Mitigating Aflatoxin Contamination in Groundnut through A Combination of Genetic Resistance and Post-Harvest Management Practices


 ,
, 
 ,
,  , , ,
, , , 

Abstract
:1. Introduction
2. Characterization of Aflatoxin Producing Pathogens
3. Adverse Impacts of Aflatoxin Contamination on Human Health and the Economy
4. Current Understanding of Resistance to Aflatoxin Contamination Based on Genetic, Genomic, Transcriptomic and Proteomic Studies
5. Pathways Impacting Host-Pathogen Interaction and Toxin Production
6. Integrated Approach for Discovering Genomic Regions and Candidate Genes
7. Moving Towards Genomics-Assisted and Transgenic-Based Genetic Improvement to Confer Aflatoxin Resistance
8. A Mix of Genetic Resistance, Effective Post-Harvest Management Practices and Safe Storage
9. Challenges and Opportunities
Funding
Acknowledgments
Conflicts of Interest
References
- Kurtzman, C.P.; Smiley, M.J.; Robnett, C.J.; Wicklow, D.T. DNA relatedness among wild and domesticated species in the Aspergillus flavus group. Mycologia 1986, 78, 955–959. [Google Scholar] [CrossRef]
- Payne, G.A. Aflatoxin in maize. Crit. Rev. Plant Sci. 1992, 10, 423–440. [Google Scholar] [CrossRef]
- Goto, T.; Wicklow, D.T.; Ito, Y. Aflatoxin and cyclopiazonic acid production by a sclerotium-producing Aspergillus tamarii strain. Appl. Environ. Microbiol. 1996, 62, 4036–4038. [Google Scholar] [PubMed]
- Severns, D.E.; Clements, M.J.; Lambert, R.J.; White, D.G. Comparison of Aspergillus ear rot and aflatoxin contamination in grain of high-oil and normal oil corn hybrids. J. Food Prot. 2003, 66, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Roebuck, B.D.; Wogan, G.N.; Groopman, J.D. Aflatoxin: A 50-year odyssey of mechanistic and translational toxicology. Toxicol. Sci. 2011, 120, S28–S48. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.; Min, W.K.; Kweon, D.H.; Park, K.; Park, Y.C. Characterization of monoclonal antibody against aflatoxin B(1) produced in hybridoma 2C12 and its single-chain variable fragment expressed in recombinant Escherichia coli. Food Chem. 2011, 126, 1316–1323. [Google Scholar]
- Awasthi, V.; Bahman, S.; Thakur, L.K.; Singh, S.K.; Dua, A.; Ganguly, S. Contaminants in milk and impact of heating: An assessment study. Indian J. Public Health. 2012, 56, 95. [Google Scholar] [PubMed]
- Hameed, H.G. Extrusion and Chemical Treatments for Destruction of Aflatoxin in Naturally-Contaminated Corn. Ph.D. Thesis, University of Arizona, Tucson, AZ, USA, 1993. [Google Scholar]
- Saalia, F.K.; Phillips, R.D. Reduction of aflatoxins in peanut meal by extrusion cooking in the presence of nucleophiles. LWT Food Sci. Technol. 2011, 44, 1511–1516. [Google Scholar] [CrossRef]
- Torres, P.; Guzmán-Ortiz, M.; Ramírez-Wong, B. Revising the role of pH and thermal treatments in aflatoxin content reduction during the tortilla and deep frying processes. J. Agric. Food Chem. 2001, 6, 2825–2829. [Google Scholar] [CrossRef]
- Waliyar, F.; Ba, A.; Hassan, H.; Bonkoubgou, S.; Bose, J.P. Sources of resistance to aspergillus flavus and aflatoxin contamination in groundnut genotypes in West Africa. Plant Dis. 1992, 78, 704–708. [Google Scholar] [CrossRef]
- Thakur, R.P.; Rao, V.P.; Reddy, S.V.; Ferguson, M. Evaluation of wild Arachis germplasm accessions for in vitro seed colonization and aflatoxin production by Aspergillus flavus. Int. Arachis Newslett. 2000, 20, 44–46. [Google Scholar]
- Olwari, F.; Bisikwa, J.; Kaaya, A.N.; Okello, D.K. Tolerance levels of peanut varieties against Aspergillus flavus infection. J. Plant Pathol. Microb. 2013, 4, 195. [Google Scholar]
- Korani, W.A.; Chu, Y.; Holbrook, C.; Clevenger, J.; Ozias-Akins, P. Genotypic regulation of aflatoxin accumulation but not aspergillus fungal growth upon post-harvest infection of peanut (Arachis hypogaea L.) Seeds. Toxins 2017, 9, 218. [Google Scholar] [CrossRef] [PubMed]
- Dieme, R.M.A.; Faye, I.; Zoclanclounon, Y.A.B.; Fonceka, D.; Ndoye, O.; Diedhiou, P.M. Identification of sources of resistance for Peanut Aspergillus flavus colonization and Aflatoxin contamination. Int. J. Agron. 2018, 2018, 5468602. [Google Scholar] [CrossRef]
- Pandey, M.K.; Monyo, E.; Ozias-Akins, P.; Liang, X.; Guimarães, P.; Nigam, S.N.; Upadhyaya, H.D.; Janila, P.; Zhang, X.; Guo, B.; et al. Advances in Arachis genomics for peanut improvement. Biotechnol. Adv. 2012, 30, 639–651. [Google Scholar] [CrossRef]
- Pandey, M.K.; Roorkiwal, M.; Singh, V.K.; Ramalingam, A.; Kudapa, H.; Thudi, M.; Chitikineni, A.; Rathore, A.; Varshney, R.K. Emerging genomic tools for legume breeding: Current status and future prospects. Front. Plant Sci. 2016, 7, 455. [Google Scholar] [CrossRef]
- Varshney, R.K.; Pandey, M.K.; Bohra, A.; Singh, V.K.; Thudi, M.; Saxena, R.K. Toward the sequence-based breeding in legumes in the post-genome sequencing era. Theor. Appl. Genet. 2019, 132, 797–816. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, H.; Pandey, M.K.; Yang, Q.; Wang, X.; Garg, V.; Li, H.; Chi, X.; Doddamani, D.; Hong, Y.; et al. Draft genome of the peanut A-genome progenitor (Arachis duranensis) provides insights into geocarpy, oil biosynthesis, and allergens. Proc. Natl. Acad. Sci. USA 2016, 113, 6785–6790. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Cannon, S.B.; Froenicke, L.; Huang, G.; Farmer, A.D.; Cannon, E.K.; Liu, X.; Gao, D.; Clevenger, J.; Dash, S.; et al. The genome sequences of Arachis duranensis and Arachis ipaensis, the diploid ancestors of cultivated peanut. Nat. Genet. 2016, 48, 438–446. [Google Scholar] [CrossRef]
