Venomous Landmines: Clinical Implications of Extreme Coagulotoxic Diversification and Differential Neutralization by Antivenom of Venoms within the Viperid Snake Genus Bitis


 ,
, 
Abstract
:1. Introduction
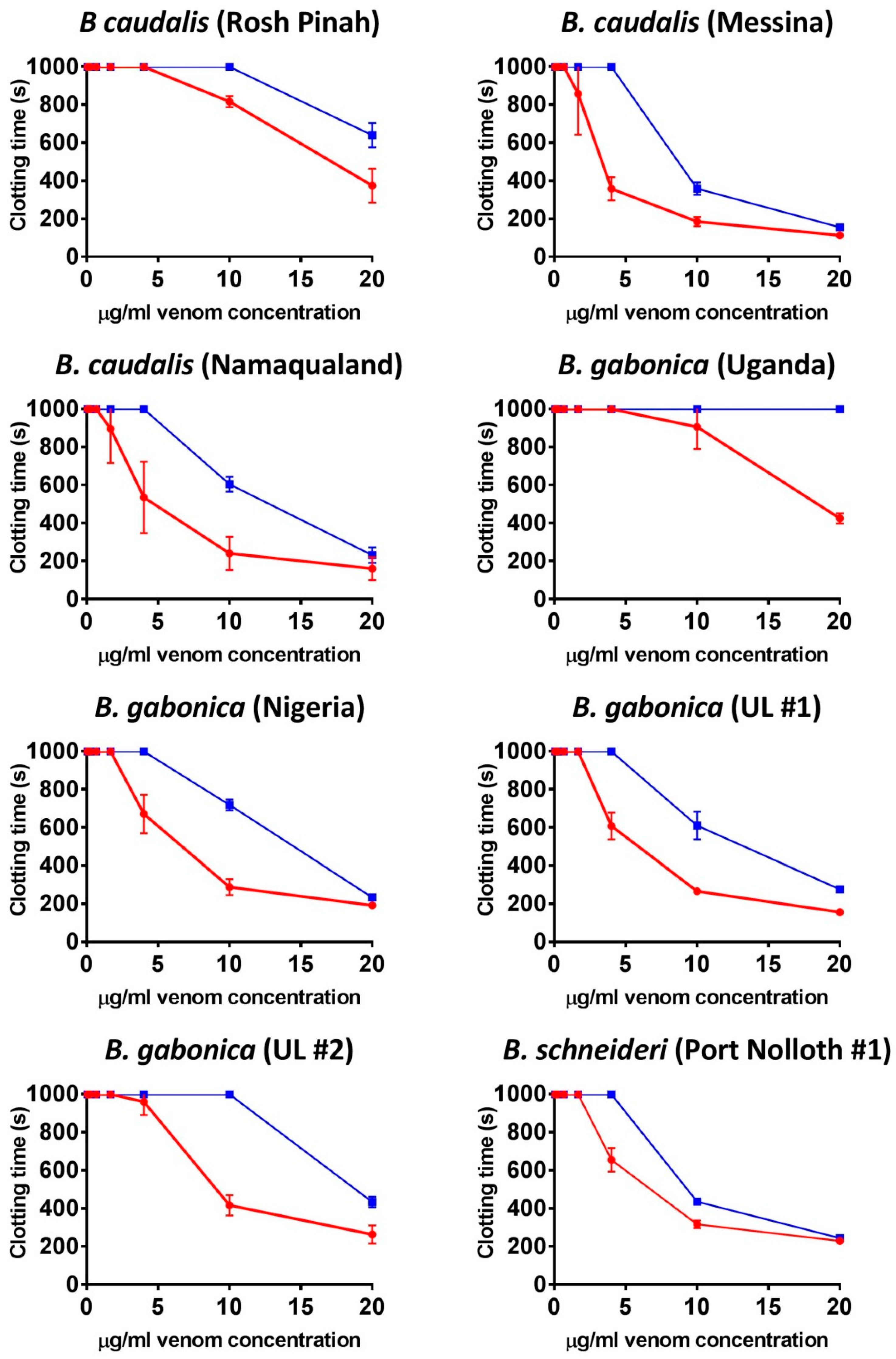
