Application of Zearalenone (ZEN)-Detoxifying Bacillus in Animal Feed Decontamination through Fermentation

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Screening of Bacillus Strains for ZEN Detoxification Potential
2.2. ZEN Detoxification Capability, Adsorption Ability, and Degradation Ability of ZDB Strains
2.3. Enzymatic Profile of ZDB Strains
2.4. Detoxification of ZEN-Contaminated Maize by ZDB Strains
2.5. The Effect of ZEN on B2 Strain Growth in TSB Medium
2.6. The Effect of ZEN on Fermentation Characteristics of B2 Strain in Maize
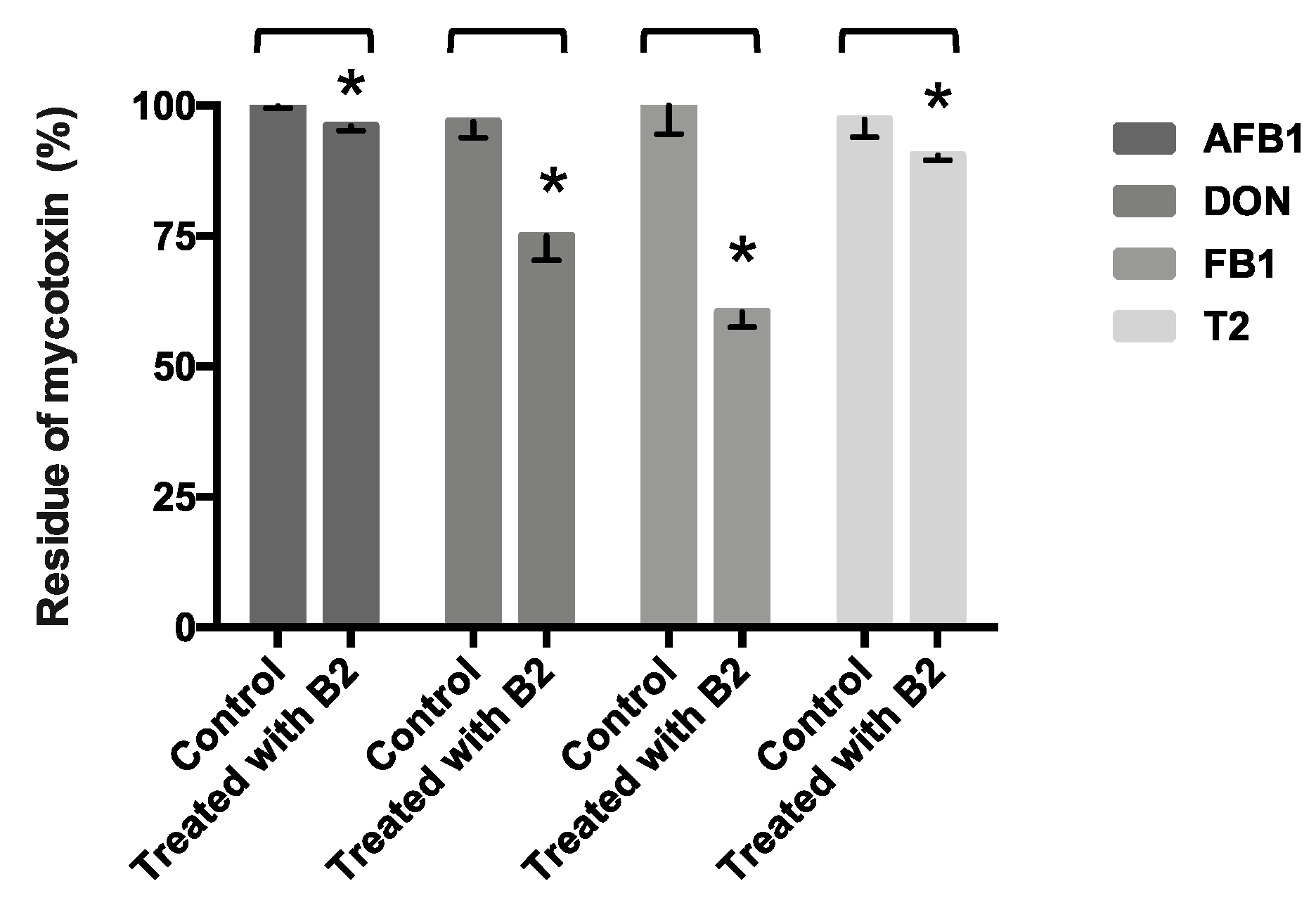
2.7. Other Mycotoxins’ (AFB1, DON, FB1, and T2 Toxin) Detoxification
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Isolation of Bacillus Strains
5.3. Screening of ZEN Detoxification Potential Strains
5.4. Bacterial Strain Identification
5.5. Bacillus-Related Enterotoxin Detection
5.6. ZEN Detoxification Capability Test in Culturing Condition (TSB Medium)
5.7. ZEN Adsorption Ability and ZEN Degradation Ability
5.8. Enzymatic Profile of ZDB Strains
5.9. Detoxification of ZEN-Contaminated Maize by ZDB Strains
5.10. The Effect of ZEN on the Candidate Strain (B2 Strain) Growth in TSB Medium
5.11. The Effect of ZEN on Fermentation Characteristics of the Candidate Strain (B2) in Maize
5.12. Other Mycotoxin (AFB1, DON, FB1 and T2 Toxin) Detoxification Test
5.13. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Chaytor, A.C.; Hansen, J.A.; van Heugten, E.; See, M.T.; Kim, S.W. Occurrence and decontamination of mycotoxins in swine feed. Asian-Australas. J. Anim. Sci. 2011, 24, 723–738. [Google Scholar] [CrossRef]
- Schatzmayr, G.; Streit, E. Global occurrence of mycotoxins in the food and feed chain: Facts and figures. World Mycotoxin J. 2013, 6, 213–222. [Google Scholar] [CrossRef]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Piastowska-Ciesielska, A.W. Zearalenone as an endocrine disruptor in humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Coulombe, R.A., Jr. Biological action of mycotoxins. J. Dairy Sci. 1993, 6, 880–891. [Google Scholar] [CrossRef]
- Schatzmayr, G.; Zehner, F.; Täubel, M.; Schatzmayr, D.; Klimitsch, A.; Loibner, A.P.; Binder, E.M. Microbiologicals for deactivating mycotoxins. Mol. Nutr. Food Res. 2006, 50, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Vekiru, E.; Hametner, C.; Mitterbauer, R.; Rechthaler, J.; Adam, G.; Schatzmayr, G.; Krska, R.; Schuhmacher, R. Cleavage of zearalenone by Trichosporon mycotoxinivorans to a novel nonestrogenic metabolite. Appl. Environ. Microbiol. 2010, 76, 2353–2359. [Google Scholar] [CrossRef] [PubMed]