- Lu, Q.; Li, H.; Hong, Y.; Zhang, G.; Wen, S.; Li, X.; Zhou, G.; Li, S.; Liu, H.; Liu, H.; et al. Genome sequencing and analysis of the peanut B-genome progenitor (Arachis ipaensis). Front. Plant Sci. 2018, 9, 604. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.; Ji, C.; Ma, X.; Li, H.; Zhang, W.; Li, S.; Liu, F.; Zhao, K.; Li, F.; Li, K.; et al. Genome of an allotetraploid wild peanut Arachis monticola: A de novo assembly. GigaScience 2018, 7, giy066. [Google Scholar] [CrossRef] [PubMed]
- Bertioli, D.J.; Jenkins, J.; Clevenger, J.; Gao, D.; Dudchenko, O.; Seijo, G.; Leal-Bertioli, S.; Ren, L.; Farmer, A.; Pandey, M.K.; et al. The genome sequence of peanut (Arachis hypogaea), a segmental allotetraploid. Nat. Genet. 2019. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, Q.; Liu, H.; Zhang, J.; Hong, Y.; Lan, H.; Li, H.; Wang, J.; Liu, H.; Li, S.; et al. Sequencing of cultivated peanut, Arachis hypogaea, yields insights into genome evolution and oil improvement. Mol. Plant. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.; Chen, H.; Yang, M.; Wang, J.; Pandey, M.K.; Zhang, C.; Chang, W.-C.; Zhang, L.; Zhang, X.; Tang, R.; et al. The Arachis hypogaea genome elucidates legume karyotypes, polyploid evolution and crop domestication. Nat. Genet. 2019. [Google Scholar] [CrossRef]
- Clevenger, J.; Chu, Y.; Arrais, G.L.; Maia, T.; Bertioli, D.; Leal-Bertioli, S.; Timper, P.; Holbrook, C.C.; Ozias-Akins, P. Gene expression profiling describes the genetic regulation of Meloidogyne arenaria resistance in Arachis hypogaea and reveals a candidate gene for resistance. Sci. Rep. 2017, 7, 1317. [Google Scholar] [CrossRef] [PubMed]
- Sinha, P.; Bajaj, P.; Pazhamala, P.T.; Nayak, S.N.; Pandey, M.K.; Chitikineni, A.; Huai, D.; Khan, A.W.; Desai, A.; Guo, B.; et al. VI NGGIBCI Conference on Crop Genomics: Present & Future; ICRISAT: Hyderabad, India, 7–8 December 2017. [Google Scholar]
- Pandey, M.K.; Agarwal, G.; Kale, S.M.; Clevenger, J.; Nayak, S.N.; Sriswathi, M.; Chitikineni, A.; Chavarro, C.; Chen, X.; Upadhyaya, H.D.; et al. Development and evaluation of a high density genotyping ‘Axiom_Arachis’ array with 58 K SNPs for accelerating genetics and breeding in groundnut. Sci. Rep. 2017, 7, 40577. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lei, Y.; Wan, L.; Yan, L.; Lv, J.; Dai, X.; Ren, X.; Guo, W.; Jiang, H.; Liao, B. Comparative transcript profiling of resistant and susceptible peanut post-harvest seeds in response to aflatoxin production by Aspergillus flavus. BMC Plant Biol. 2016, 1, 54. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.N.; Agarwal, G.; Pandey, M.K.; Sudini, H.K.; Jayale, A.S.; Purohit, S.; Desai, A.; Wan, L.; Guo, B.; Liao, B.; et al. Aspergillus favus infection triggered immune responses and host-pathogen cross-talks in groundnut during in-vitro seed colonization. Sci. Rep. 2017, 7, 9659. [Google Scholar] [CrossRef]
- Korani, W.; Chu, Y.; Holbrook, C.C.; Ozias-Akins, P. Insight into genes regulating postharvest aflatoxin contamination of tetraploid peanut from transcriptional profiling. Genetics 2018, 209, 143–156. [Google Scholar] [CrossRef]
- Arias, R.S.; Dang, P.M.; Sobolev, V.S. RNAi-mediated control of aflatoxins in peanut: Method to analyse mycotoxin production and transgene expression in the peanut/Aspergillus pathosystem. J. Vis. Exp. 2015, 106, e53398. [Google Scholar] [CrossRef]
- Sharma, K.K.; Pothana, A.; Prasad, K.; Shah, D.; Kaur, J.; Bhatnagar, D.; Chen, Z.Y.; Raruang, Y.; Cary, J.W.; Rajasekaran, K.; et al. Peanuts that keep aflatoxin at bay: A threshold that matters. Plant Biotechnol. J. 2018, 16, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Lakkireddy, K.; Kondapalli, K.; Rao, K.R.S. Aflatoxins in food and feed: The science of safe food. J. Food Sci. Technol. 2014, 2, 6–11. [Google Scholar]
- Ehrlich, K.C. Non-aflatoxigenic Aspergillus flavus to prevent aflatoxin contamination in crops: Advantages and limitations. Front. Microbiol. 2014, 5, 50. [Google Scholar] [CrossRef] [PubMed]
- Horn, B.W.; Moore, G.G.; Carbone, I. Sexual reproduction in Aspergillus flavus. Mycologia 2009, 101, 423–429. [Google Scholar] [CrossRef]
- Horn, B.W.; Greene, R.L.; Dorner, J.W. Effect of corn and peanut cultivation on soil populations of Aspergillus flavus and Aspergillus parasiticus in southwestern Georgia. Appl. Environ. Microbiol. 1995, 61, 2472–2475. [Google Scholar]
- Horn, B.W.; Dorner, J.W. Soil population of Aspergillus species from section Flavi along a transect through peanut-growing regions of the United States. Mycologia 1998, 90, 767–776. [Google Scholar] [CrossRef]
