Treatment and Prevention of Recurrent Clostridium difficile Infection with Functionalized Bovine Antibody-Enriched Whey in a Hamster Primary Infection Model

 , ,
, ,
Abstract
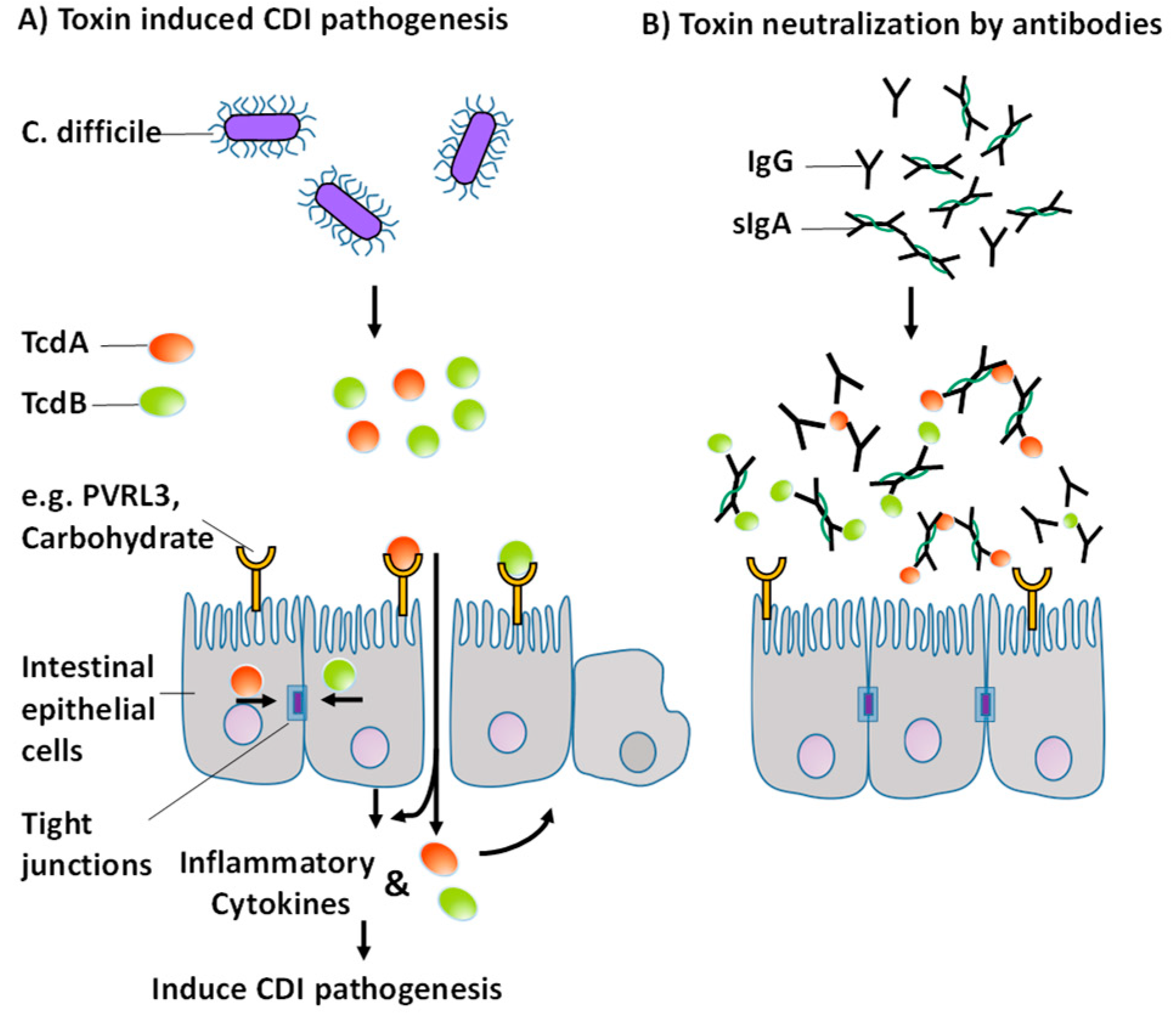
:1. Introduction
2. Results
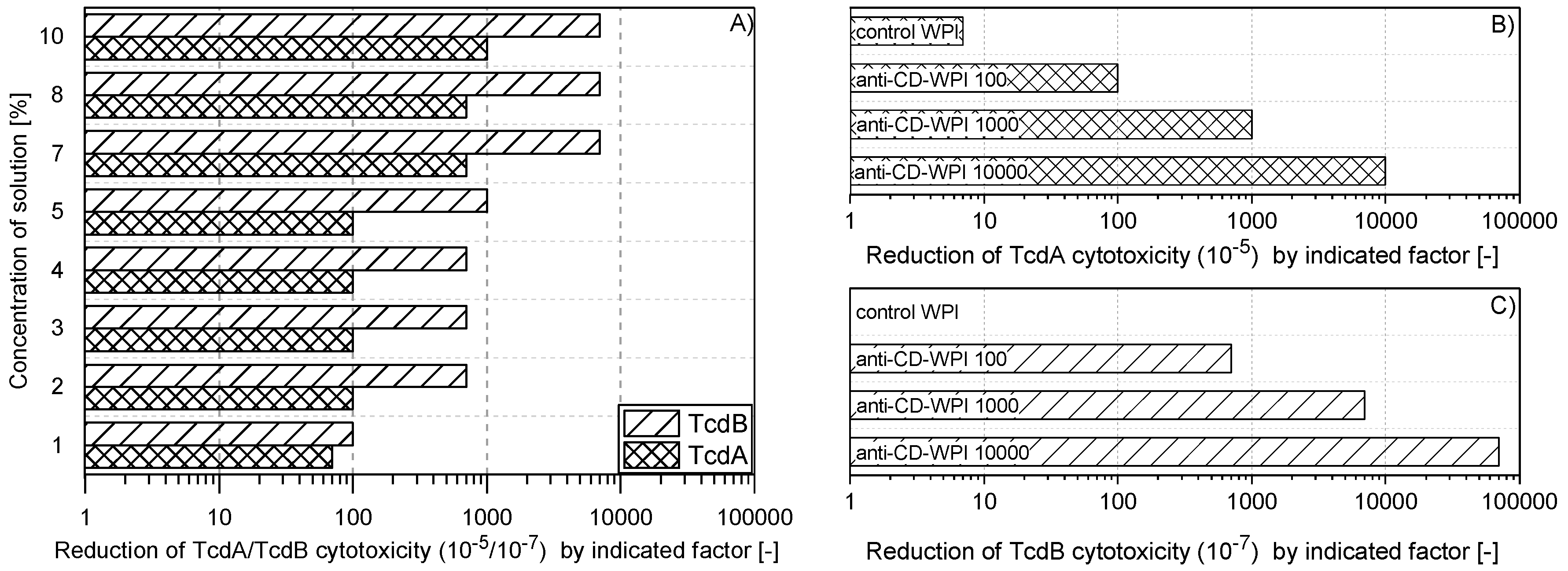
2.1. Reduction of TcdA and TcdB Cytotoxicity of WPI Solutions
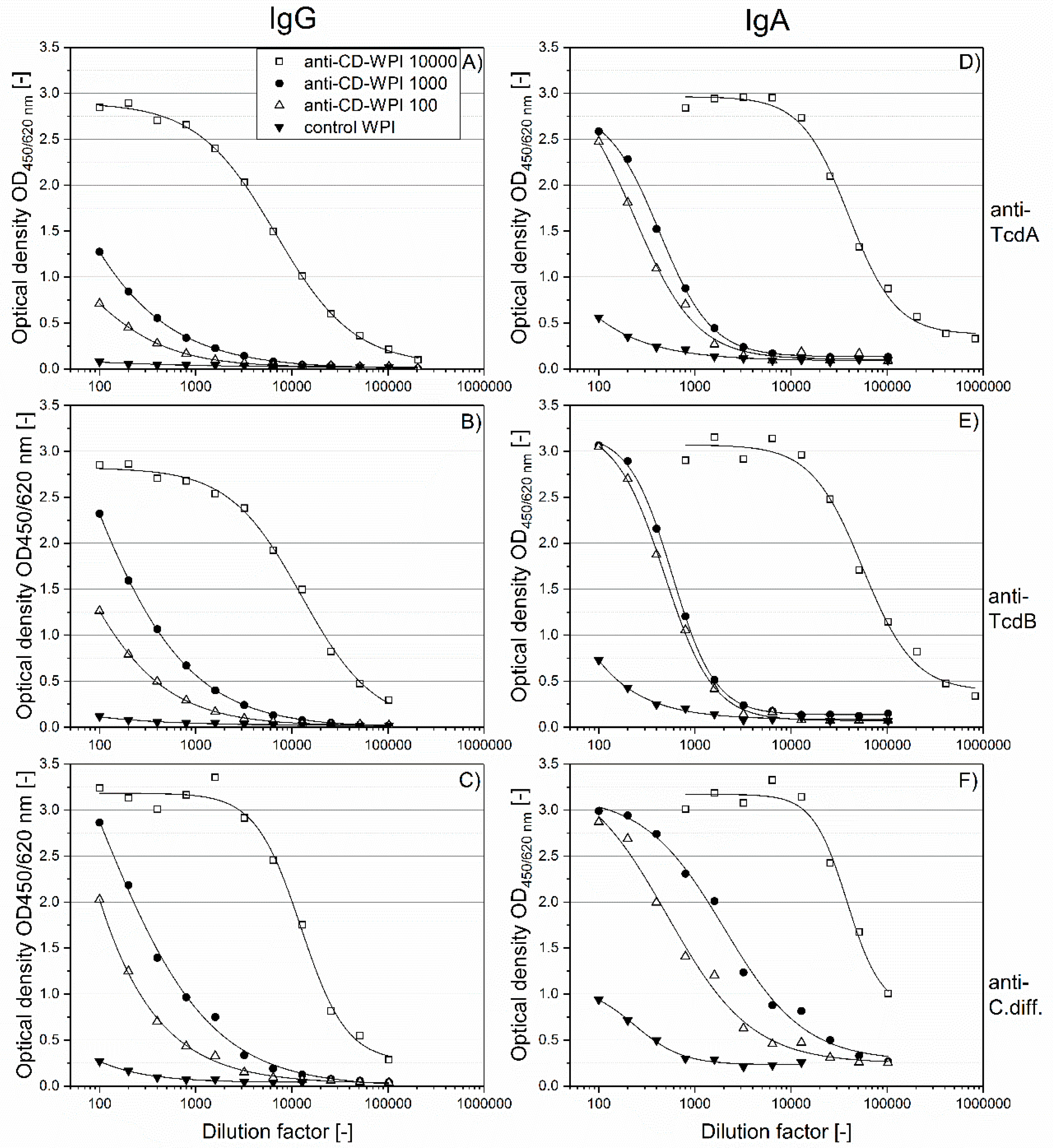
2.2. Specific Immunoglobulin Titers
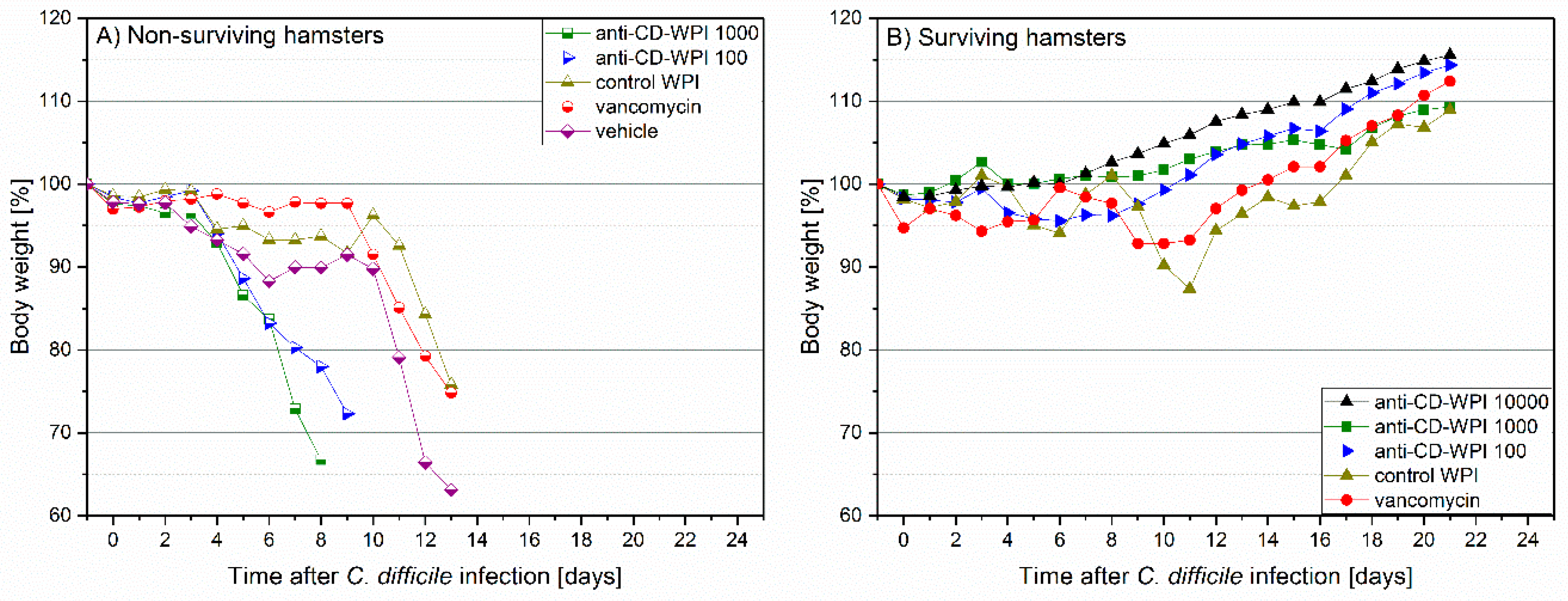
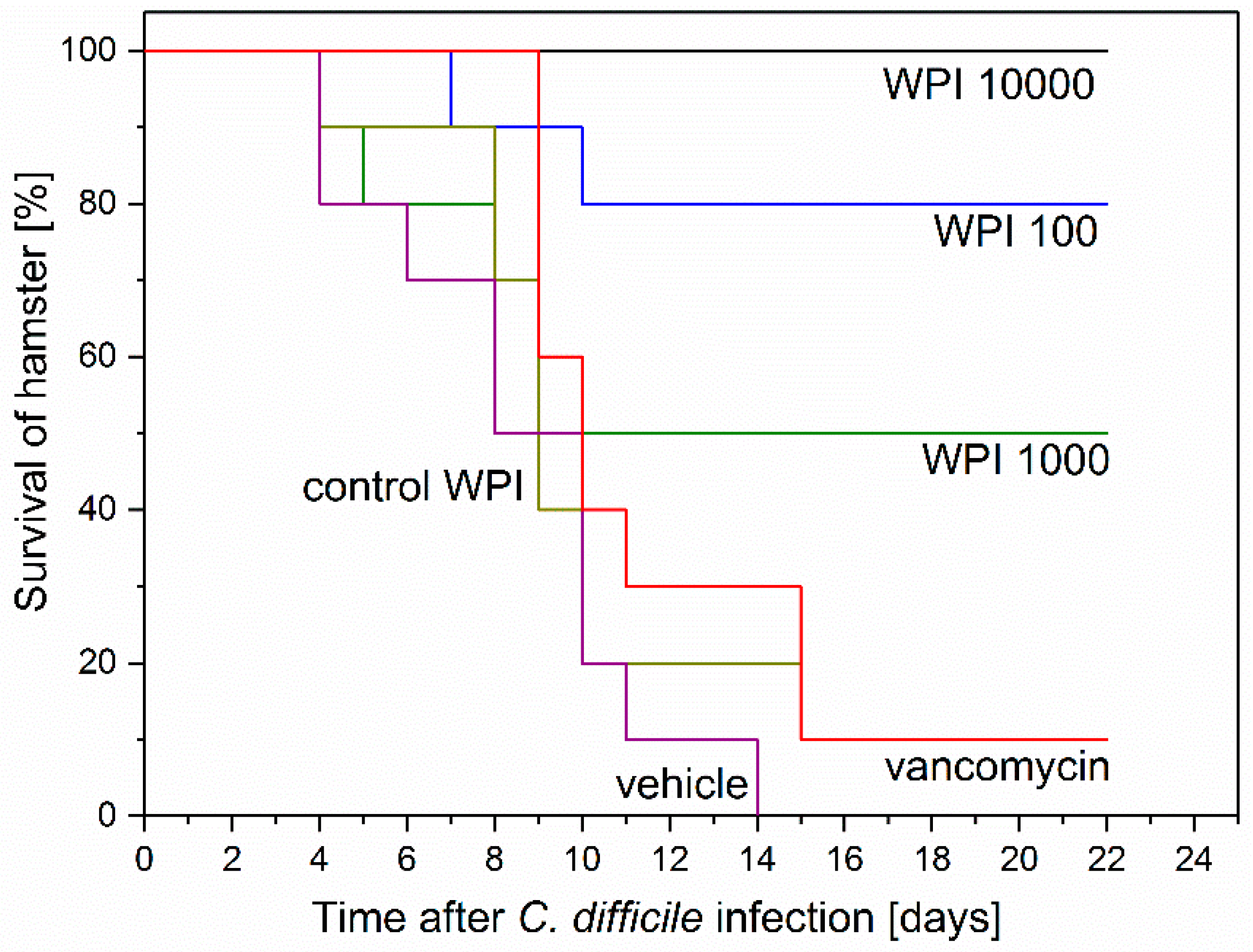
2.3. Prevention of Acute and Recurrent CDI with Anti-CD-WPI
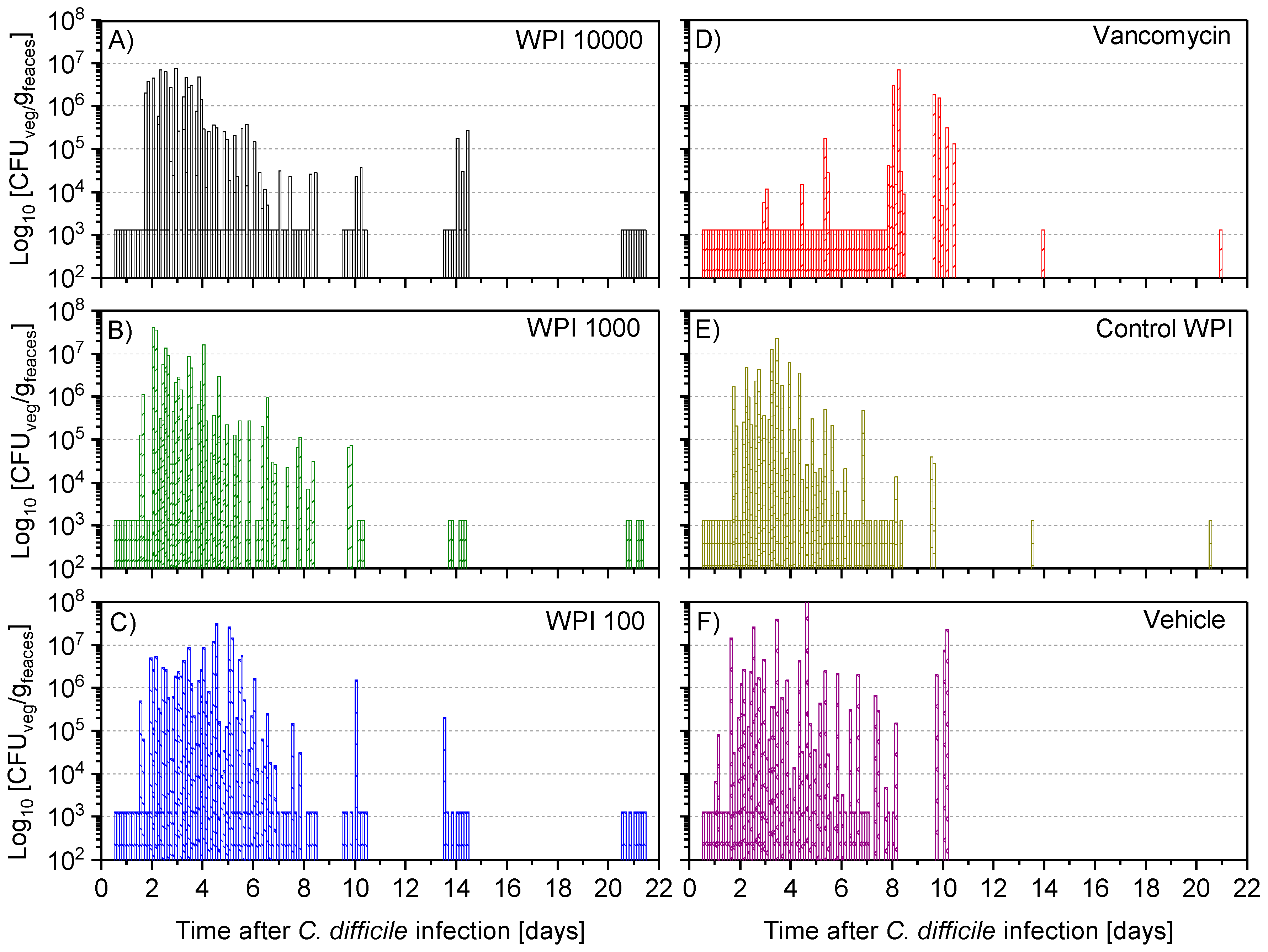
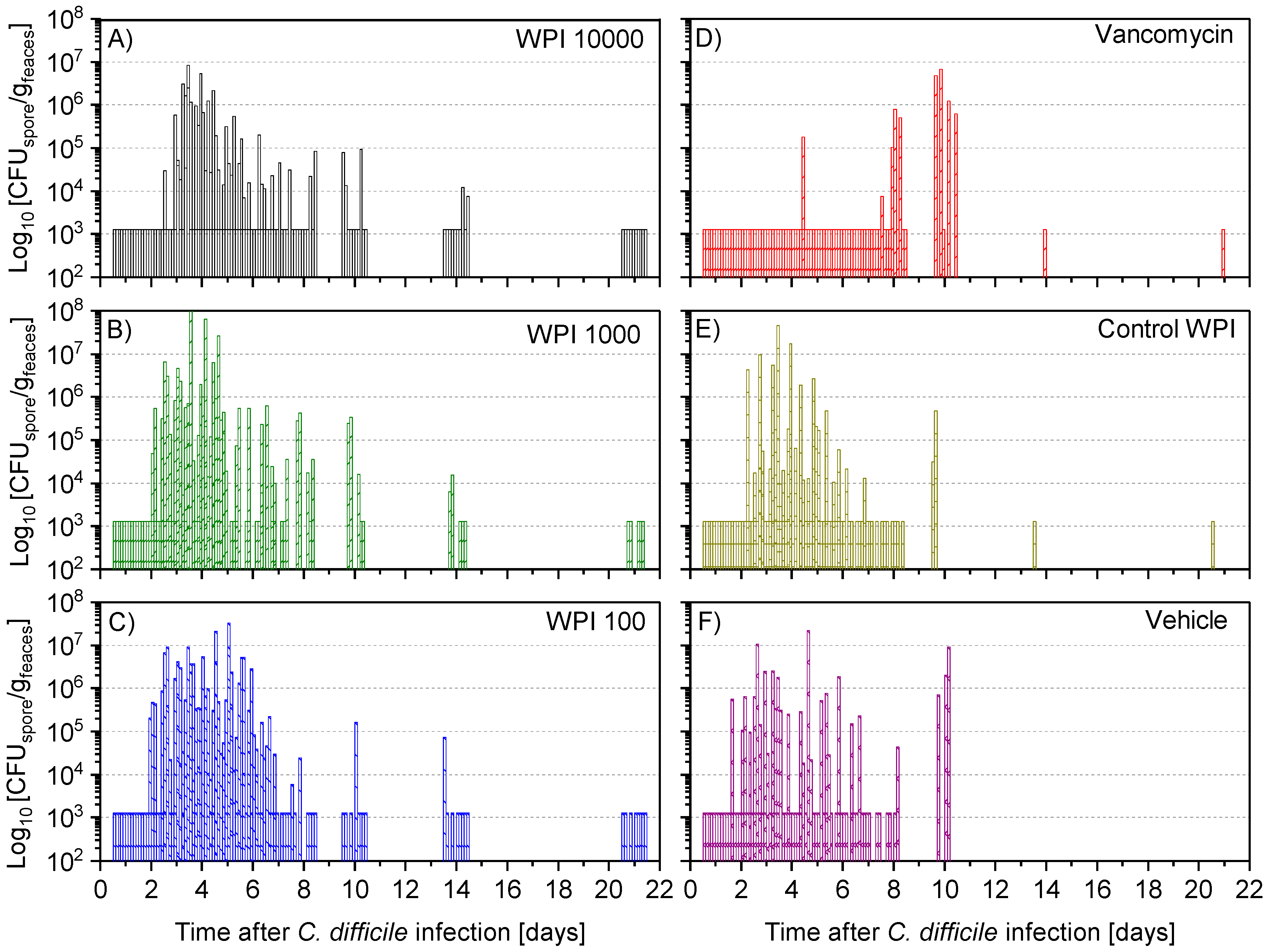
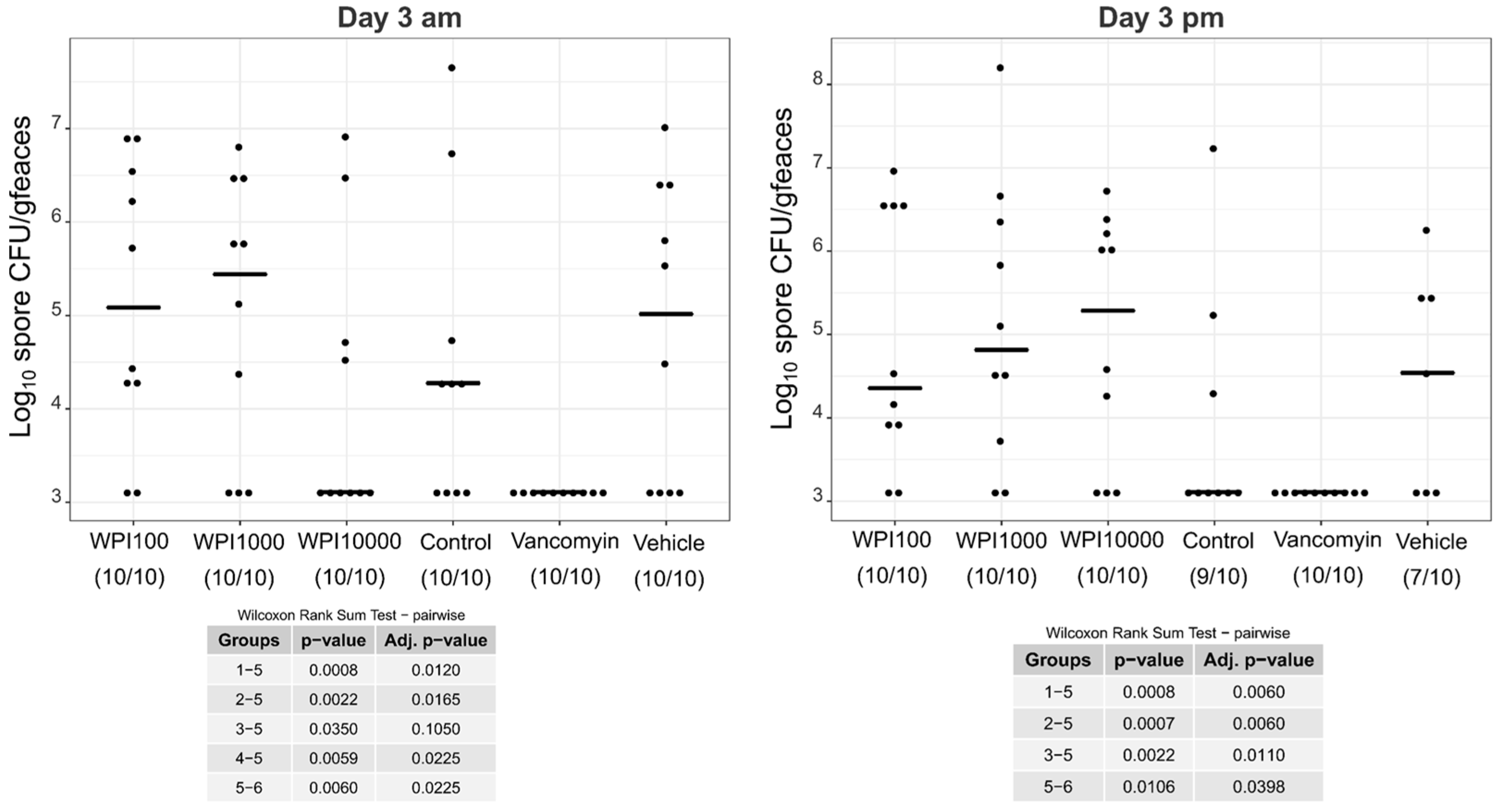
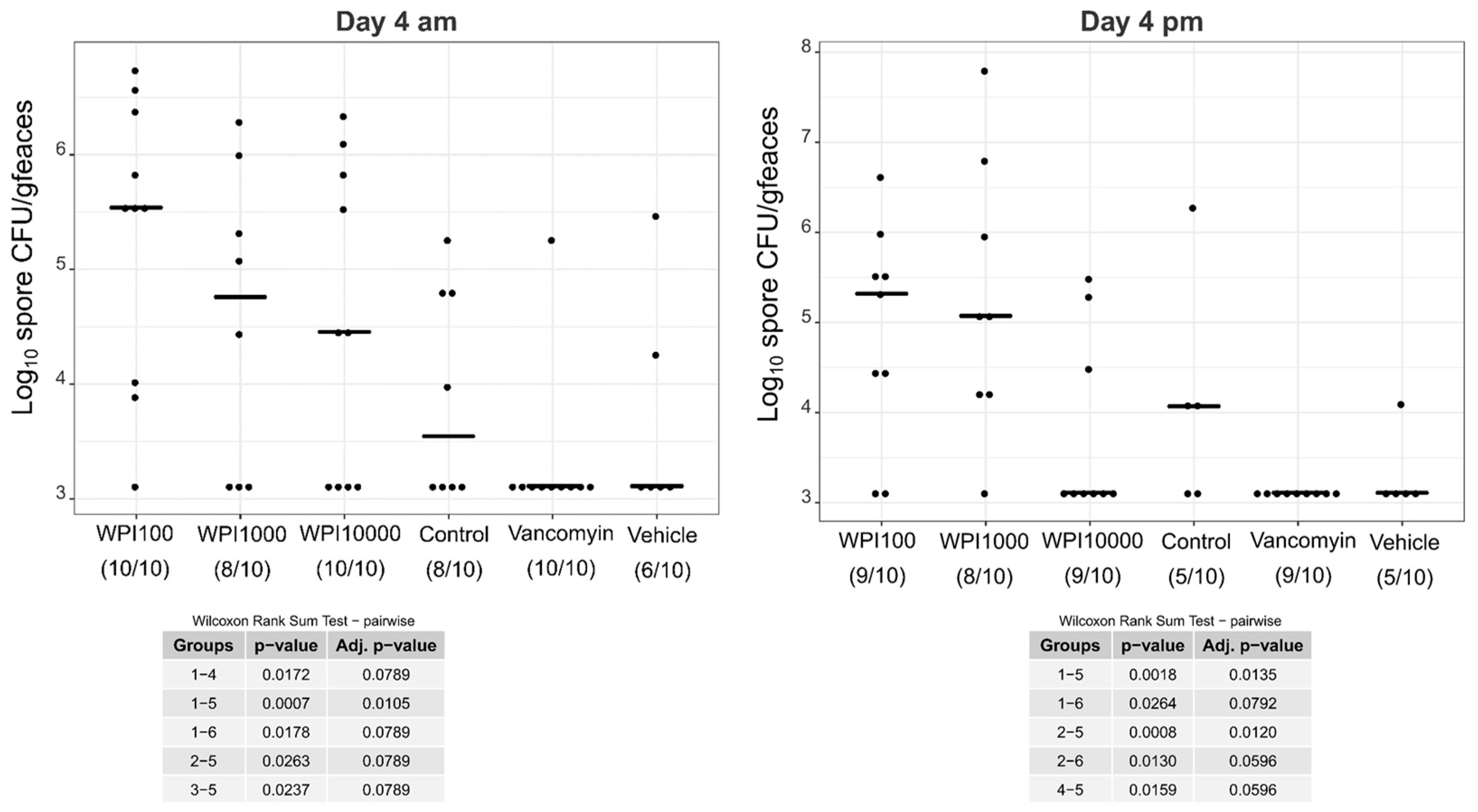
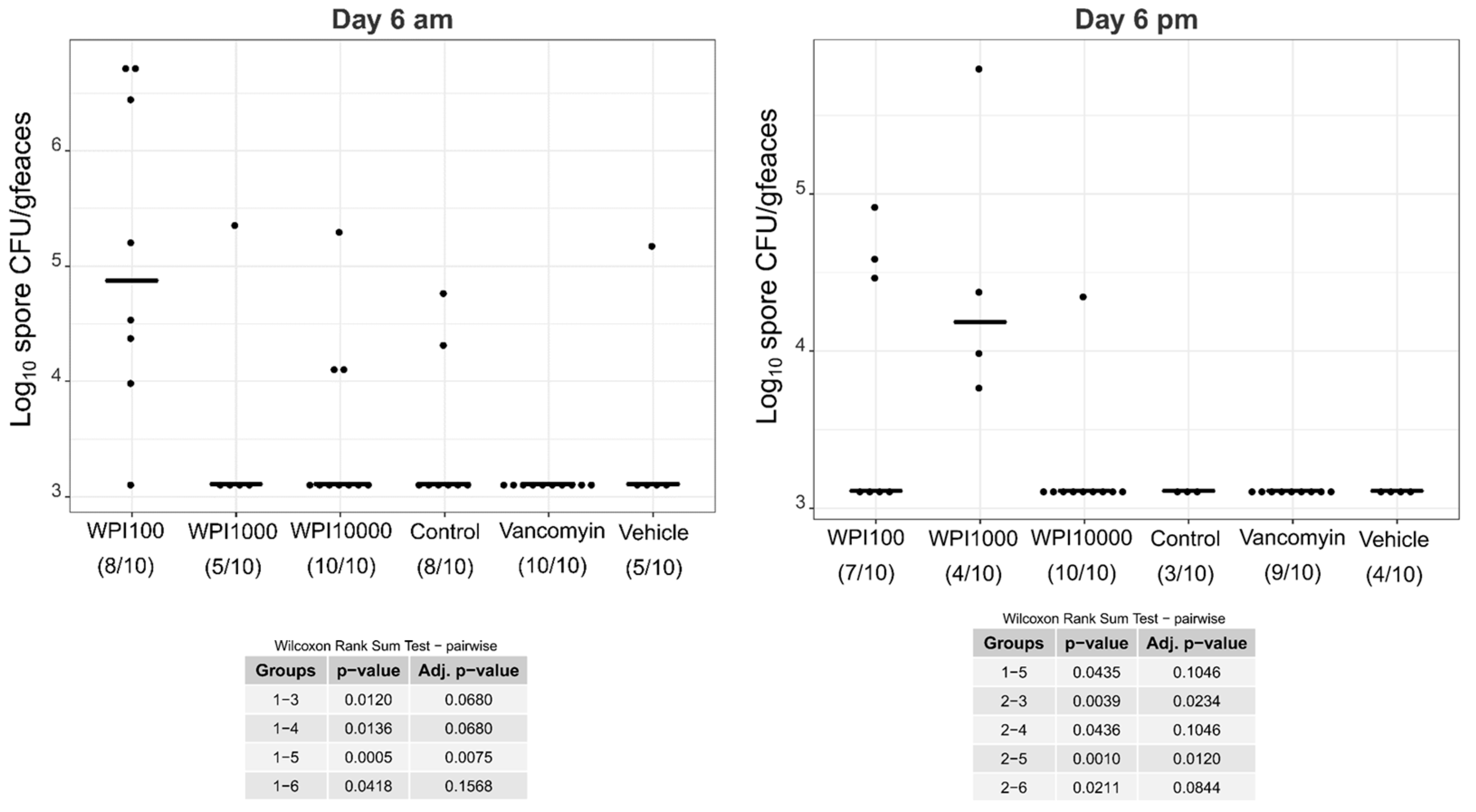
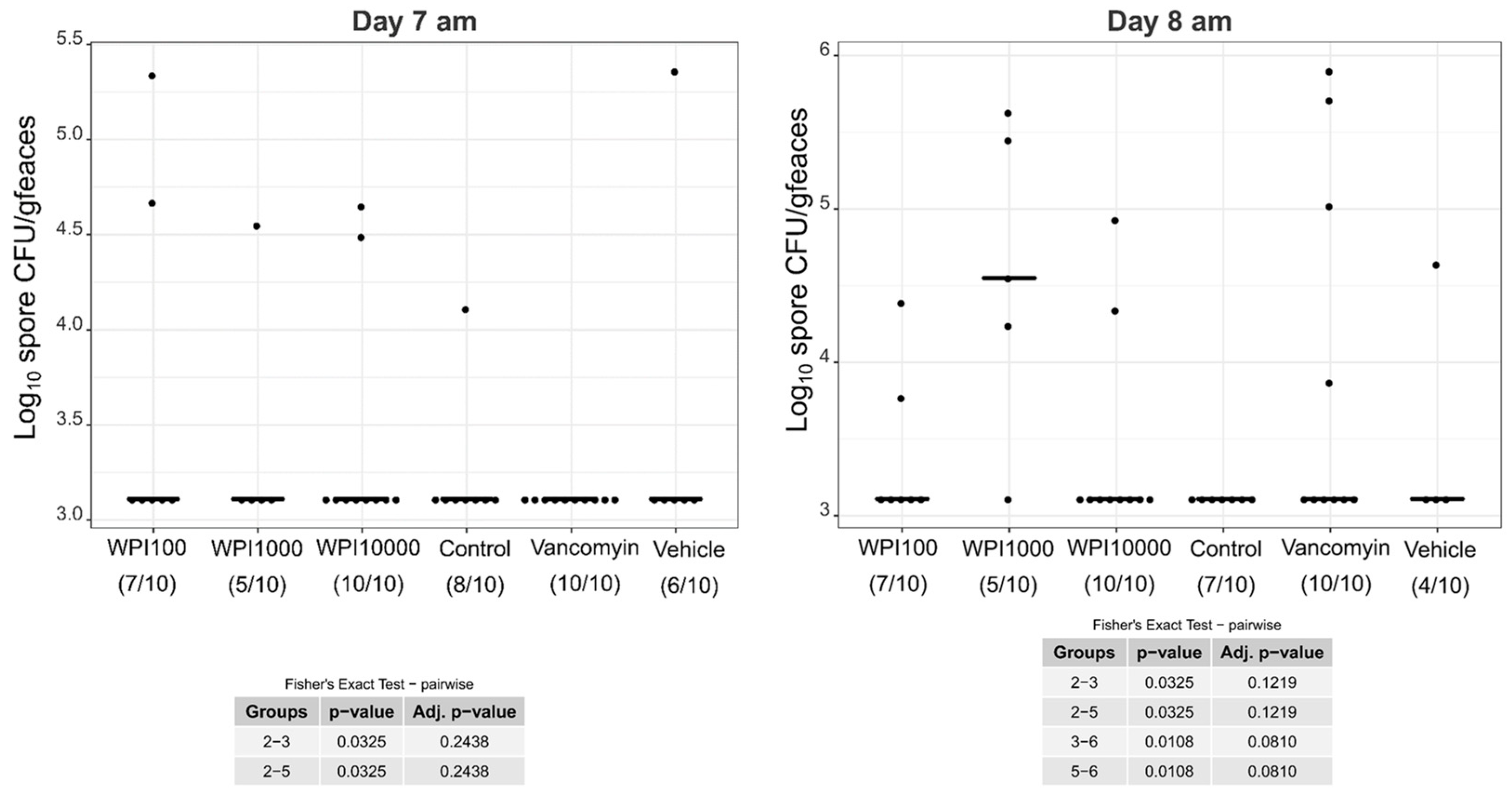
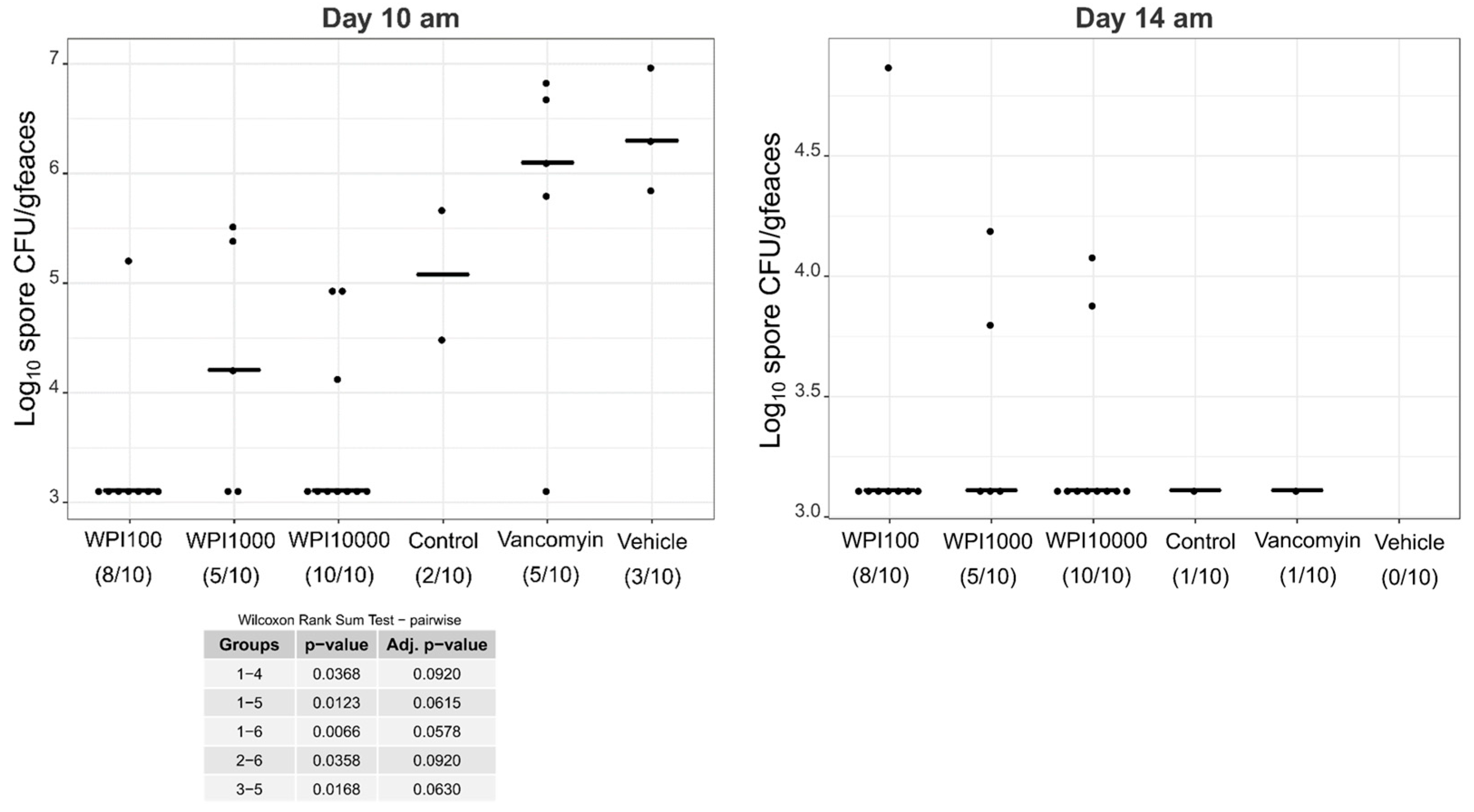
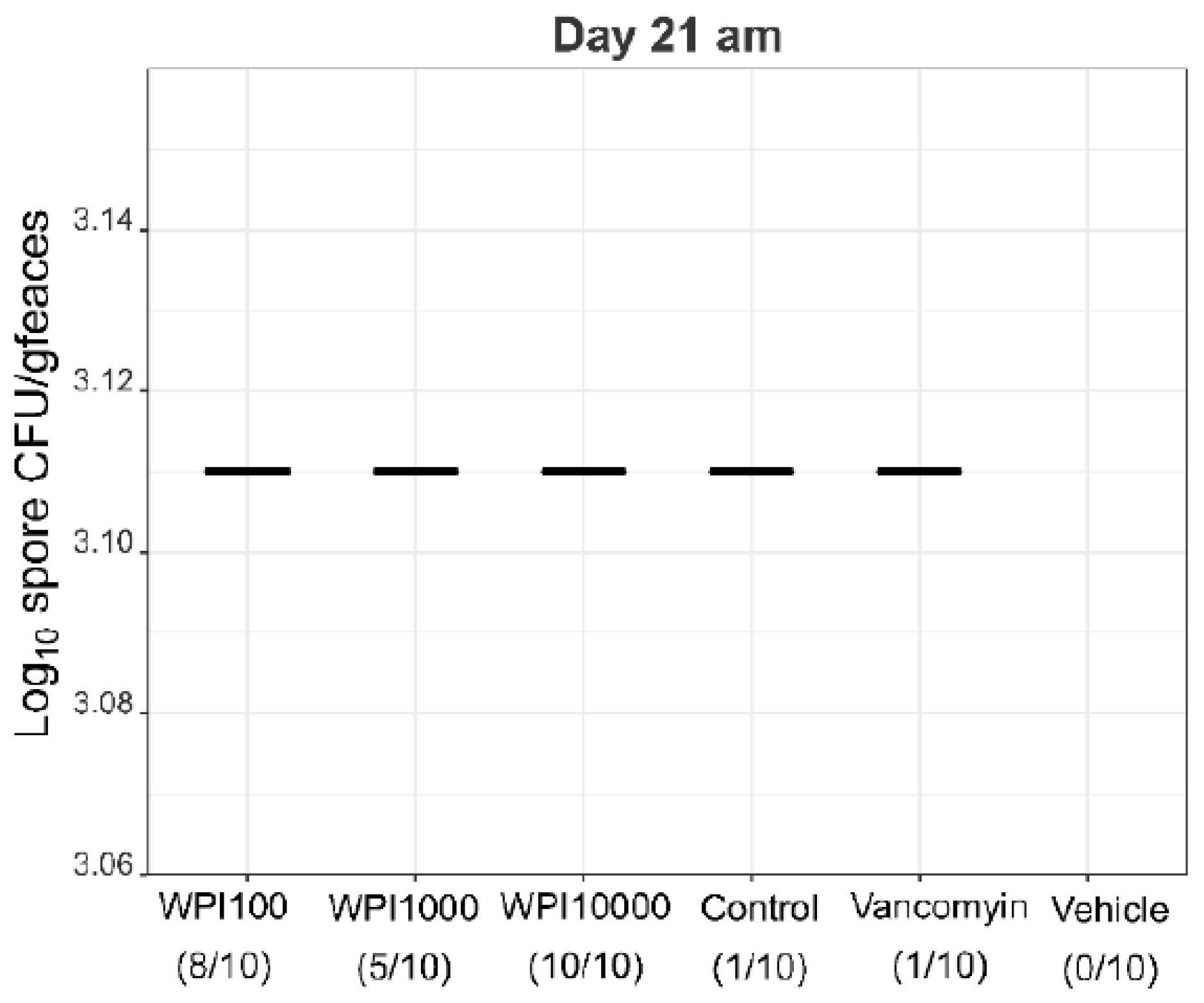
2.4. Fecal Vegetative and Spore Colony-Forming Units
3. Discussion
4. Materials and Methods
4.1. Preparation of Whey Protein Isolates
4.2. Whey Protein Characterization
4.3. Determination of IgG and IgA against TcdA, TcdB and C. difficile 630
4.4. Inhibition of Cytotoxicity of TcdA and TcdB
4.5. Hamster Treatment
4.6. Fecal Total CFU Detection and Spore Detection
4.7. Data Regression and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A







References
- Lessa, F.C.; Mu, Y.; Bamberg, W.M.; Beldavs, Z.G.; Dumyati, G.K.; Dunn, J.R.; Farley, M.M.; Holzbauer, S.M.; Meek, J.I.; Phipps, E.C.; et al. Burden of Clostridium difficile infection in the United States. N. Engl. J. Med. 2015, 372, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.J.; Curns, A.T.; McDonald, L.C.; Parashar, U.D.; Lopman, B.A. The roles of Clostridium difficile and norovirus among gastroenteritis-associated deaths in the United States, 1999–2007. Clin. Infect. Dis. 2012, 55, 216–223. [Google Scholar] [CrossRef] [PubMed]