- Huwig, A.; Freimund, S.; Käppeli, O.; Dutler, H. Mycotoxin detoxication of animal feed by different adsorbents. Toxicol. Lett. 2001, 122, 179–188. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Park, M.A.; Ha, J.K. Mycotoxins and their biotransformation in the rumen: A review. Asian-Australas. J. Anim. Sci. 2010, 23, 1250–1260. [Google Scholar] [CrossRef]
- EFSA. Introduction of a Qualified Presumption of Safety (QPS) approach for assessment of selected microorganisms referred to EFSA. EFSA J. 2007, 587, 1–16. [Google Scholar] [CrossRef]
- Missotten, J.A.M.; Michiels, J.; Degroote, J.; De Smet, S. Fermented liquid feed for pigs: An ancient technique for the future. J. Anim. Sci. Biotechnol. 2015, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Canibe, N.; Jensen, B.B. Fermented liquid feed-microbial and nutritional aspects and impact on enteric diseases in pigs. Anim. Feed Sci. Technol. 2012, 173, 17–40. [Google Scholar] [CrossRef]
- Reddy, K.R.N.; Salleh, B.; Saad, B.; Abbas, H.K.; Abel, C.A.; Shier, W.T. An overview of mycotoxin contamination in foods and its implications for human health. Toxin Rev. 2010, 29, 3–26. [Google Scholar] [CrossRef]
- El-Sharkawy, S.; Abul-Hajj, Y.J. Microbial cleavage of zearalenone. Xenobiotica 1988, 18, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Kakeya, H.; Takahashi-Ando, N.; Kimura, M.; Onose, R.; Yamaguchi, I.; Osada, H. Biotransformation of the mycotoxin, zearalenone, to a non-estrogenic compound by a fungal strain of Clonostachys sp. Biosci. Biotechnol. Biochem. 2002, 66, 2723–2726. [Google Scholar] [CrossRef] [PubMed]
- Molnar, O.; Schatzmayr, G.; Fuchs, E.; Prillinger, H. Trichosporon mycotoxinivorans sp. nov., a new yeast species useful in biological detoxification of various mycotoxins. Syst. Appl. Microbiol. 2004, 27, 661–671. [Google Scholar] [CrossRef]
- Varga, J.; Péteri, Z.; Tábori, K.; Téren, J.; Vágvölgyi, C. Degradation of ochratoxin A and other mycotoxins by Rhizopus isolates. Int. J. Food Microbiol. 2005, 99, 321–328. [Google Scholar] [CrossRef]
- Altalhi, A.D.; El-Deeb, B. Localization of zearalenone detoxification gene(s) in pZEA-1 plasmid of seudomonas putida ZEA-1 and expressed in Escherichia coli. J. Hazard Mater. 2009, 161, 1166–1172. [Google Scholar] [CrossRef]