- Molyneux, R.J.; Mahoney, N.; Kim, J.H.; Campbell, B.C. Mycotoxins in edible tree nuts. Int. J. Food Microbiol. 2007, 119, 72–78. [Google Scholar] [CrossRef]
- Cotty, P.J. Virulence and cultural characteristics of two Aspergillus flavus strains pathogenic on cotton. Phytopathologia 1989, 79, 808–814. [Google Scholar] [CrossRef]
- Miller, J.D. Mycotoxins in small grains and maize: Old problems, new challenges. Food Addit. Contam. 2008, 25, 219–230. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Nigam, S.N.; Thakur, R.P. Genetic enhancement for resistance to aflatoxin contamination in groundnut. In Summary proceedings of the Seventh ICRISAT Regional Groundnut Meeting for Western and Central Africa, Cotonu, Benin, 6–8 December 2000; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, India, 2002; pp. 29–36. [Google Scholar]
- Waliyar, F.; Ntare, B.R.; Diallo, A.T.; Kodio, O.; Diarra, B. On-Farm Management of Aflatoxin Contamination of Groundnut in West Africa: A Synthesis Report; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, India, 2007. [Google Scholar]
- Wilson, D.M.; Stansell, J.R. Effect of irrigation regimes on aflatoxin contamination of peanut pods. Peanut Sci. 1983, 10, 54–56. [Google Scholar] [CrossRef]
- Cole, R.J.; Sanders, T.H.; Hill, R.A.; Blankenship, P.D. Mean geocarposphere temperatures that induce pre-harvest aflatoxin contamination of peanuts under drought stress. Mycopathologia 1985, 91, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Girdthai, T.; Jogloy, S.; Vorasoot, N.; Akkasaeng, C.; Wongkaew, S.; Holbrook, C.C.; Patanothai, A. Associations between physiological traits for drought tolerance and aflatoxin contamination in peanut genotypes under terminal drought. Plant Breed. 2010, 129, 693–699. [Google Scholar] [CrossRef]
- Sudhakar, P.; Latha, P.; Babitha, M.; Reddy, P.V.; Naidu, P.H. Relationship of drought tolerance traits with aflatoxin contamination in groundnut. Indian J. Plant Physiol. 2007, 3, 261. [Google Scholar]
- Dorner, J.W.; Cole, R.J.; Sanders, T.H.; Blankenship, P.D. Interrelationship of kernel water activity, soil temperature, maturity, and phytoalexin production in preharvest aflatoxin contamination of drought-stressed peanuts. Mycopathologia 1989, 105, 117–128. [Google Scholar] [CrossRef]
- Waliyar, F.; Traore, D.; Fatondji, D.; Ntare, B.R. Effect of irrigation interval, planting date, and cultivar on Aspergillus flavus and aflatoxin contamination of peanut in a sandy soil of Niger. Peanut Sci. 2003, 30, 79–84. [Google Scholar] [CrossRef]
- Hamidou, F.; Rathore, A.; Waliyar, F.; Vadez, V. Although drought intensity increases aflatoxin contamination, drought tolerance does not lead to less aflatoxin contamination. Field Crops Res. 2014, 156, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus, G.H. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef]
- Buchanan, R.L.; Stahl, H.G. Ability of various carbon sources to induce and support aflatoxin synthesis by Aspergillus parasiticus. J. Food Saf. 1984, 6, 271–279. [Google Scholar] [CrossRef]
- Shih, C.N.; Marth, E.H. Aflatoxin formation, lipid synthesis, and glucose metabolism by Aspergillus parasiticus during incubation with and without agitation. Biochem. Biophys. Acta 1974, 338, 286–296. [Google Scholar] [CrossRef]
- Clevstrom, G.; Ljunggren, H.; Tegelstrom, S.; Tideman, K. Production of aflatoxin by an Aspergillus flavus isolate cultured under a limited oxygen supply. Appl. Environ. Microbiol. 1983, 46, 400–405. [Google Scholar]
- Watanabe, C.M.H.; Wilson, D.; Linz, J.E.; Townsend, C.A. Demonstration of the catalytic roles and evidence for the physical association of type I fatty acid synthases and a polyketide synthase in the biosynthesis of aflatoxin B1. Chem. Biol. 1996, 3, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Fountain, J.C.; Khera, P.; Yang, L.; Nayak, S.N.; Scully, B.T.; Lee, R.D.; Chen, Z.Y.; Kemerait, R.C.; Varshney, R.K.; Guo, B. Resistance to Aspergillus flavus in maize and peanut: Molecular biology, breeding, environmental stress, and future perspectives. Crop J. 2015, 3, 229–237. [Google Scholar] [CrossRef]
- Brown, D.W.; Yu, J.H.; Kelkar, H.S.; Fernandes, M.; Nesbitt, T.C.; Keller, N.P.; Adams, T.H.; Leonard, T.J. Twenty-five co-regulated transcripts define a sterigmatocystin gene cluster in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 1996, 93, 1418–1422. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.H.; Chu, F.S. Regulation of aflR and its product, AflR, associated with aflatoxin biosynthesis. Appl. Environ. Microbiol. 1998, 64, 3718–3723. [Google Scholar] [PubMed]
- Bhatnagar, D.; Ehrlich, K.C.; Cleveland, T.E. Molecular genetic analysis and regulation of aflatoxin biosynthesis. Appl. Microbiol. Biotechnol. 2003, 61, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Bhatnagar, D.; Cleveland, T.E. Completed sequence of aflatoxin pathway gene cluster in Aspergillus parasiticus. FEBS Lett. 2004, 564, 126–130. [Google Scholar] [CrossRef]
- Georgianna, D.R.; Payne, G.A. Genetic regulation of aflatoxin biosynthesis: From gene to genome. Fungal Genet. Biol. 2009, 46, 113–125. [Google Scholar] [CrossRef]