- McDonald, L.C.; Gerding, D.N.; Johnson, S.; Bakken, J.S.; Carroll, K.C.; Coffin, S.E.; Dubberke, E.R.; Garey, K.W.; Gould, C.V.; Kelly, C.; et al. Clinical Practice Guidelines for Clostridium difficile Infection in Adults and Children: 2017 Update by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA). Clin. Infect. Dis. 2018, 66, e1–e48. [Google Scholar] [CrossRef] [PubMed]
- Abt, M.C.; McKenney, P.T.; Pamer, E.G. Clostridium difficile colitis: Pathogenesis and host defence. Nat. Rev. Microbiol. 2016, 14, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Seekatz, A.M.; Young, V.B. Clostridium difficile and the microbiota. J. Clin. Invest. 2014, 124, 4182–4189. [Google Scholar] [CrossRef] [PubMed]
- Kuehne, S.A.; Collery, M.M.; Kelly, M.L.; Cartman, S.T.; Cockayne, A.; Minton, N.P. Importance of toxin A, toxin B, and CDT in virulence of an epidemic Clostridium difficile strain. J. Infect. Dis. 2014, 209, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Poutanen, S.M.; Simor, A.E. Clostridium difficile-associated diarrhea in adults. CMAJ 2004, 171, 51–58. [Google Scholar] [CrossRef]
- Reineke, J.; Tenzer, S.; Rupnik, M.; Koschinski, A.; Hasselmayer, O.; Schrattenholz, A.; Schild, H.; von Eichel-Streiber, C. Autocatalytic cleavage of Clostridium difficile toxin B. Nature 2007, 446, 415–419. [Google Scholar] [CrossRef]
- McFarland, L.V. Alternative treatments for Clostridium difficile disease: What really works? J. Med. Microbiol. 2005, 54, 101–111. [Google Scholar] [CrossRef]
