Differentiated Effects of Secondary Metabolites from Solanaceae and Brassicaceae Plant Families on the Heartbeat of Tenebrio molitor Pupae

 , ,
, ,  ,
,  ,
,  ,
, 
 and
and 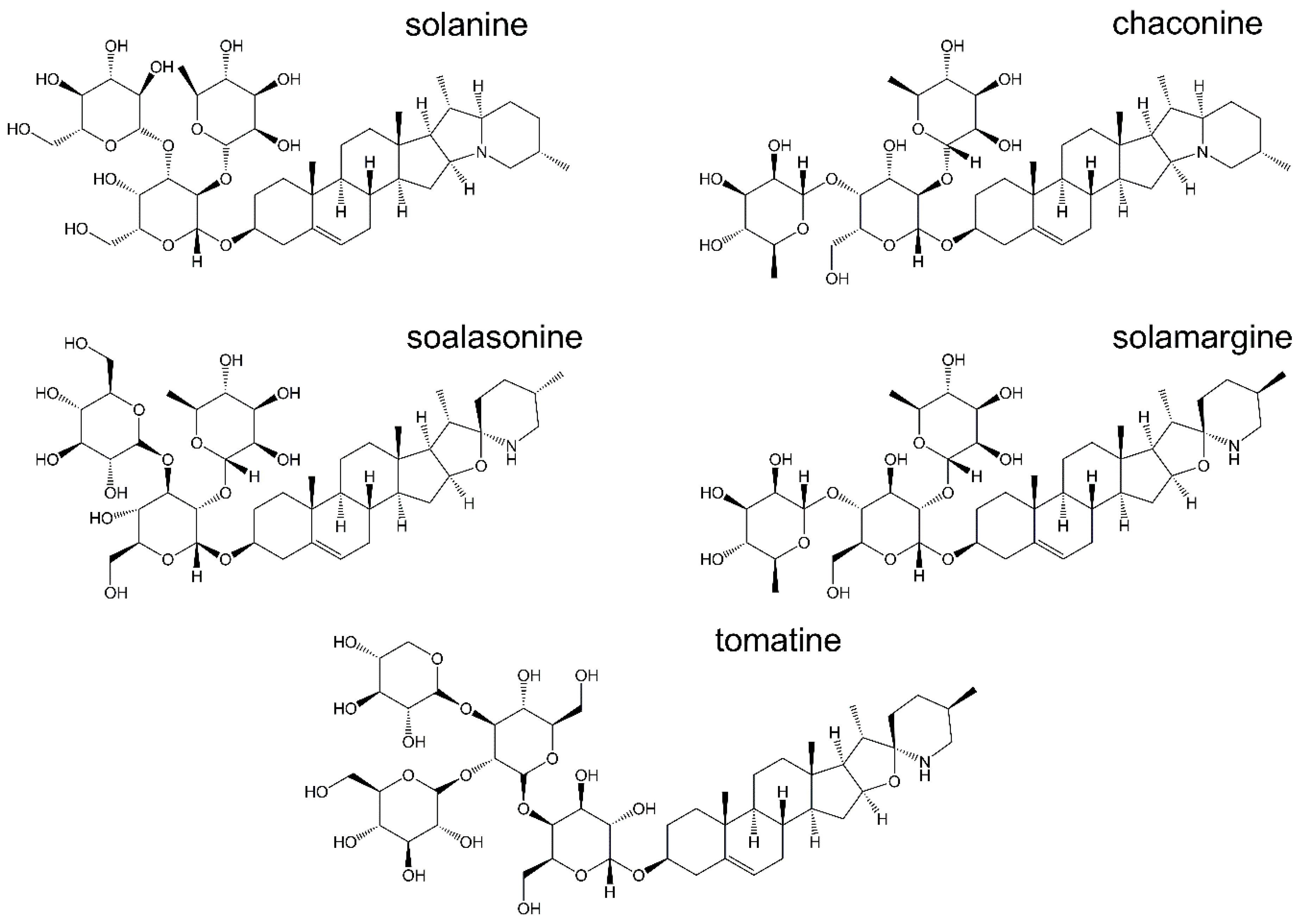{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
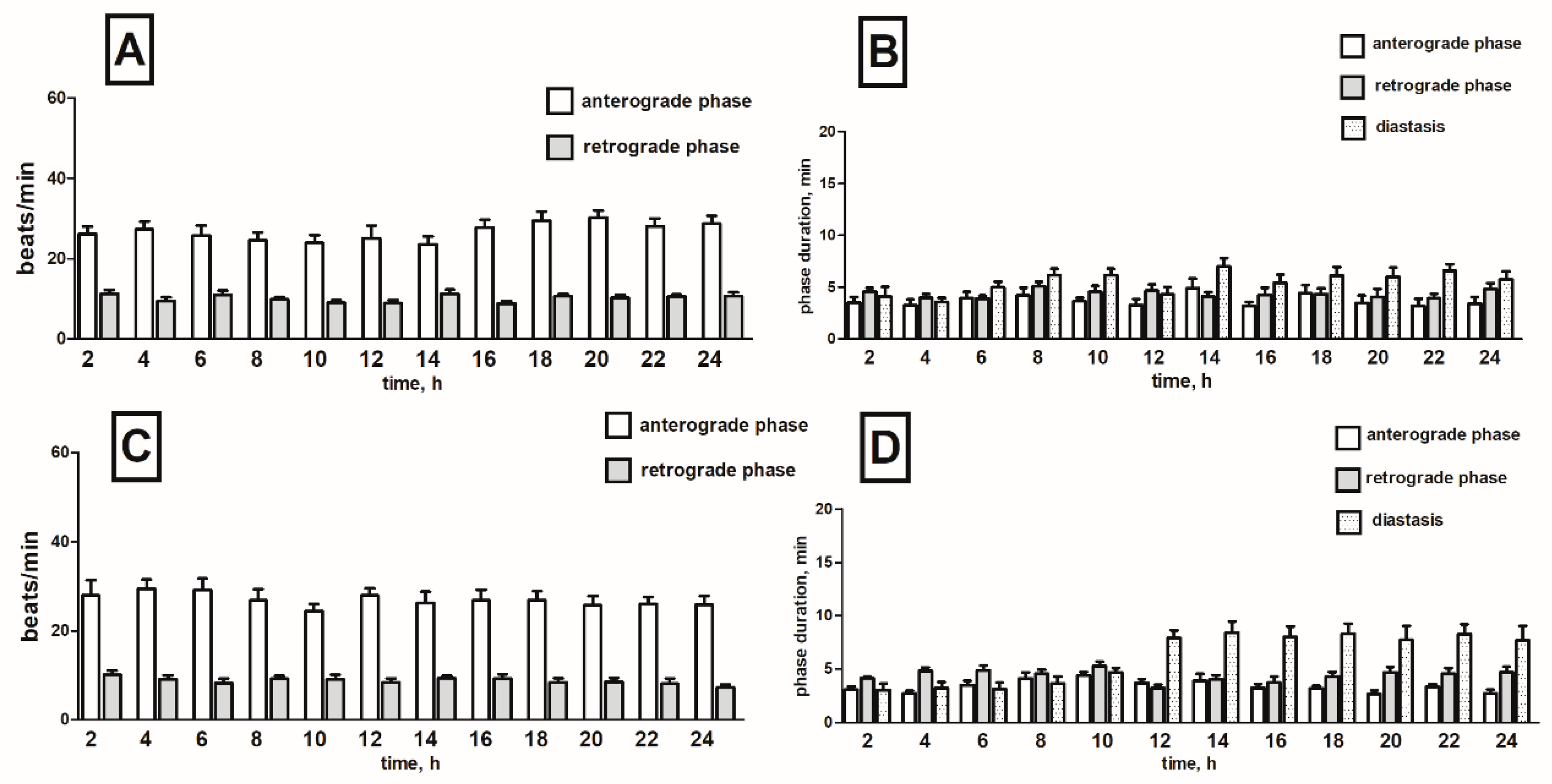
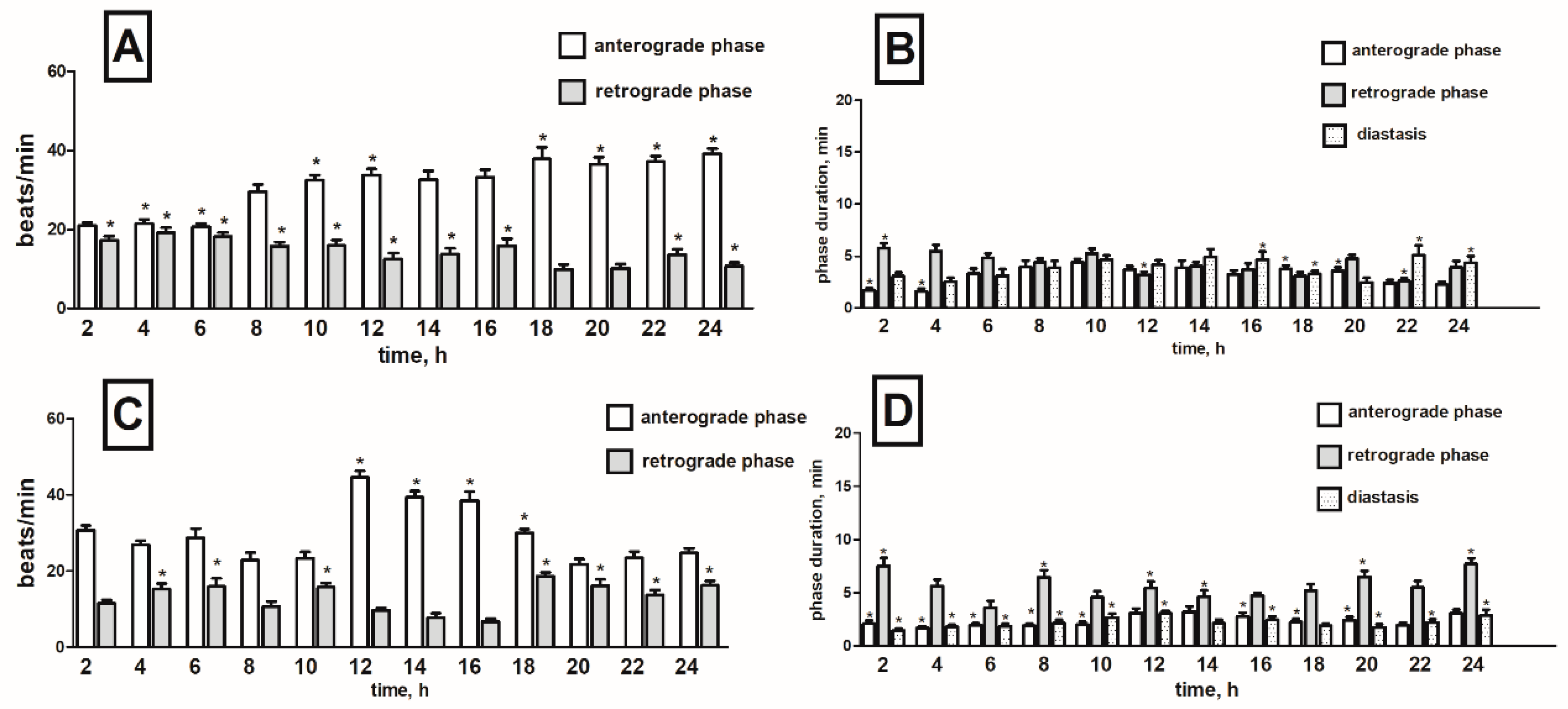
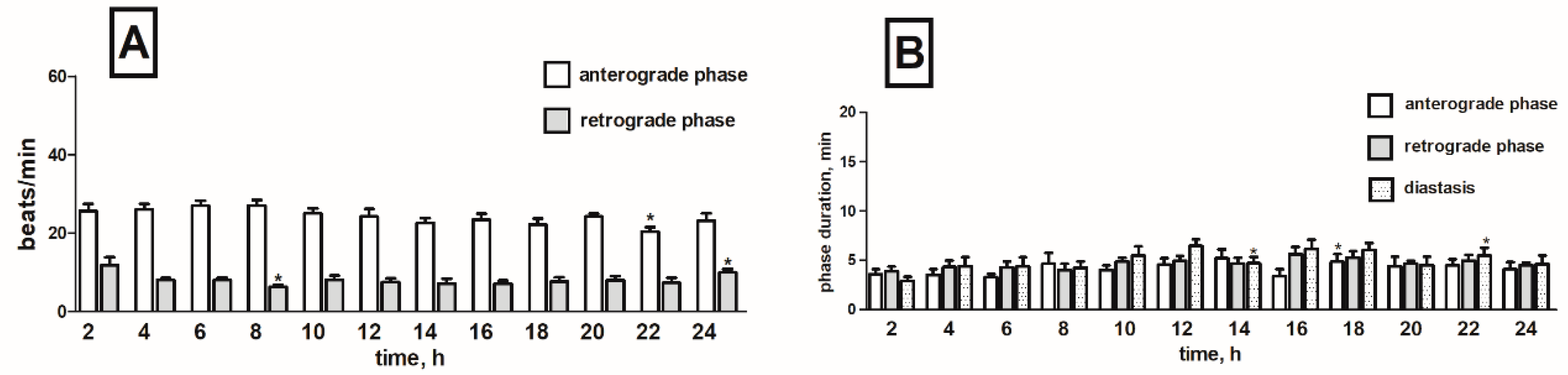
2.1. Heart Rhythm of the Tenebrio molitor Pupae
2.2. Effects of Potato Glycoalkaloids on the Heart Rhythm of T. molitor Pupae
2.2.1. Pure Alkaloids
α-Solanine
α-Chaconine
2.2.2. S. tuberosum Extract
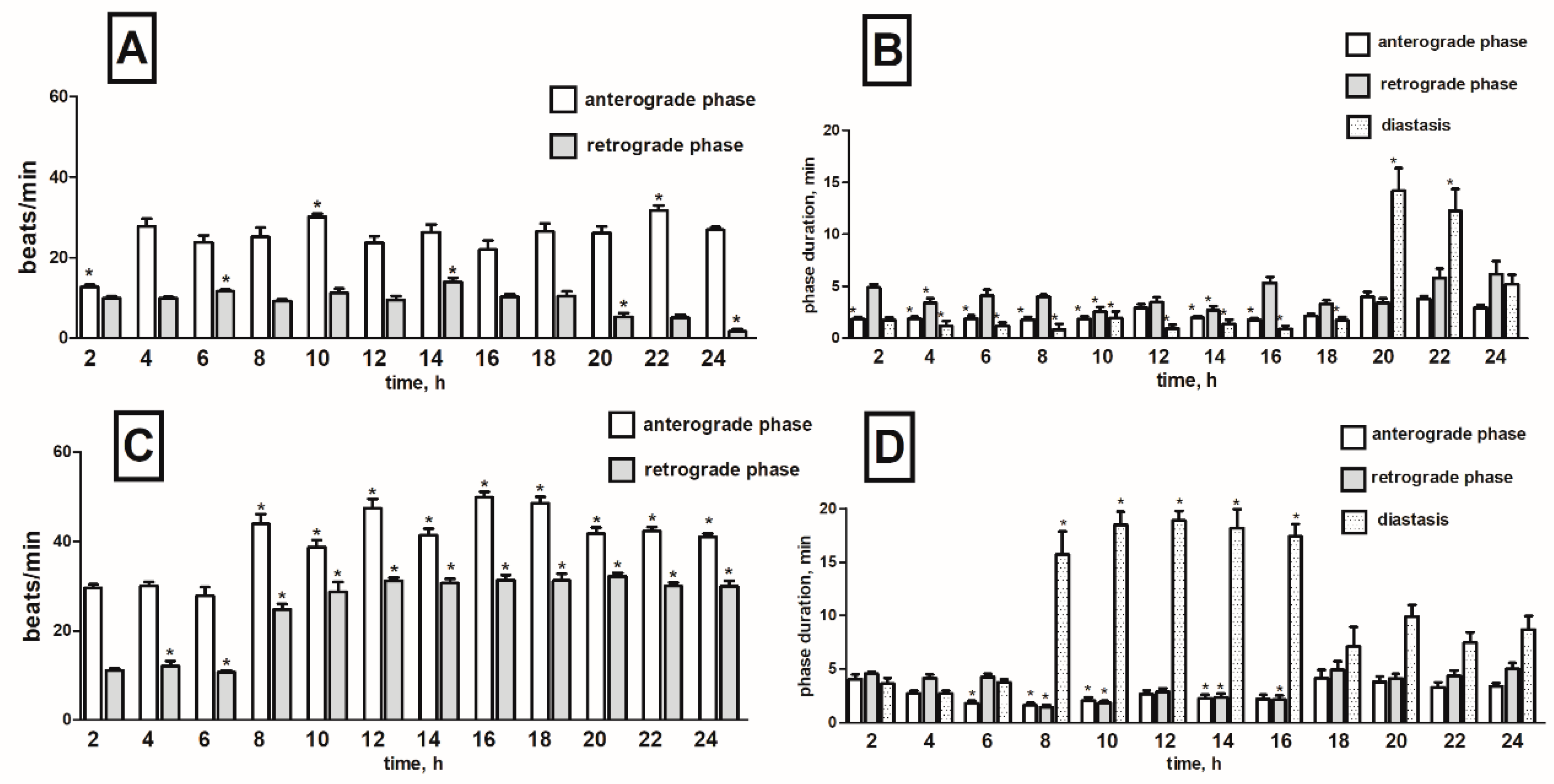
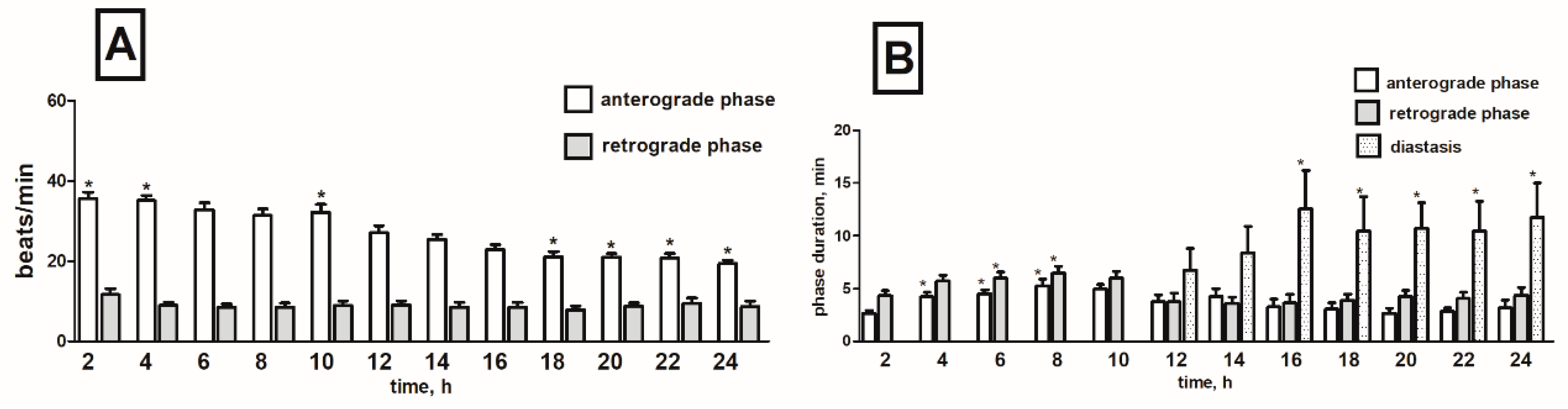
2.3. Effects of Black Nightshade Glycoalkaloids on the Heart Rhythm of T. molitor Pupae
2.3.1. Pure Alkaloids
Solamargine
Solasonine
2.3.2. S. nigrum Extract
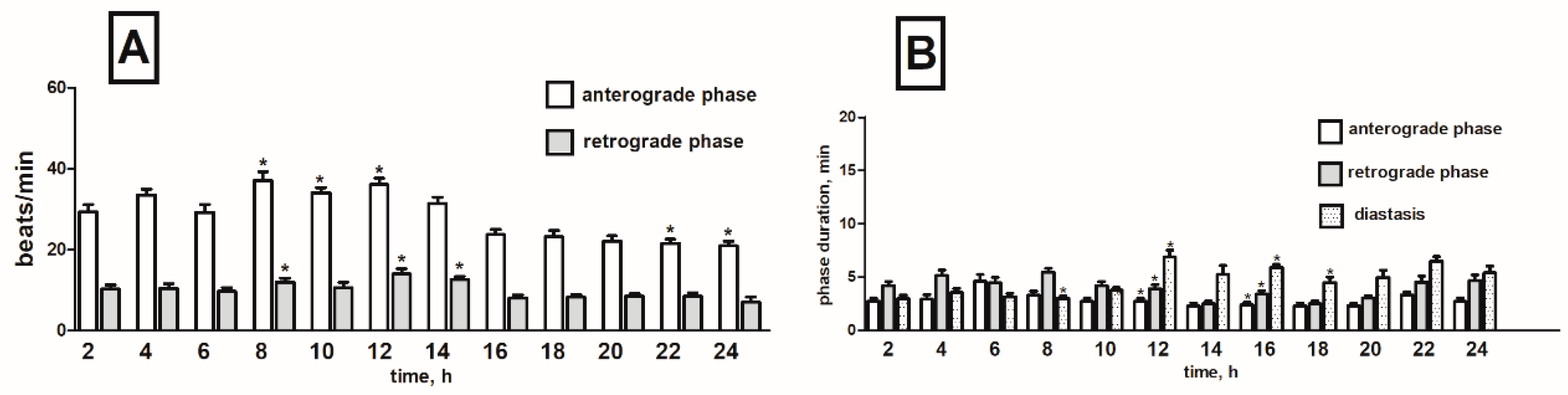
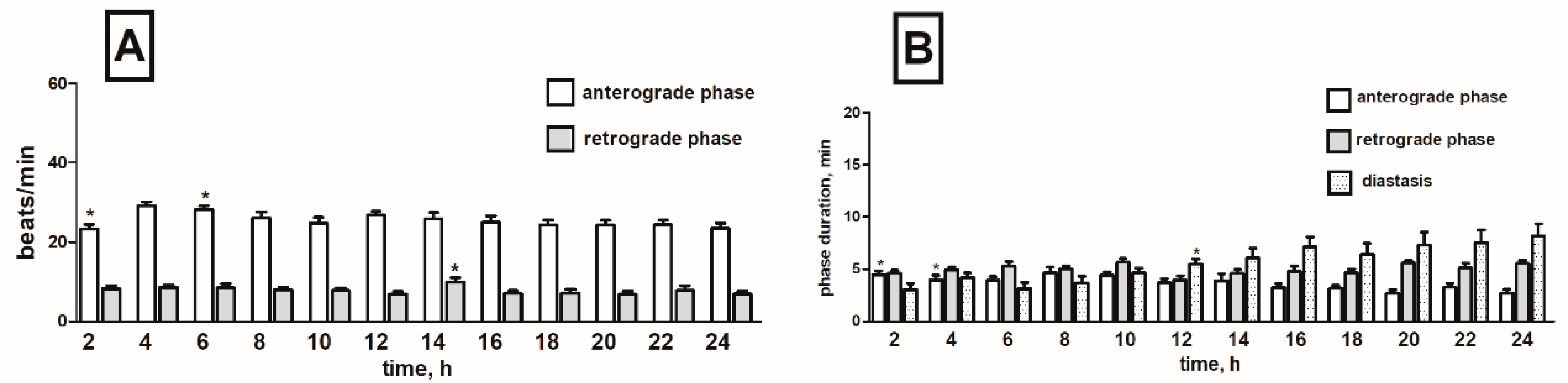
2.4. Effects of Tomato Glycoalkaloids on the Heart Rhythm of T. molitor Pupae
2.4.1. Synthetic α-Tomatine
2.4.2. S. lycopersicum Extract
2.5. Effects of Horseradish Glucosinolates on the Heart Rhythm of T. molitor Pupae
3. Discussion
4. Materials and Methods
4.1. Insects
4.2. Chemical Standards
4.3. Plant Material and Extracts Preparation
4.4. In Vivo Heart Bioassay