- Calvo, A.M. The VeA regulatory system and its role in morphological and chemical development in fungi. Fungal Genet. Biol. 2008, 45, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Strosnider, H.; Azziz-Baumgartner, E.; Banziger, M.; Bhat, R.V.; Breiman, R.; Brune, M.N.; DeCock, K.; Dilley, A.; Groopman, J.; Hell, K.; et al. Workgroup report: Public health strategies for reducing aflatoxin exposure in developing countries. Environ. Health Perspect. 2006, 114, 1898–1903. [Google Scholar] [CrossRef]
- Krishnamachari, K.A.; Bhat, R.V.; Nagarajan, V.; Tilak, T.B. Hepatitis due to aflatoxicosis. An outbreak in Western India. Lancet 1975, 1, 1061–1063. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, F. Global burden of aflatoxin-induced hepatocellular carcinoma: A risk assessment. Environ. Health Perspect. 2010, 118, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Valery, P.C.; Laversanne, M.; Clark, P.J.; Petrick, J.L.; McGlynn, K.A.; Bray, F. Projections of primary liver cancer to 2030 in 30 countries worldwide. Hepatology 2018, 67, 600–611. [Google Scholar] [CrossRef] [PubMed]
- Gopal, T.; Zaki, S.; Narayanaswami, M.; Premlata, S. Aflatoxicosis in fowls. Indian Vet. J. 1969, 46, 348–349. [Google Scholar]
- Blount, W.P. Turkey “X” disease. Turkeys 1961, 9, 52–67. [Google Scholar]
- Wannop, C.C. The histopathology of turkey “X” disease in Great Britain. Avian Dis. 1961, 5, 371–381. [Google Scholar] [CrossRef]
- Kelly, J.D.; Eaton, D.L.; Guengerich, F.P.; Coulombe, R.J. Aflatoxin B sub (1) activation in human lung. Toxicol. Appl. Pharmacol. 1997, 144, 88–95. [Google Scholar] [CrossRef]
- Wang, J.S.; Huang, T.; Su, J.; Liang, F.; Wei, Z.; Liang, Y.; Luo, H.; Kuang, S.Y.; Qian, G.S.; Sun, G.; et al. Hepatocellular carcinoma and aflatoxin exposure in Zhuqing village, Fusui county, People’s Republic of China. Cancer Epidemiol. Prev. Biomark. 2001, 10, 143–146. [Google Scholar]
- Tajkarimi, M.; Shojaee, M.H.; Yazdanpanah, H.; Ibrahim, S.A. Aflatoxin in agricultural commodities and herbal medicine. In Aflatoxins-Biochemistry and Molecular Biology; IntechOpen: London, UK, 2011. [Google Scholar] [CrossRef]
- Gbashi, S.; Madala, N.E.; Adekoya, I.; Adebo, O.; De Saeger, S.; De Boevre, M.; Njobeh, P.B. The socio-economic impact of mycotoxin contamination in Africa. In Fungi and Mycotoxins—Their Occurrence, Impact on Health and the Economy as well as Pre- and Postharvest Management Strategies; InTechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Coulibaly, O.; Hell, K.; Bandyopadhyay, R.; Hounkponou, S.; Leslie, J.F. Economic impact of aflatoxin contamination in Sub-Saharan Africa. In Mycotoxins Detection Methods, Management, Public Health and Agricultural Trade, 1st ed.; Leslie, J., Bandyopadhyay, R., Visconti, A., Eds.; CABI Publishing: Cambridge, UK, 2008; pp. 67–76. [Google Scholar]
- Hendrickse, R.G.; Coulter, J.B.; Lamplugh, S.M.; Macfarlane, S.B.; Williams, T.E.; Omer, M.I.; Suliman, G.I. Aflatoxins and kwashiorkor: A study in Sudanese children. Br. Med. J. (Clin. Res. Ed.) 1982, 285, 843–846. [Google Scholar] [CrossRef]
- Ramjee, G.; Berjak, P.; Adhikari, M.; Dutton, M.F. Aflatoxins and kwashiorkor in Durban, South Africa. Ann. Trop. Paediatr. 1992, 12, 241–247. [Google Scholar] [CrossRef]
- Lamplugh, S.M.; Hendrickse, R.G. Aflatoxins in the livers of children with kwashiorkor. Ann. Trop. Paediatr. 1982, 2, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Wagacha, J.M.; Muthomi, J.W. Mycotoxin problem in Africa: Current status, implications to food safety and health and possible management strategies. Int. J. Food Microbiol. 2008, 124, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gebrehiwet, Y.; Kirsten, J.; Qangweni, W. Quantifying the trade effect of sanitary and phytosanitary regulations of Organization for Economic Cooperation and Development (OECD) countries on South African food exports. Agrekon 2007, 46, 1–17. [Google Scholar] [CrossRef]
- Hall, A.J.; Wild, C.P. Epidemiology of aflatoxin-related disease. In The Toxicology of Aflatoxins: Human Health, Veterinary, and Agricultural Significance; Eaton, D.L., Groopman, J.D., Eds.; Academic Press: London, UK, 1993; pp. 233–258. [Google Scholar]
- Paterson, R.R.; Lima, N. Toxicology of mycotoxins. EXS 2010, 100, 31–63. [Google Scholar]
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human aflatoxicosis in developing countries: A review of toxicology, exposure, potential health consequences, and interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [CrossRef] [PubMed]
- Awuah, R.T.; Kpodo, K.A. High incidence of Aspergillus flavus and aflatoxins in stored groundnut in Ghana and the use of a microbial assay to assess the inhibitory effects of plant extracts on aflatoxin synthesis. Mycopathologia 1996, 134, 109–114. [Google Scholar] [CrossRef]
- Sharma, R.K.; Parisi, S. Toxins and Contaminants in Indian Food Products; Springer International Publishing: New York, NY, USA, 2017; pp. 13–24. [Google Scholar]
- Cooper, J.; Dobson, H. The benefits of pesticides to mankind and the environment. Crop Prot. 2007, 23, 1337–1348. [Google Scholar] [CrossRef]