- Sartelli, M.; Malangoni, M.A.; Abu-Zidan, F.M.; Griffiths, E.A.; Di Bella, S.; McFarland, L.V.; Eltringham, I.; Shelat, V.G.; Velmahos, G.C.; Kelly, C.P.; et al. WSES guidelines for management of Clostridium difficile infection in surgical patients. World J. Emerg. Surg. 2015, 10, 38. [Google Scholar] [CrossRef]
- Young, K.W.H.; Munro, I.C.; Taylor, S.L.; Veldkamp, P.; van Dissel, J.T. The safety of whey protein concentrate derived from the milk of cows immunized against Clostridium difficile. Regul. Toxicol. Pharmacol. 2007, 47, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, H.; Marnila, P.; Gill, H. Bovine milk antibodies for health. BJN 2000, 84. [Google Scholar] [CrossRef]
- Steele, J.; Sponseller, J.; Schmidt, D.; Cohen, O.; Tzipori, S. Hyperimmune bovine colostrum for treatment of GI infections: A review and update on Clostridium difficile. Hum. Vaccin. Immunother. 2013, 9, 1565–1568. [Google Scholar] [CrossRef] [PubMed]
- Freedman, D.J.; Tacket, C.O.; Delehanty, A.; Maneval, D.R.; Nataro, J.; Crabb, J.H. Milk immunoglobulin with specific activity against purified colonization factor antigens can protect against oral challenge with enterotoxigenic Escherichia coli. J. Infect. Dis. 1998, 177, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Otto, W.; Najnigier, B.; Stelmasiak, T.; Robins-Browne, R.M. Randomized control trials using a tablet formulation of hyperimmune bovine colostrum to prevent diarrhea caused by enterotoxigenic Escherichia coli in volunteers. Scand. J. Gastroenterol. 2011, 46, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.C.; Wong, V.; Muller, B.; Rawlin, G.; Brown, L.E. Prevention and treatment of influenza with hyperimmune bovine colostrum antibody. PloS ONE 2010, 5, e13622. [Google Scholar] [CrossRef] [PubMed]