- Yu, Y.; Qiu, L.; Wu, H.; Tang, Y.; Yu, Y.; Li, X.; Liu, D. Degradation of zearalenone by the extracellular extracts of Acinetobacter sp. SM04 liquid cultures. Biodegradation 2011, 22, 613–622. [Google Scholar] [CrossRef]
- Cserháti, M.; Kriszt, B.; Krifaton, C.; Szoboszlay, S.; Háhn, J.; Tóth, S.; Nagy, I.; Kukolya, J. Mycotoxin-degradation profile of Rhodococcus strains. Int. J. Food Microbiol. 2013, 166, 176–185. [Google Scholar] [CrossRef]
- Kosawang, C.; Karlsson, M.; Vélëz, H.; Rasmussen, P.H.; Collinge, D.B.; Jensen, B.; Jensen, D.F. Zearalenone detoxification by zearalenone hydrolase is important for the antagonistic ability of Clonostachys rosea against mycotoxigenic Fusarium graminearum. Fungal Biol. 2014, 118, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; He, X.; Xue, K.; Li, Y.; Xu, D.; Qian, H. Biological detoxification of zearalenone by Aspergillus niger strain FS10. Food Chem. Toxicol. 2014, 72, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Hu, Y.; He, J.; Wu, L.; Liao, F.; Luo, B.; He, Y.; Zuo, Z.; Ren, Z.; Zhong, Z.; et al. Zearalenone degradation by two Pseudomonas strains from soil. Mycotoxin Res. 2014, 30, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Shetty, P.H.; Jespersen, L. Saccharomyces cerevisiae and lactic acid bacteria as potential mycotoxin decontaminating agents. Trends Food Sci. Technol. 2006, 17, 48–55. [Google Scholar] [CrossRef]
- Setlow, P. Spores of Bacillus subtilis: Their resistance to and killing by radiation, heat and chemicals. J. Appl. Microbiol. 2006, 101, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Yu, M.; Dong, F.; Shi, J.; Xu, J. Esterase activity inspired selection and characterization of zearalenone degrading bacteria Bacillus pumilus ES-21. Food Control 2017, 77, 57–64. [Google Scholar] [CrossRef]
- Tinyiro, S.E.; Wokadala, C.; Xu, D.; Yao, W. Adsorption and degradation of zearalenone by Bacillus strains. Folia Microbiol. 2011, 56, 321–327. [Google Scholar] [CrossRef]
- El-Nezami, H.; Polychronaki, N.; Lee, Y.K.; Haskard, C.; Juvonen, R.; Salminen, S.; Mykkänen, H. Chemical moieties and interactions involved in the binding of zearalenone to the surface of Lactobacillus rhamnosus strains GG. J. Agric. Food Chem. 2004, 52, 4577–4581. [Google Scholar] [CrossRef]
- Sangsila, A.; Faucet-Marquis, V.; Pfohl-Leszkowicz, A.; Itsaranuwat, P. Detoxification of zearalenone by Lactobacillus pentosus strains. Food Control 2016, 62, 187–192. [Google Scholar] [CrossRef]