- Lawley, T.D.; Walker, A.W. Intestinal colonization resistance. Immunology 2013, 138, 1–11. [Google Scholar] [CrossRef]
- Yu, B.; Huai, D.; Huang, L.; Kang, Y.; Ren, X.; Chen, Y.; Zhou, X.; Luo, H.; Liu, N.; Chen, W.; et al. Identification of genomic regions and diagnostic markers for resistance to aflatoxin contamination in peanut (Arachis hypogaea L.). BMC Genet. 2019, 20, 32. [Google Scholar] [CrossRef]
- Holbrook, C.C.; Wilson, D.M.; Matheron, M.E.; Hunter, J.E.; Knauft, D.A.; Gorbet, D.W. Aspergillus colonization and aflatoxin contamination in peanut genotypes with reduced linoleic acid composition. Plant Dis. 2000, 84, 148–150. [Google Scholar] [CrossRef]
- ICRISAT (International Crops Research Institute for the Semi-Arid Tropics). Aflatoxin contamination of groundnut: Proceedings of the international workshop, 6–9 October 1987; ICRISAT: Patancheru, India, 1987. [Google Scholar]
- ICRISAT Center. Aflatoxin contamination of groundnut: proceedings of the International Workshop. 6–9 October 1987, ICRISAT Center, India; ICRISAT: Patancheru, India, 1989. [Google Scholar]
- Mixon, A.C.; Rogers, K.M. Peanut accessions resistant to seed infection by Aspergillus flavus. Agron. J. 1973, 65, 560–562. [Google Scholar] [CrossRef]
- Kisyombe, C.T.; Beute, M.K.; Payne, G.A. Field evaluation of peanut genotypes for resistance to infection by Aspergillus parasiticus. Peanut Sci. 1985, 12, 12–17. [Google Scholar] [CrossRef]
- Anderson, W.F.; Holbrook, C.C.; Wilson, D.M.; Matheron, M.E. Evaluation of peanut aflatoxin contamination in several potentially resistant peanut genotypes. Peanut Sci. 1995, 22, 29–32. [Google Scholar] [CrossRef]
- Mehan, V.K.; McDonald, D. Research on the aflatoxin problem in groundnut at ICRISAT. Plant and Soil. 1979, 79, 255–260. [Google Scholar] [CrossRef]
- Jiang, H.F.; Ren, X.P.; Wang, S.Y.; Zhang, X.J.; Huang, J.Q.; Liao, B.S.; Holbrook, C.C.; Upadhyaya, H.D. Development and evaluation of peanut germplasm with resistance to Aspergillus flavus from core collection. Acta Agron. Sin. 2010, 3, 428–434. [Google Scholar] [CrossRef]
- Guo, B.; Fedorova, N.D.; Chen, X.; Wan, C.H.; Wang, W.; Nierman, W.C.; Bhatnagar, D.; Yu, J. Gene expression profiling and identification of resistance genes to Aspergillus flavus infection in peanut through EST and microarray strategies. Toxins 2011, 3, 737–753. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Chen, X.; Dang, P.; Scully, B.T.; Liang, X.; Holbrook, C.C.; Yu, J.; Culbreath, A.K. Peanut gene expression profiling in developing seeds at different reproduction stages during Aspergillus parasiticus infection. BMC Dev. Biol. 2008, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yan, S.; Liu, C.; Chen, F.; Wang, T. Proteomic analysis reveals an aflatoxin-triggered immune response in cotyledons of Arachis hypogaea infected with Aspergillus flavus. J. Proteome Res. 2012, 11, 2739–2753. [Google Scholar] [CrossRef] [PubMed]
- Bedre, R.; Rajasekaran, K.; Mangu, V.R.; Timm, L.E.; Bhatnagar, D.; Baisakh, N. Genome-wide transcriptome analysis of cotton (Gossypium hirsutum L.) identifies candidate gene signatures in response to aflatoxin producing fungus Aspergillus flavus. PLoS ONE 2015, 14, 0138025. [Google Scholar] [CrossRef]
- Wang, H.; Lei, Y.; Yan, L.; Cheng, K.; Dai, X.; Wan, L.; Guo, W.; Cheng, L.; Liao, B. Deep sequencing analysis of transcriptomes in Aspergillus flavus in response to resveratrol. BMC Microbiol. 2015, 15, 182. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, R.; Lebar, M.; Mack, B.; Minocha, R.; Minocha, S.; Carter-Wientjes, C.; Sickler, C.; Rajasekaran, K.; Cary, J.W. The Aspergillus flavus Spermidine synthase (spds) gene, is required for normal development, aflatoxin production, and pathogenesis during infection of maize kernels. Front. Plant Sci. 2018, 9, 317. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Brown, R.L.; Damann, K.E.; Cleveland, T.E. Identification of unique or elevated levels of kernel proteins in aflatoxin-resistant maize genotypes through proteome analysis. Phytopatholgia 2002, 92, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.L.; Chen, Z.Y.; Warburton, M.; Luo, M.; Menkir, A.; Fakhoury, A.; Bhatnagar, D. Discovery and characterization of proteins associated with aflatoxin-resistance: Evaluating their potential as breeding markers. Toxins 2010, 2, 919–933. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, E.; Chen, X.; Li, L.; Liang, X. Identification of seed proteins associated with resistance to pre-harvested aflatoxin contamination in peanut (Arachis hypogaea L). BMC Plant Biol. 2010, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Fountain, J.C.; Koh, J.; Yang, L.; Pandey, M.K.; Nayak, S.N.; Bajaj, P.; Zhuang, W.J.; Chen, Z.Y.; Kemerait, R.C.; Lee, R.D.; et al. Proteome analysis of Aspergillus flavus isolate-specific responses to oxidative stress in relationship to aflatoxin production capability. Sci. Rep. 2018, 8, 3430. [Google Scholar] [CrossRef] [PubMed]
- Thakare, D.; Zhang, J.; Wing, R.A.; Cotty, P.J.; Schmidt, M.A. Aflatoxin-free transgenic maize using host-induced gene silencing. Sci. Adv. 2017, 3, e1602382. [Google Scholar] [CrossRef] [PubMed]