- Davidson, G.P.; Daniels, E.; Nunan, H.; Moore, A.G.; Whyte, P.B.D.; Franklin, K.; Mccloud, P.I.; Moore, D.J. Passive immunisation of children with bovine colostrum containing antibodies to human rotavirus. Lancet 1989, 334, 709–712. [Google Scholar] [CrossRef]
- Lyerly, D.M.; Bostwick, E.; Binion, S.; Wilkins, T.D. Passive immunization of hamsters against disease caused by Clostridium difficile by use of bovine immunoglobulin G concentrate. Infect. Immun. 1991, 56. [Google Scholar]
- van Dissel, J.T.; de Groot, N.; Hensgens, C.M.; Numan, S.; Kuijper, E.J.; Veldkamp, P.; van’t Wout, J. Bovine antibody-enriched whey to aid in the prevention of a relapse of Clostridium difficile-associated diarrhoea: Preclinical and preliminary clinical data. J. M. Microbiol. 2005, 54, 197–205. [Google Scholar] [CrossRef]
- Mattila, E.; Anttila, V.-J.; Broas, M.; Marttila, H.; Poukka, P.; Kuusisto, K.; Pusa, L.; Sammalkorpi, K.; Dabek, J.; Koivurova, O.-P.; et al. A randomized, double-blind study comparing Clostridium difficile immune whey and metronidazole for recurrent Clostridium difficile-associated diarrhoea: Efficacy and safety data of a prematurely interrupted trial. Scand. J. Infect. Dis. 2008, 40, 702–708. [Google Scholar] [CrossRef]
- Sponseller, J.K.; Steele, J.A.; Schmidt, D.J.; Kim, H.B.; Beamer, G.; Sun, X.; Tzipori, S. Hyperimmune bovine colostrum as a novel therapy to combat Clostridium difficile infection. J. Infect. Dis. 2015, 211, 1334–1341. [Google Scholar] [CrossRef] [PubMed]