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shitan, N. Secondary metabolites in plants: Transport and self-tolerance mechanisms. Biosci. Biotechnol. Biochem. 2016, 80, 1283–1293. [Google Scholar] [CrossRef]
- Habli, Z.; Toumieh, G.; Fatfat, M.; Rahal, O.N.; Gali-Muhtasib, H. Emerging Cytotoxic Alkaloids in the Battle against Cancer: Overview of Molecular Mechanisms. Molecules 2017, 22, 250. [Google Scholar] [CrossRef]
- Chrobot, A.; Matkowski, A. Plant natural drugs as calcium channel blockers in the cardiovascular system. Postępy Fitoter. 2007, 2, 95–108. [Google Scholar]
- Lichota, A.; Gwozdzinski, K. Anticancer Activity of Natural Compounds from Plant and Marine Environment. Int. J. Mol. Sci. 2018, 19, 3533. [Google Scholar] [CrossRef]
- Roddick, J.G. Steroidal glycoalkaloids: Nature and consequences of bioactivity. Adv. Exp. Med. Biol. 1996, 404, 277–295. [Google Scholar]
- Chowanski, S.; Adamski, Z.; Marciniak, P.; Rosinski, G.; Buyukguzel, E.; Buyukguzel, K.; Falabella, P.; Scrano, L.; Ventrella, E.; Lelario, F.; et al. A Review of Bioinsecticidal Activity of Solanaceae Alkaloids. Toxins 2016, 8, 60. [Google Scholar] [CrossRef]