- Clevenger, J.; Marasigan, K.; Liakos, V.; Sobolev, V.; Vellidis, G.; Holbrook, C.; Ozias-Akins, P. RNA sequencing of contaminated seeds reveals the state of the seed permissive for pre-harvest aflatoxin contamination and points to a potential susceptibility factor. Toxins 2016, 8, 317. [Google Scholar] [CrossRef]
- Wang, T.; Chen, X.; Li, H.; Liu, H.; Hong, Y.; Yang, Q.; Chi, X.Y.; Yang, Z.; Yu, S.L.; Li, B.L.; et al. Transcriptome identification of the resistance-associated genes (RAGs) to Aspergillus flavus infection in pre-harvested peanut (Arachis hypogaea). Funct. Plant Biol. 2013, 40, 292–303. [Google Scholar] [CrossRef]
- Rokas, A.; Payne, G.; Fedorova, N.D.; Baker, S.E.; Machida, M.; Yu, J.; Georgianna, D.R.; Dean, R.A.; Bhatnagar, D.; Cleveland, T.E.; et al. What can comparative genomics tell us about species concepts in the genus Aspergillus? Stud. Mycol. 2007, 59, 11–17. [Google Scholar] [CrossRef]
- Georgianna, D.R.; Fedorova, N.D.; Burroughs, J.L.; Dolezal, A.L.; Bok, J.W.; Horowitz-Brown, S.I.; Woloshuk, C.P.; Yu, J.; Keller, N.P.; Payne, G.A. Beyond aflatoxin: Four distinct expression patterns and functional roles associated with Aspergillus flavus secondary metabolism gene clusters. Mol. Plant Pathol. 2010, 11, 213–226. [Google Scholar] [CrossRef]
- Marui, J.; Yamane, N.; Ohashi-Kunihiro, S.; Ando, T.; Terabayashi, Y.; Sano, M.; Ohashi, S.; Ohshima, E.; Tachibana, K.; Higa, Y.; et al. Kojic acid biosynthesis in Aspergillus oryzae is regulated by a Zn(II)(2)Cys(6) transcriptional activator and induced by kojic acid at the transcriptional level. J. Biosci. Bioeng. 2011, 112, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chang, P.K.; Bhatnagar, D.; Cleveland, T.E. Cloning of a sugar utilization gene cluster in Aspergillus parasiticus. Biochem. Biophys. Acta 2000, 1493, 211–214. [Google Scholar] [CrossRef]
- Amare, M.G.; Keller, N.P. Molecular mechanisms of Aspergillus flavus secondary metabolism and development. Fungal Genet. Biol. 2014, 66, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, J.E.; Payne, G.A. Overexpression of aflR leads to upregulation of pathway gene transcription and increased aflatoxin production in Aspergillus flavus. Appl. Environ. Microbiol. 1997, 63, 3995–4000. [Google Scholar] [PubMed]
- Meyers, D.M.; Obrian, G.; Du, W.L.; Bhatnagar, D.; Payne, G.A. Characterization of aflJ, a gene required for conversion of pathway intermediates to aflatoxin. Appl. Environ. Microbiol. 1998, 64, 3713–3717. [Google Scholar]
- Chang, P.K. The Aspergillus parasiticus protein AFLJ interacts with the aflatoxin pathway-specific regulator AFLR. Mol. Genet. Genom. 2003, 268, 711–719. [Google Scholar]
- Chang, P.K.; Yu, J.; Yu, J.H. aflT a MFS transporter-encoding gene located in the aflatoxin gene cluster, does not have a significant role in aflatoxin secretion. Fungal Genet. Biol. 2004, 41, 911–920. [Google Scholar] [CrossRef]
- Ehrlich, K.C.; Mack, B.M.; Wei, Q.; Li, P.; Roze, L.V.; Dazzo, F.; Cary, J.; Bhatnagar, D.; Linz, J. Association with AflR in endosomes reveals new functions for AflJ in aflatoxin biosynthesis. Toxins 2012, 4, 1582–1600. [Google Scholar] [CrossRef]
- Yang, K.; Liang, L.; Ran, F.; Liu, Y.; Li, Z.; Lan, H.; Gao, P.; Zhuang, Z.; Zhang, F.; Nie, X.; et al. The DmtA methyltransferase contributes to Aspergillus flavus conidiation, sclerotial production, aflatoxin biosynthesis and virulence. Sci. Rep. 2016, 6, 23259. [Google Scholar] [CrossRef]
- Wan, L.; Li, B.; Lei, Y.; Yan, L.; Huai, D.; Kang, Y.; Jiang, H.; Tan, J.; Liao, B. Transcriptomic profiling reveals pigment regulation during peanut testa development. Plant Physiol. Biochem. 2018, 125, 116–125. [Google Scholar] [CrossRef]
- Solano, R.; Gimenez-Ibanez, S. Nuclear jasmonate and salicylate signaling and crosstalk in defense against pathogens. Front. Plant Sci. 2013, 4, 72. [Google Scholar] [Green Version]
- Blankenship, P.; Cole, R.; Sanders, T.; Hill, R. Effect of geo carposphere temperature on pre-harvest colonization of drought-stressed peanuts by Aspergillus flavus and subsequent aflatoxin contamination. Mycopathologia 1984, 85, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhou, G.; Hong, Y.; Chen, X.; Liu, H.; Li, S. Overview of research progress on peanut (Arachis hypogaea L.) host resistance to aflatoxin contamination and genomics at the Guangdong Academy of Agricultural Sciences. Peanut Sci. 2009, 36, 29–34. [Google Scholar] [CrossRef]
- Assis, R.; Barrionuevo, D.L.; Giorda, L.M.; Nores, M.E.; Aldao, M.A. Aflatoxin production in six peanut (Arachis hypogaea L.) genotypes infected with Aspergillus flavus and Aspergillus parasiticus, isolated from peanut production areas of Cordoba, Argentina. J. Agric. Food Chem. 2005, 53, 9274–9280. [Google Scholar] [CrossRef] [PubMed]
- Waliyar, F.; Kumar, P.L.; Traoré, A.; Ntare, B.R.; Diarra, B.; Kodio, O. Pre and post-harvest management of aflatoxin contamination in peanuts. In Mycotoxins: Detection Methods, Management, Public Health and Agricultural Trade; Leslie, J.F., Bandyopadhyay, R., Visconti, A., Eds.; CABI: Wallingford, UK, 2008; pp. 209–218. [Google Scholar]