- Steele, J.; Feng, H.; Parry, N.; Tzipori, S. Piglet models of acute or chronic Clostridium difficile illness. J. Infect. Dis. 2010, 201, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Hutton, M.L.; Cunningham, B.A.; Mackin, K.E.; Lyon, S.A.; James, M.L.; Rood, J.I.; Lyras, D. Bovine antibodies targeting primary and recurrent Clostridium difficile disease are a potent antibiotic alternative. Sci. Rep. 2017, 7, 3665. [Google Scholar] [CrossRef] [PubMed]
- Marnila, P.; Korhonen, H. Milk | Colostrum. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 591–597. [Google Scholar]
- Crottet, P.; Corthésy, B. Secretory component delays the conversion of secretory IgA into antigen-binding competent F(ab′)2: A possible implication for mucosal defense. J. Immunol. 1998, 161, 5445–5453. [Google Scholar] [PubMed]
- Woof, J.M.; Kerr, M.A. The function of immunoglobulin A in immunity. J. Pathol. 2006, 208, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.P.; Chetham, S.; Keates, S.; Bostwick, E.F.; Roush, A.M.; Castagliuolo, I.; LaMont, J.T.; Pothoulakis, C. Survival of anti-Clostridium difficile bovine immunoglobulin concentrate in the human gastrointestinal tract. Antimicrob. Agents Chemother. 1997, 41, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Best, E.L.; Freeman, J.; Wilcox, M.H. Models for the study of Clostridium difficile infection. Gut Microbes 2012, 3, 145–167. [Google Scholar] [CrossRef]
- Dionne, L.-L.; Raymond, F.; Corbeil, J.; Longtin, J.; Gervais, P.; Longtin, Y. Correlation between Clostridium difficile bacterial load, commercial real-time PCR cycle thresholds, and results of diagnostic tests based on enzyme immunoassay and cell culture cytotoxicity assay. J. Clin. Microbiol. 2013, 51, 3624–3630. [Google Scholar] [CrossRef]
- Engevik, M.A.; Yacyshyn, M.B.; Engevik, K.A.; Wang, J.; Darien, B.; Hassett, D.J.; Yacyshyn, B.R.; Worrell, R.T. Human Clostridium difficile infection: Altered mucus production and composition. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G510–G524. [Google Scholar] [CrossRef]