- Dalié, D.K.D.; Deschamps, A.M.; Richard-Forget, F. Lactic acid bacteria-potential for control of mould growth and mycotoxins: A review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Olsen, M.; Malmlöf, K.; Pettersson, H.; Sandholm, K.; Kiessling, K.H. Plasma and urinary levels of zearalenone and alpha-zearalenol in a prepubertal gilt fed zearalenone. Acta Pharmacol. Toxicol. 1985, 56, 239–243. [Google Scholar] [CrossRef]
- Van Winsen, R.L.; Lipman, L.J.A.; Biesterveld, S.; Urlings, B.A.P.; Snijders, J.M.A.; Van Knapen, F. Mechanism of Salmonella reduction in fermented pig feed. J. Sci. Food Agric. 2001, 81, 342–346. [Google Scholar] [CrossRef]
- Missotten, J.A.M.; Goris, J.; Michiels, J.; Van Coillie, E.; Herman, L.; De Smet, S.; Dierick, N.A.; Heyndrickx, M. Screening of isolated lactic acid bacteria as potential beneficial strains for fermented liquid pig feed production. Anim. Feed Sci. Tech. 2009, 150, 122–138. [Google Scholar] [CrossRef]
- Sonenshein, A.L. Control of key metabolic intersections in Bacillus subtilis. Nat. Rev. Microbiol. 2007, 5, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Shivers, R.P.; Dineen, S.S.; Sonenshein, A.L. Positive regulation of Bacillus subtilis ackA by CodY and CcpA: Establishing a potential hierarchy in carbon flow. Mol. Microbiol. 2006, 62, 811–822. [Google Scholar] [CrossRef]
- Grenier, B.; Oswald, I. Mycotoxin co-contamination of food and feed: Meta-analysis of publications describing toxicological interactions. World Mycotoxin J. 2011, 4, 285–313. [Google Scholar] [CrossRef]
- Wang, L.T.; Lee, F.L.; Tai, C.J.; Kasai, H. Comparison of gyrB gene sequences, 16S rRNA gene sequences and DNA-DNA hybridization in the Bacillus subtilis group. Int. J. Syst. Evol. Microbiol. 2007, 57, 1846–1850. [Google Scholar] [CrossRef]
- Ok, H.E.; Choi, S.W.; Kim, M.; Chun, H.S. HPLC and UPLC methods for the determination of zearalenone in noodles, cereal snacks and infant formula. Food Chem. 2014, 163, 252–257. [Google Scholar] [CrossRef]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar]


{kind=link}
| Strains | Source | Enterotoxins b | Emetic Toxin | Code c | 16S rDNA Sequencing (Identify%) | ||
|---|---|---|---|---|---|---|---|
| Nhe A | Nhe B | Hbl L2 | Cereulide | ||||
| Isolate 1 | Fermented soybean product | − | − | − | − | B1 | B. subtilis (99) |
| Isolate 2 | Fermented soybean product | + | − | − | − | ||