- Hell, K.; Mutegi, C. Aflatoxin control and prevention strategies in key crops of Sub-Saharan Africa. Afr. J. Microbiol. Res. 2011, 55, 459–466. [Google Scholar]
- Nigam, S.N.; Waliyar, F.; Aruna, R.; Reddy, S.V.; Kumar, P.L.; Craufurd, P.Q.; Diallo, A.T.; Ntare, B.R.; Upadhyaya, H.D. Breeding for resistance to aflatoxin contamination at ICRISAT. Peanut Sci. 2009, 36, 42–49. [Google Scholar] [CrossRef]
- Njoroge, S.M.C. A critical review of aflatoxin contamination of peanuts in Malawi and Zambia: The past, present, and future. Plant Dis. 2018, 102, 2394–2406. [Google Scholar] [CrossRef]
- Mixon, A.C.; Bell, D.K.; Wilson, D.M. Effect of chemical and biological agents on the incidence of A. flavus and aflatoxin contamination of peanut seed. Phytopatho 1984, 74, 1440–1444. [Google Scholar] [CrossRef]
- Chalwe, H.; Mweetwa, A.M.; Lungu, O.I.; Phiri, E.; Njoroge, S.; Brandenburg, R. Reducing pre-harvest aflatoxin content in groundnuts through soil water. RUFORUM Work. Doc. Ser. 2016, 14, 921–926. Available online: http://repository.ruforum.org (accessed on 30 March 2019).
- Dorner, J.W. Development of biocontrol technology to manage aflatoxin contamination in peanuts. Peanut Sci. 2009, 36, 60–67. [Google Scholar] [CrossRef]
- Bandyopadhyay, R.; Cotty, P.J. Biological controls for aflatoxin reduction. In Aflatoxins: Finding Solutions for Improved Food Safety; Grace, D., Unnevehr, L., Eds.; International Food Policy Research Institute: Washington, DC, USA, 2013. [Google Scholar]
- Dick, K.M. Pest Management in Stored Groundnuts. Information Bulletin 22; ICRISAT: Patancheru, India, 1987; p. 28. [Google Scholar]
- Devi, R.; Hall, A. Strategies for reducing aflatoxin levels in groundnut based foods and feeds in India: A step towards improving health of humans and livestocks; Section C: Scientific Background; ICRISAT: Patancheru, India, 2000; pp. 9–17. [Google Scholar]
- Attah, I.A.; Awuah, R.T.; Kpodo, K.A.; Fialor, S.C.; Jolly, C.M. Cost effectiveness of selected post-harvest pod handling techniques against damage, mouldiness and aflatoxin contamination of shelled groundnut in Ghana. J. Food Sci. Technol. 2007, 27, 17–26. [Google Scholar]
- Bulaong, S.S.P.; Dharmaputra, O.S. Fungal population, aflatoxin and free fatty acid contents of peanuts packed in different bag types. Biotropica 2002, 19, 1–25. [Google Scholar]
- Sudini, H.; Ranga Rao, G.V.; Gowda, C.L.L.; Chandrika, R.; Margam, V.; Rathore, A.; Murdock, L.L. Purdue Improved Crop Storage (PICS) bags for safe storage of groundnuts. J. Stored Prod. Res. 2015, 64, 133–138. [Google Scholar] [CrossRef]



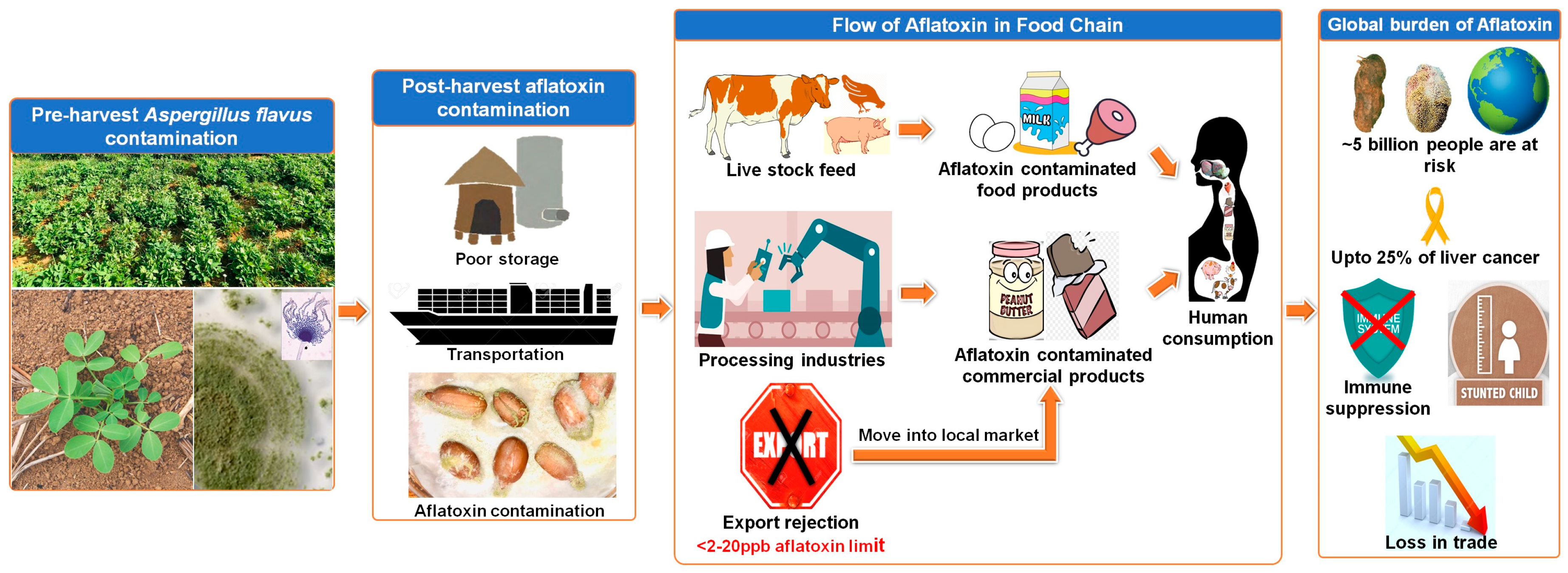{kind=link}
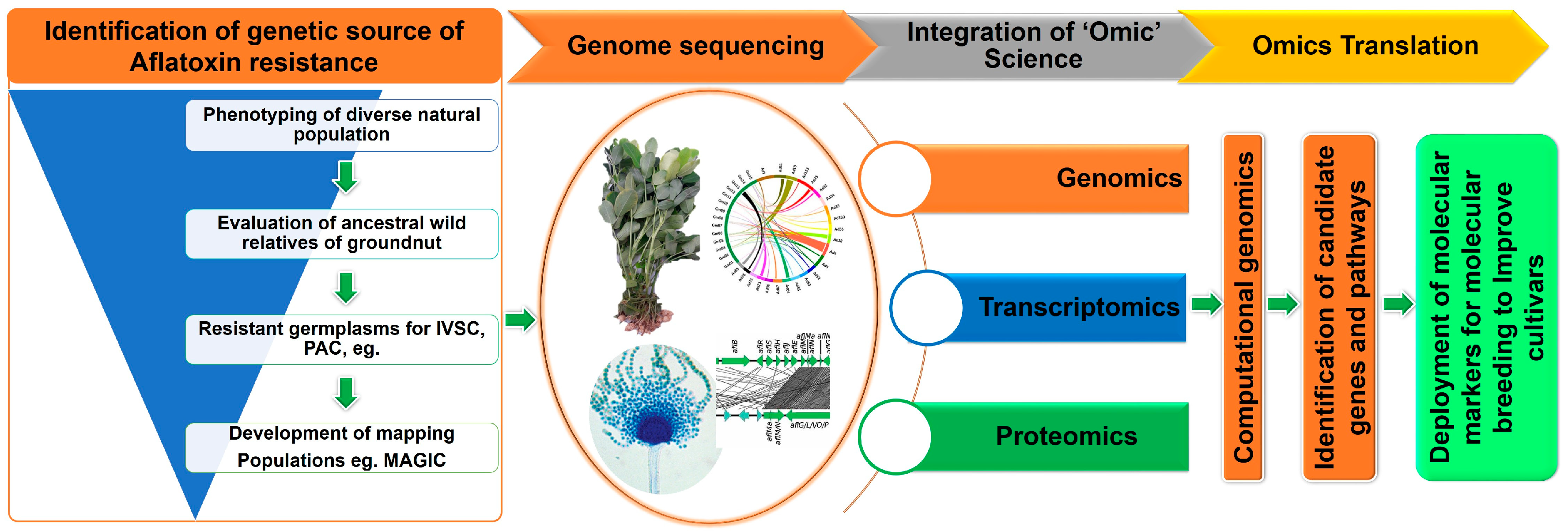{kind=link}
| Resistance Mechanism | Sample Size and Material Type | Toxigenic Species | Screening Method | Significant Outcome | Resistant Varieties Identified | Reference |
|---|---|---|---|---|---|---|
| 1. PHAC | 13 elite varieties and landraces | A. flavus | Mycelial growth on the surface of kernels | The compact arrangement of palisade-like layers of the seed testa is resistant to PHAC | Igola, Serenut 1, Serenut 2 and entry 99527 | [14] |
| 10 elite varieties | A. flavus | Seed infection coverage and intensity analyzer (SICIA) | All groundnut genotypes support PHAC, but AP varies among genotypes | ICG 1471, NC3033, ICGV 88145, GT-C20 | [15] | |
| 2. PAC | 7 elite varieties | A. flavus | Green conidial heads of A. flavus group and black conidial heads of A. niger | Low levels of linoleic acid do not affect aflatoxin production during PAC | F1334 and F1344 | [89] |
| 11 germplasm lines | A. flavus | Aflatoxin estimated using ELISA | PAC increases when exposed to terminal drought | ICGV 98305, ICGV 98348, ICGV 98353, Tifton 8 | [47] | |
| 3. IVSC | 40 elite varieties | A. flavus | Seeds with A. flavus colonies were counted | IVSC increases with increased drought stress | 55–437, PI 337409, PI 337394F, 73–30 | [90] |
| 35 wild accessions | A. flavus | IVSC and AP | A. pusilla, A. chiquitana, A. triseminata species resistant to IVSC and AP | ICG 13212, ICG 11560, ICG 8131, ICG 14875 | [13] | |
| 37 cultivars | A. flavus | A. flavus inoculated | Different varieties produce aflatoxin B1 and B2 at different levels during IVSC | PI 337394F, PI 337409, J-11 | [91] | |
| >100 accessions, breeding lines and commercial varieties | A. flavus | Visual development of conidial spores | Lower moisture has higher level of resistance to penetration by A. flavus during IVSC | PI 337394, PI 337409 | [92] | |
| 14 varieties | A. parasiticus | Fungal sporulation recorded | Higher moisture reduces infection rate during IVSC | J-11, Lampang | [93] | |
| 12 breeding and germplasm lines | A. flavus, A. parasiticus | Immunoaffinity column fluorometer method | Highly significant (E), (G) and (G × E) interactions identified | AR-2, GFA-1 | [94] | |
| IVSC and AP | 25 breeding lines and cultivars of Africa | A. flavus and A. parasiticus | ELISA for toxin estimation | VAR 27 variety produced least aflatoxin but showed higher IVSC | ICGV 87084, ICGV 87094, ICGV 87110 | [12] |