- Sattar, A.; Thommes, P.; Payne, L.; Warn, P.; Vickers, R.J. SMT19969 for Clostridium difficile infection (CDI): In vivo efficacy compared with fidaxomicin and vancomycin in the hamster model of CDI. J. Antimicrob. Chemother. 2015, 70, 1757–1762. [Google Scholar] [CrossRef]
- Warn, P.; Thommes, P.; Sattar, A.; Corbett, D.; Flattery, A.; Zhang, Z.; Black, T.; Hernandez, L.D.; Therien, A.G. Disease Progression and Resolution in Rodent Models of Clostridium difficile Infection and Impact of Antitoxin Antibodies and Vancomycin. Antimicrob. Agents Chemother. 2016, 60, 6471–6482. [Google Scholar] [CrossRef]
- Smits, W.K.; Lyras, D.; Lacy, D.B.; Wilcox, M.H.; Kuijper, E.J. Clostridium difficile infection. Nat. Rev. Dis. Primers 2016, 2, 16020. [Google Scholar] [CrossRef] [PubMed]
- Shen, A. Clostridium difficile Toxins: Mediators of Inflammation. J. Innate. Immun. 2012, 4, 149–158. [Google Scholar] [CrossRef]
- Hensgens, C.M.H.; de Groot, N. Process for producing milk fractions rich in secretory immunoglobulins. EP Patent 2280999A1, 11 May 2009. [Google Scholar]
- Heidebrecht, H.-J.; Kainz, B.; Schopf, R.; Godl, K.; Karcier, Z.; Kulozik, U.; Förster, B. Isolation of biofunctional bovine immunoglobulin G from milk- and colostral whey with mixed-mode chromatography at lab and pilot scale. J. Chromatogr. A 2018, 1562, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Warny, M.; Fatimi, A.; Bostwick, E.F.; Laine, D.C.; Lebel, F.; LaMont, J.T.; Pothoulakis, C.; Kelly, C.P. Bovine immunoglobulin concentrate-Clostridium difficile retains C difficile toxin neutralising activity after passage through the human stomach and small intestine. Gut 1999, 44, 212–217. [Google Scholar] [CrossRef]