- Agneta, R.; Rivelli, A.R.; Ventrella, E.; Lelario, F.; Sarli, G.; Bufo, S.A. Investigation of glucosinolate profile and qualitative aspects in sprouts and roots of horseradish (Armoracia rusticana) using LC-ESI-hybrid linear ion trap with Fourier transform ion cyclotron resonance mass spectrometry and infrared multiphoton dissociation. J. Agric. Food Chem. 2012, 60, 7474–7482. [Google Scholar] [CrossRef]
- Spochacz, M.; Chowanski, S.; Walkowiak-Nowicka, K.; Szymczak, M.; Adamski, Z. Plant-Derived Substances Used Against Beetles-Pests of Stored Crops and Food-and Their Mode of Action: A Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1339–1366. [Google Scholar] [CrossRef] [Green Version]
- Chowanski, S.; Adamski, Z.; Lubawy, J.; Marciniak, P.; Pacholska-Bogalska, J.; Slocinska, M.; Spochacz, M.; Szymczak, M.; Urbanski, A.; Walkowiak-Nowicka, K.; et al. Insect Peptides-Perspectives in Human Diseases Treatment. Curr. Med. Chem. 2017, 24, 3116–3152. [Google Scholar] [CrossRef]
- Adamski, Z.; Bufo, S.A.; Chowanski, S.; Falabella, P.; Lubawy, J.; Marciniak, P.; Pacholska-Bogalska, J.; Salvia, R.; Scrano, L.; Slocinska, M.; et al. Beetles as Model Organisms in Physiological, Biomedical and Environmental Studies—A Review. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Ventrella, E.; Marciniak, P.; Adamski, Z.; Rosinski, G.; Chowanski, S.; Falabella, P.; Scrano, L.; Bufo, S.A. Cardioactive properties of Solanaceae plant extracts and pure glycoalkaloids on