| Isolate 3 | Fermented soybean product | − | − | − | − | B2 | B. subtilis (99) |
| Isolate 4 | Fermented soybean product | + | − | − | − | ||
| Isolate 5 | Fermented soybean product | − | − | − | − | B3 | B. subtilis (99) |
| Isolate 6 | Fermented soybean product | − | − | − | − | B4 | Bacillus sp. (100) |
| Isolate 7 | Fermented soybean product | + | − | − | − | ||
| Isolate 8 | Sewage | + | − | − | − | ||
| Isolate 9 | Soil | + | − | − | − | ||
| Isolate 10 | Soil | + | − | − | − | ||
| Isolate 11 | Soil | + | − | − | − | ||
| Isolate 12 | Soil | + | + | − | − | ||
| Isolate 13 | Soil | − | − | − | − | B5 | Bacillus sp. (100) |
| Isolate 14 | Soil | + | − | − | − | ||
| Bacillus Strains | Detoxification Rate (%) * | Absorption Rate (%) * | Degradation Rate (%) * |
|---|---|---|---|
| BCRC 17,441 # | 51.9 ± 7.55 a | 47.0 ± 9.26 a | 31.0 ± 0.45 |
| B1 | 41.8 ± 6.35 b | 29.0 ± 3.43 b | 24.0 ± 8.09 |
| B2 | 41.4 ± 2.76 b | 26.5 ± 6.11 b | 35.0 ± 4.22 |
| B3 | 28.1 ± 3.50 c | 29.6 ± 3.79 b | 27.2 ± 0.24 |
| B4 | 58.1 ± 3.02 a | 24.7 ± 9.31 b | 31.0 ± 4.15 |
| B5 | 43.1 ± 0.94 b | 30.4 ± 2.86 b | 32.8 ± 5.57 |
| Enzyme | BCRC 17,441 | B1 | B2 | B3 | B4 | B5 |
|---|---|---|---|---|---|---|
| Alkaline phosphatase | + a | + | + | ++ | + | ++ |
| Acid phosphatase | + | ++ | ++ | + | + | + |
| Esterase (C4) | + | ++ | ++ | + | + | + |
| Esterase lipase (C8) | + | + | + | ++ | + | + |
| Lipase (C14) | − | − | − | − | − | − |
| Leucine arylamidase | + | − | − | + | + | ++ |
| Valine arylamidase | + | − | − | + | + | + |
| Cystine arylamidase | − | − | − | + | − | + |
| Trypsin | − | − | − | − | − | − |
| α-chymotrypsin | + | − | − | + | + | + |
| Naphthol-AS-BI-phosphohydrolase | + | + | + | + | + | + |
| α-galactosidase | − | − | − | + | − | − |
| ß-galactosidase | + | + | − | + | + | + |
| ß-glucuronidase | − | − | − | − | − | − |
| α-glucosidase | + | − | − | ++ | + | + |
| ß-glucosidase | − | − | − | + | − | + |
| N-acetyl-ß-glucosaminidase | − | − | − | − | − | − |
| α-mannosidase | − | − | − | − | − | − |
| α-fucosidase | + | − | − | − | − | − |
| Bacillus Strains | Bacterial Count (log CFU mL−1) | Detoxification of ZEN in Maize after 48 h (%) | |
|---|---|---|---|
| 24 h | 48 h | ||
| BCRC 17,441 | 9.67 ± 0.561 a | 8.69 ± 0.499 a | 32.7 ± 10.42 b,c |
| B1 | 6.32 ± 0.922 b | 7.09 ± 0.379 b | 49.0 ± 9.43 a,b |
| B2 | 9.44 ± 0.175 a | 9.20 ± 0.153 a | 55.8 ± 6.20 a |