- Marx, M.; Kulozik, U. Spore inactivation in differently composed whey concentrates. Int. Dairy J. 2018, 76, 1–9. [Google Scholar] [CrossRef]
- Moos, M.; von Eichel-Streiber, C. Purification and evaluation of large clostridial cytotoxins that inhibit small GTPases of Rho and ras subfamilies. Methods Enzymol. 2000, 325, 114–125. [Google Scholar] [PubMed]
- Kaplan, E.L.; Meier, P. Nonparametric Estimation from Incomplete Observations. J. Am. Stat. Assoc. 1958, 53, 457. [Google Scholar] [CrossRef]
- Dunn, J.; Wild, D. Calibration Curve Fitting. In The Immunoassay Handbook: Theory and Applications of Ligand Binding, ELISA and Related Techniques; Wild, D., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 323–336. [Google Scholar]
- Lagkouvardos, I.; Fischer, S.; Kumar, N.; Clavel, T. Rhea: A transparent and modular R pipeline for microbial profiling based on 16S rRNA gene amplicons. PeerJ 2017, 5, e2836. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar]
- Benjamini, Y.; Yosef, H. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Series B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | WPI 10,000 | WPI 1000 | WPI 100 | Control-WPI |
|---|---|---|---|---|
| Total IgG [mg/mL] | 57.7 | 3.5 | 1.55 | 3.81 |
| Total IgA [mg/mL] | 3.01 | 0.43 | 0.17 | 0.43 |
| Total IgM [mg/mL] | 5.81 | 0.89 | 0.39 | 0.81 |
| Total protein [%] | 96.78 | 91.24 | 91.24 | 90.1 |
| Dry matter [%] | 98.09 | 94.15 | 94.15 | 92.51 |
| EC50 | WPI 10,000 | WPI 1000 | WPI 100 | Control-WPI |
|---|---|---|---|---|
| IgG TcdA | 6793 ± 291 | NA | NA | NA |
| IgG TcdB | 12665 ± 1312 | NA | NA | NA |
| IgG C. difficile | 12445 ± 124 | NA | NA | NA |