Zophobas atratus. Insect Sci. 2015, 22, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Szymczak, M.; Marciniak, P.; Rosiński, G. Insect myocardium-model to biomedical research. Adv. Cell Biol. 2014, 41, 59–77. [Google Scholar]
- Bier, E.; Bodmer, R. Drosophila, an emerging model for cardiac disease. Gene 2004, 342, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ocorr, K.; Akasaka, T.; Bodmer, R. Age-related cardiac disease model of Drosophila. Mech. Ageing Dev. 2007, 128, 112–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slama, K.; Rosinski, G. Delayed pharmacological effects of proctolin and CCAP on heartbeat in pupae of the tobacco hornworm, Manduca sexta. Physiol. Entomol. 2005, 30, 14–28. [Google Scholar] [CrossRef]
- Pacholska-Bogalska, J.; Szymczak, M.; Marciniak, P.; Walkowiak-Nowicka, K.; Rosinski, G. Heart mechanical and hemodynamic parameters of a beetle, Tenebrio molitor, at selected ages. Arch. Insect Biochem. Physiol. 2018, 99. [Google Scholar] [CrossRef] [PubMed]
- Ejaz, A.; Lange, A.B. Peptidergic control of the heart of the stick insect, Baculum extradentatum. Peptides 2008, 29, 214–225. [Google Scholar] [CrossRef]
