regulation of Aflatoxin Biosynthesis and Conidiation in Aspergillus flavus by Wickerhamomyces anomalus WRL-076 for Reduction of Aflatoxin Contamination" Toxins 11, no. 2: 81. https://doi.org/10.3390/toxins11020081
APA StyleHua, S. S. T., Sarreal, S. B. L., Chang, P. -K., & Yu, J. (2019). Transcriptional Regulation of Aflatoxin Biosynthesis and Conidiation in Aspergillus flavus by Wickerhamomyces anomalus WRL-076 for Reduction of Aflatoxin Contamination. Toxins, 11(2), 81. https://doi.org/10.3390/toxins11020081