| IgA TcdA | 39882 ± 2737 | 428 ± 17 | 230 ± 47 | NA |
| IgA TcdB | 55708 ± 6601 | 578 ± 11 | 480 ± 18 | NA |
| IgA C. difficile | 38222 ± 7014# | 2021 ± 292 | 516 ± 146 | 235 ± 51 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heidebrecht, H.-J.; Weiss, W.J.; Pulse, M.; Lange, A.; Gisch, K.; Kliem, H.; Mann, S.; Pfaffl, M.W.; Kulozik, U.; von Eichel-Streiber, C. Treatment and Prevention of Recurrent Clostridium difficile Infection with Functionalized Bovine Antibody-Enriched Whey in a Hamster Primary Infection Model. Toxins 2019, 11, 98. https://doi.org/10.3390/toxins11020098
Heidebrecht H-J, Weiss WJ, Pulse M, Lange A, Gisch K, Kliem H, Mann S, Pfaffl MW, Kulozik U, von Eichel-Streiber C. Treatment and Prevention of Recurrent Clostridium difficile Infection with Functionalized Bovine Antibody-Enriched Whey in a Hamster Primary Infection Model. Toxins. 2019; 11(2):98. https://doi.org/10.3390/toxins11020098
Chicago/Turabian StyleHeidebrecht, Hans-Jürgen, William J Weiss, Mark Pulse, Anton Lange, Karina Gisch, Heike Kliem, Sacha Mann, Michael W. Pfaffl, Ulrich Kulozik, and Christoph von Eichel-Streiber. 2019. "Treatment and Prevention of Recurrent Clostridium difficile Infection with Functionalized Bovine Antibody-Enriched Whey in a Hamster Primary Infection Model" Toxins 11, no. 2: 98. https://doi.org/10.3390/toxins11020098
APA StyleHeidebrecht, H. -J., Weiss, W. J., Pulse, M., Lange, A., Gisch, K., Kliem, H., Mann, S., Pfaffl, M. W., Kulozik, U., & von Eichel-Streiber, C. (2019). Treatment and Prevention of Recurrent Clostridium difficile Infection with Functionalized Bovine Antibody-Enriched Whey in a Hamster Primary Infection Model. Toxins, 11(2), 98. https://doi.org/10.3390/toxins11020098