| 67 CSL lines and varieties | A. flavus | Seed colonization test | Varieties with compact and thicker testa resistant to IVSC | 12CS-104, 73-33 | [16] | |
| 850 cultivars and elite lines | A. flavus | Seed colonies and aflatoxin estimated | Some varieties are susceptible to IVSC but, resistant to AP | PI 337394F, PI337409 and UF71513 | [95] | |
| 561 germplasm lines | A. flavus | Seed infection percentage and aflatoxin production recorded | ICRISAT core collection has more resistance to IVSC than the China core collection | ICG 12625 (resistant to AP) and ICG 4750 (resistant to seed invasion) | [96] |
| Resistance Mechanism | Key Genes/TFs and Pathways Identified | Functional Description | References |
|---|---|---|---|
| Aflatoxin Production (AP) | WRKY | Stress regulative transcription factor | [32] |
| Toll/Interleukin1 receptor-nucleotide binding site leucine-rich repeat (TIR-NBS-LRR) | Highly conserved disease resistant genes in plants | ||
| Ethylene responsive factors | Transcriptionally regulates jasmonate signaling pathway | ||
| Heat shock proteins | Regulates heat shock factors which play vital role in plant defense | ||
| Pathogenesis-related (PR) 1,2,5 | Defense-related genes | [30] | |
| NBS-LRR genes | Disease resistance gene | ||
| WRKY | Stress regulative transcription factor | ICRISAT, Unpublished | |
| Ethylene responsive factors | Plays intermediary role in salicylic acid pathway | ||
| Linoleate 9S-lipoxygenase | Plays role in Jasmonic acid signal transduction pathway | ||
| Pre-Harvest Aflatoxin Contamination (PAC) | ABR1 | Ethylene responsive transcription factor and repressor of ABA signaling | [108] |
| Pathogenesis related-2 | Stress and defense responsive gene | ||
| BIG | Auxin transport gene | ||
| WRINKLED1 | Controls fatty acid biosynthesis pathway | ||
| Defensin | Defense response | [109] | |
| TIR | Defense response | ||
| Chalcone isomerase 3 | Flavonoids biosynthesis | ||
| EM protein | Stress response | ||
| Cupin/Oxalate oxidase | Seed storage protein | [97] | |
| Fatty acid desaturase 1 | Regulates fatty acid-biosynthesis pathway | ||
| Lipoxygenase | Plays role in Jasmonic acid signal transduction pathway | ||
| Iso-Ara h3 | Seed Storage protein | [98] | |
| LEA 4 | Stress related protein | ||
| Cu/Zn superoxide dismutase II | Antioxidant defensive protein | ||
| Heat shock protein | Regulates heat shock factors which play vital role in plant defense | ||
| In Vitro Seed Colonization (IVSC) | Linoleate 9S-lipoxygenase | Plays role in Jasmonic acid signaling transduction | [31] |
| Resveratrol synthase | Biosynthesize stilbene type-phytoalexins | ||
| Chalcone synthase | Flavonoids biosynthesis | ||
| Defensins | Defense response | ||
| Chitinases | Modulates immune response | ||
| Post-Harvest Aflatoxin Contamination (PHAC) | Heat shock protein 70 | Maintains internal cell stability like folding-unfolding of proteins | [106] |
| Heat shock protein 90 | Cellular immunity, signal transduction | ||
| NB-LRR | PAMPs perception | [99] | |
| Hypersensitive induced response protein | Hypersensitive response | ||
| S-locus glycoprotein | Induction of defense | ||
| Cytochrome P450 | Degradation of toxins | ||
| Alcohol dehydrogenase-1F | Detoxification | ||
| SAM dependent isoflavone 7-O-methyltransferase | Biosynthesis of phytoalexins | ||
| Seed linoleate | Lipid metabolism | ICRISAT, Unpublished | |
| Resveratrol synthase | Biosynthesis stilbene type-phytoalexins | ||
| ABA responsive genes | Regulates stress responsive genes |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandey, M.K.; Kumar, R.; Pandey, A.K.; Soni, P.; Gangurde, S.S.; Sudini, H.K.; Fountain, J.C.; Liao, B.; Desmae, H.; Okori, P.; et al. Mitigating Aflatoxin Contamination in Groundnut through A Combination of Genetic Resistance and Post-Harvest Management Practices. Toxins 2019, 11, 315. https://doi.org/10.3390/toxins11060315
Pandey MK, Kumar R, Pandey AK, Soni P, Gangurde SS, Sudini HK, Fountain JC, Liao B, Desmae H, Okori P, et al. Mitigating Aflatoxin Contamination in Groundnut through A Combination of Genetic Resistance and Post-Harvest Management Practices. Toxins. 2019; 11(6):315. https://doi.org/10.3390/toxins11060315
Chicago/Turabian StylePandey, Manish K., Rakesh Kumar, Arun K. Pandey, Pooja Soni, Sunil S. Gangurde, Hari K. Sudini, Jake C. Fountain, Boshou Liao, Haile Desmae, Patrick Okori, and et al. 2019. "Mitigating Aflatoxin Contamination in Groundnut through A Combination of Genetic Resistance and Post-Harvest Management Practices" Toxins 11, no. 6: 315. https://doi.org/10.3390/toxins11060315
APA StylePandey, M. K., Kumar, R., Pandey, A. K., Soni, P., Gangurde, S. S., Sudini, H. K., Fountain, J. C., Liao, B., Desmae, H., Okori, P., Chen, X., Jiang, H., Mendu, V., Falalou, H., Njoroge, S., Mwololo, J., Guo, B., Zhuang, W., Wang, X., ... Varshney, R. K. (2019). Mitigating Aflatoxin Contamination in Groundnut through A Combination of Genetic Resistance and Post-Harvest Management Practices. Toxins, 11(6), 315. https://doi.org/10.3390/toxins11060315