| B3 | 9.11 ± 0.494 a | 8.81 ± 0.438 a | 31.4 ± 10.96 c |
| B4 | 8.21 ± 1.861 a | 8.79 ± 0.358 a | 49.2 ± 8.46 a,b |
| B5 | 9.32 ± 0.210 a | 9.19 ± 0.516 a | 38.3 ± 3.57 b,c |
| Measurement | −ZEN | +ZEN |
|---|---|---|
| Bacterial count (log CFU mL−1) (8 h) | 10.4 ± 0.60 | 11.8 ± 0.58 * |
| Bacterial count (log CFU mL−1) (24 h) | 10.3 ± 0.05 | 11.3 ± 0.14 * |
| pH | 4.77 ± 0.042 | 5.64 ± 0.056 * |
| NH3-N (mmol L−1) | 11.33 ± 0.891 | 27.57 ± 1.819 * |
| Lactic acid (mmol L−1) | 208.2 ± 8.02 | 284.8 ± 20.02 * |
| Acetic acid (mmol L−1) | 5.22 ± 0.269 | 11.55 ± 1.030 * |
| Total VFAs (mml L−1) | 5.88 ± 0.325 | 12.41 ± 1.042 * |
| Measurement | −ZEN | +ZEN |
|---|---|---|
| Bacterial count (log CFU·mL−1) (48 h) | 8.65 ± 0.334 | 8.99 ± 0.289 |
| Bacterial count (log CFU·mL−1) (72 h) | 8.84 ± 0.186 | 8.50 ± 0.327 |
| pH (48 h) | 4.58 ± 0.022 * | 4.46 ± 0.070 |
| pH (72 h) | 4.35 ± 0.068 | 4.41 ± 0.079 |
| NH3-N (mmol·L−1) (48 h) | 6.03 ± 0.372 | 6.10 ± 0.259 |
| NH3-N (mmol·L−1) (72 h) | 5.97 ± 0.230 | 6.65 ± 0.269 * |
| Lactic acid (mmol·L−1) (48 h) | 67.8 ± 13.51 | 96.8 ± 8.88 * |
| Lactic acid (mmol·L−1) (72 h) | 88.8 ± 12.26 | 111.9 ± 15.33 |
| Acetic acid (mmol·L−1) (48 h) | 21.3 ± 0.18 | 19.0 ± 1.02 |
| Acetic acid (mmol·L−1) (72 h) | 21.6 ± 0.82 * | 19.4 ± 0.62 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-W.; Wang, H.-T.; Shih, W.-Y.; Ciou, Y.-A.; Chang, Y.-Y.; Ananda, L.; Wang, S.-Y.; Hsu, J.-T. Application of Zearalenone (ZEN)-Detoxifying Bacillus in Animal Feed Decontamination through Fermentation. Toxins 2019, 11, 330. https://doi.org/10.3390/toxins11060330
Chen S-W, Wang H-T, Shih W-Y, Ciou Y-A, Chang Y-Y, Ananda L, Wang S-Y, Hsu J-T. Application of Zearalenone (ZEN)-Detoxifying Bacillus in Animal Feed Decontamination through Fermentation. Toxins. 2019; 11(6):330. https://doi.org/10.3390/toxins11060330
Chicago/Turabian StyleChen, Shiau-Wei, Han-Tsung Wang, Wei-Yuan Shih, Yan-An Ciou, Yu-Yi Chang, Laurensia Ananda, Shu-Yin Wang, and Jih-Tay Hsu. 2019. "Application of Zearalenone (ZEN)-Detoxifying Bacillus in Animal Feed Decontamination through Fermentation" Toxins 11, no. 6: 330. https://doi.org/10.3390/toxins11060330
APA StyleChen, S. -W., Wang, H. -T., Shih, W. -Y., Ciou, Y. -A., Chang, Y. -Y., Ananda, L., Wang, S. -Y., & Hsu, J. -T. (2019). Application of Zearalenone (ZEN)-Detoxifying Bacillus in Animal Feed Decontamination through Fermentation. Toxins, 11(6), 330. https://doi.org/10.3390/toxins11060330