- Al Chami, L.; Mendez, R.; Chataing, B.; O’Callaghan, J.; Usubillaga, A.; LaCruz, L. Toxicological effects of alpha-solamargine in experimental animals. Phytother. Res. 2003, 17, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Bergers, W.W.A.; Alink, G.M. Toxic Effect of the Glycoalkaloids Solanine and Tomatine on Cultured Neonatal Rat-Heart Cells. Toxicol. Lett. 1980, 6, 29–32. [Google Scholar] [CrossRef]
- Nishie, K.; Fitzpatrick, T.J.; Swain, A.P.; Keyl, A.C. Positive Inotropic Action of Solanaceae Glycoalkaloids. Res. Commun. Chem. Pathol. Pharmacol. 1976, 15, 601–607. [Google Scholar]
- Marciniak, P.; Adamski, Z.; Bednarz, P.; Slocinska, M.; Ziemnicki, K.; Lelario, F.; Scrano, L.; Bufo, S.A. Cardioinhibitory properties of potato glycoalkaloids in beetles. Bull. Environ. Contam. Toxicol. 2010, 84, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Spochacz, M.; Chowanski, S.; Szymczak, M.; Lelario, F.; Bufo, S.A.; Adamski, Z. Sublethal Effects of Solanum nigrum Fruit Extract and Its Pure Glycoalkaloids on the Physiology of Tenebrio molitor (Mealworm). Toxins 2018, 10, 504. [Google Scholar] [CrossRef]
- Chowanski, S.; Chudzinska, E.; Lelario, F.; Ventrella, E.; Marciniak, P.; Miadowicz-Kobielska, M.; Spochacz, M.; Szymczak, M.; Scrano, L.; Bufo, S.A.; et al. Insecticidal properties of Solanum nigrum and Armoracia rusticana extracts on reproduction and development of Drosophila melanogaster. Ecotoxicol. Environ. Saf. 2018, 162, 454–463. [Google Scholar] [CrossRef]
- Adamski, Z.; Radtke, K.; Kopiczko, A.; Chowanski, S.; Marciniak, P.; Szymczak, M.; Spochacz, M.; Falabella, P.; Lelario, F.; Scrano, L.; et al. Ultrastructural and developmental toxicity of potato and tomato leaf extracts to beet armyworm, Spodoptera exigua (lepidoptera: Noctuidae). Microsc. Res. Tech. 2016, 79, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Adamski, Z.; Adamski, Z.; Marciniak, P.; Ziemnicki, K.; Buyukguzel, E.; Erdem, M.; Buyukguzel, K.; Ventrella, E.; Falabella, P.; Cristallo, M.; et al. Potato leaf extract and its component, alpha-solanine, exert similar impacts on development and oxidative stress in Galleria mellonella L. Arch. Insect Biochem. Physiol. 2014, 87, 26–39. [Google Scholar] [CrossRef]
- Wang, X.; Zou, S.; Lan, Y.L.; Xing, J.S.; Lan, X.Q.; Zhang, B. Solasonine inhibits glioma growth through anti-inflammatory pathways. Am. J. Transl. Res. 2017, 9, 3977–3989. [Google Scholar]
- Zhang, X.H.; Yan, Z.P.; Xu, T.T.; An, Z.T.; Chen, W.Z.; Wang, X.S.; Huang, M.M.; Zhu, F.S. Solamargine derived from Solanum nigrum induces apoptosis of human cholangiocarcinoma QBC939 cells. Oncol. Lett. 2018, 15, 6329–6335. [Google Scholar] [CrossRef]
- Abellán, Á.; Domínguez-Perles, R.; Moreno, D.A.; García-Viguera, C. Sorting out the value of cruciferous sprouts as sources of bioactive compounds for nutrition and health. Nutrients 2019, 11, 429. [Google Scholar] [CrossRef]
- Chaudhary, P.; Sharma, A.; Singh, B.; Nagpal, A.K. Bioactivities of phytochemicals present in tomato. J. Food. Sci. Technol. 2018, 55, 2833–2849. [Google Scholar] [CrossRef]
- Rosinski, G.; Pilc, L.; Obuchowicz, L. Effect of Hydrocortisone on Growth and Development of Larvae Tenebrio-Molitor. J. Insect Physiol. 1978, 24, 97–99. [Google Scholar] [CrossRef]
- Cataldi, T.R.; Lelario, F.; Bufo, S.A. Analysis of tomato glycoalkaloids by liquid chromatography coupled with electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 3103–3110. [Google Scholar] [CrossRef]
- Agneta, R.; Lelario, F.; De Maria, S.; Mollers, C.; Bufo, S.A.; Rivelli, A.R. Glucosinolate profile and distribution among plant tissues and phenological stages of field-grown horseradish. Phytochemistry 2014, 106, 178–187. [Google Scholar] [CrossRef]
- Chowanski, S.; Rosinski, G. Myotropic Effects of Cholinergic Muscarinic Agonists and Antagonists in the Beetle Tenebrio molitor L. Curr. Pharm. Biotechnol. 2017, 18, 1088–1097. [Google Scholar] [CrossRef] [PubMed]










© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marciniak, P.; Kolińska, A.; Spochacz, M.; Chowański, S.; Adamski, Z.; Scrano, L.; Falabella, P.; Bufo, S.A.; Rosiński, G. Differentiated Effects of Secondary Metabolites from Solanaceae and Brassicaceae Plant Families on the Heartbeat of Tenebrio molitor Pupae. Toxins 2019, 11, 287. https://doi.org/10.3390/toxins11050287
Marciniak P, Kolińska A, Spochacz M, Chowański S, Adamski Z, Scrano L, Falabella P, Bufo SA, Rosiński G. Differentiated Effects of Secondary Metabolites from Solanaceae and Brassicaceae Plant Families on the Heartbeat of Tenebrio molitor Pupae. Toxins. 2019; 11(5):287. https://doi.org/10.3390/toxins11050287
Chicago/Turabian StyleMarciniak, Paweł, Angelika Kolińska, Marta Spochacz, Szymon Chowański, Zbigniew Adamski, Laura Scrano, Patrizia Falabella, Sabino A. Bufo, and Grzegorz Rosiński. 2019. "Differentiated Effects of Secondary Metabolites from Solanaceae and Brassicaceae Plant Families on the Heartbeat of Tenebrio molitor Pupae" Toxins 11, no. 5: 287. https://doi.org/10.3390/toxins11050287
APA StyleMarciniak, P., Kolińska, A., Spochacz, M., Chowański, S., Adamski, Z., Scrano, L., Falabella, P., Bufo, S. A., & Rosiński, G. (2019). Differentiated Effects of Secondary Metabolites from Solanaceae and Brassicaceae Plant Families on the Heartbeat of Tenebrio molitor Pupae. Toxins, 11(5), 287. https://doi.org/10.3390/toxins11050287