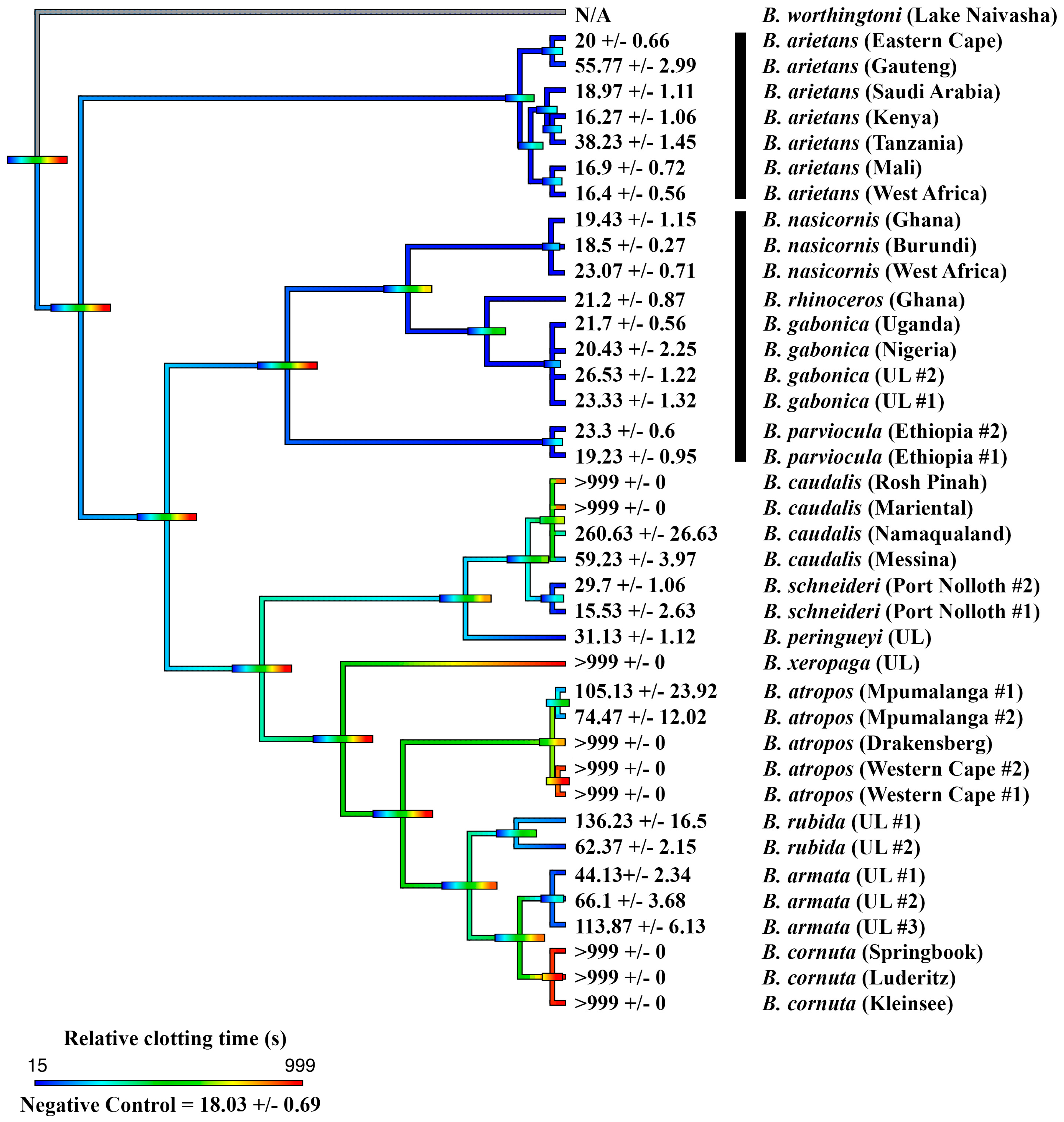
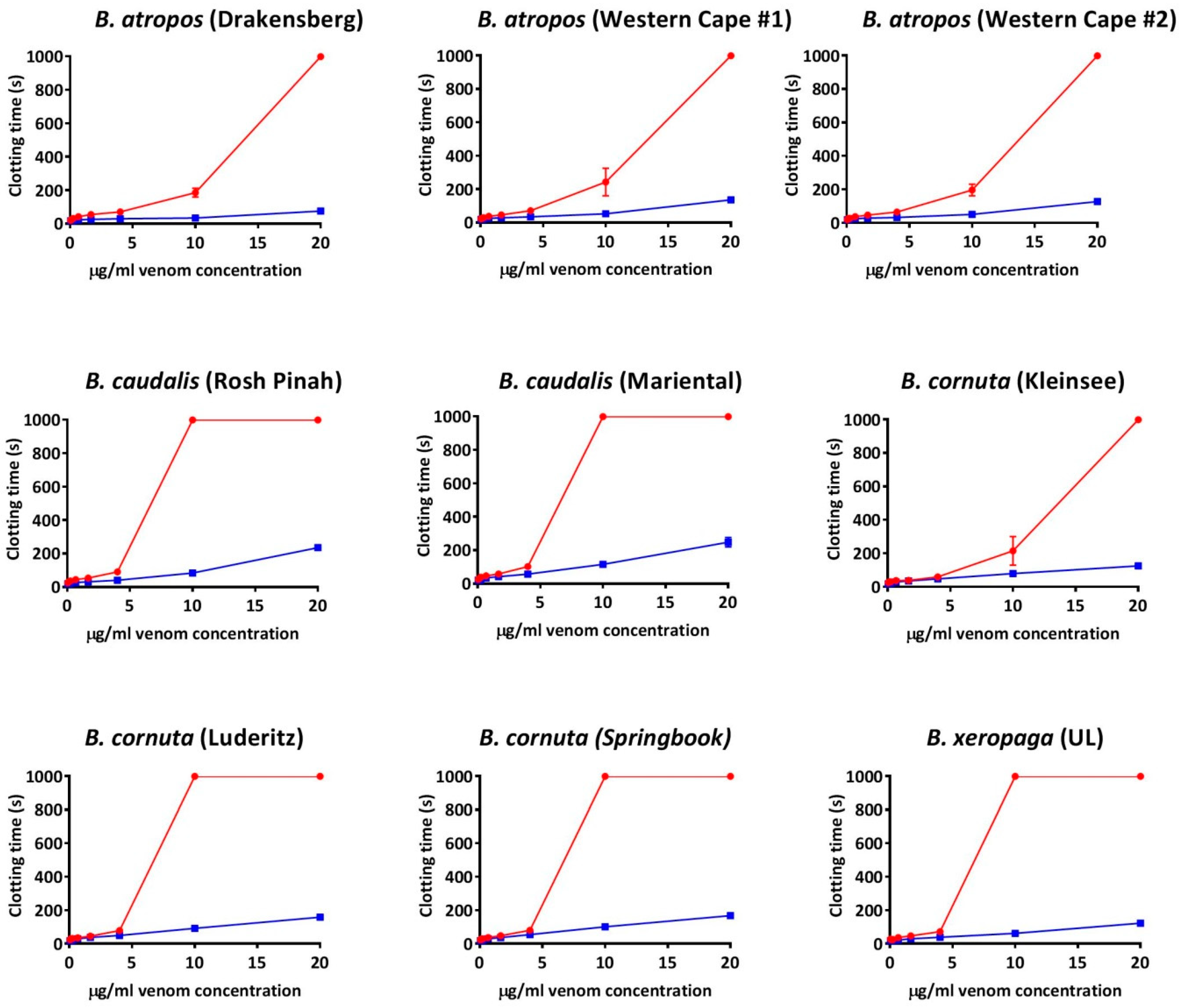
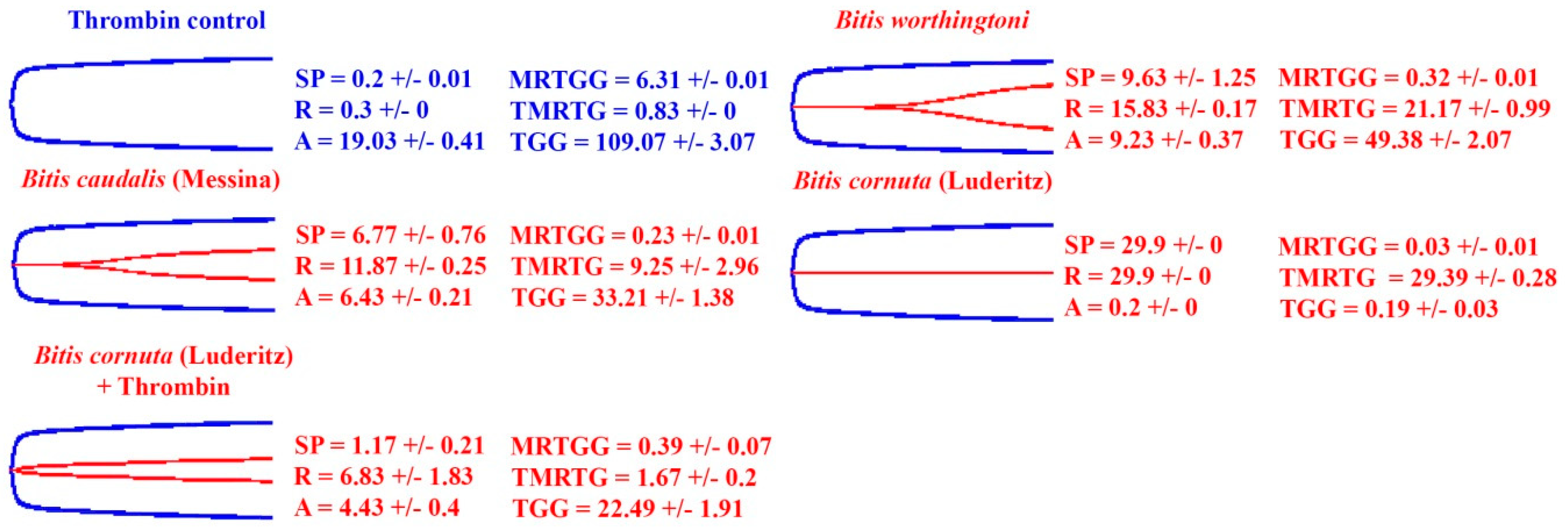
2. Results
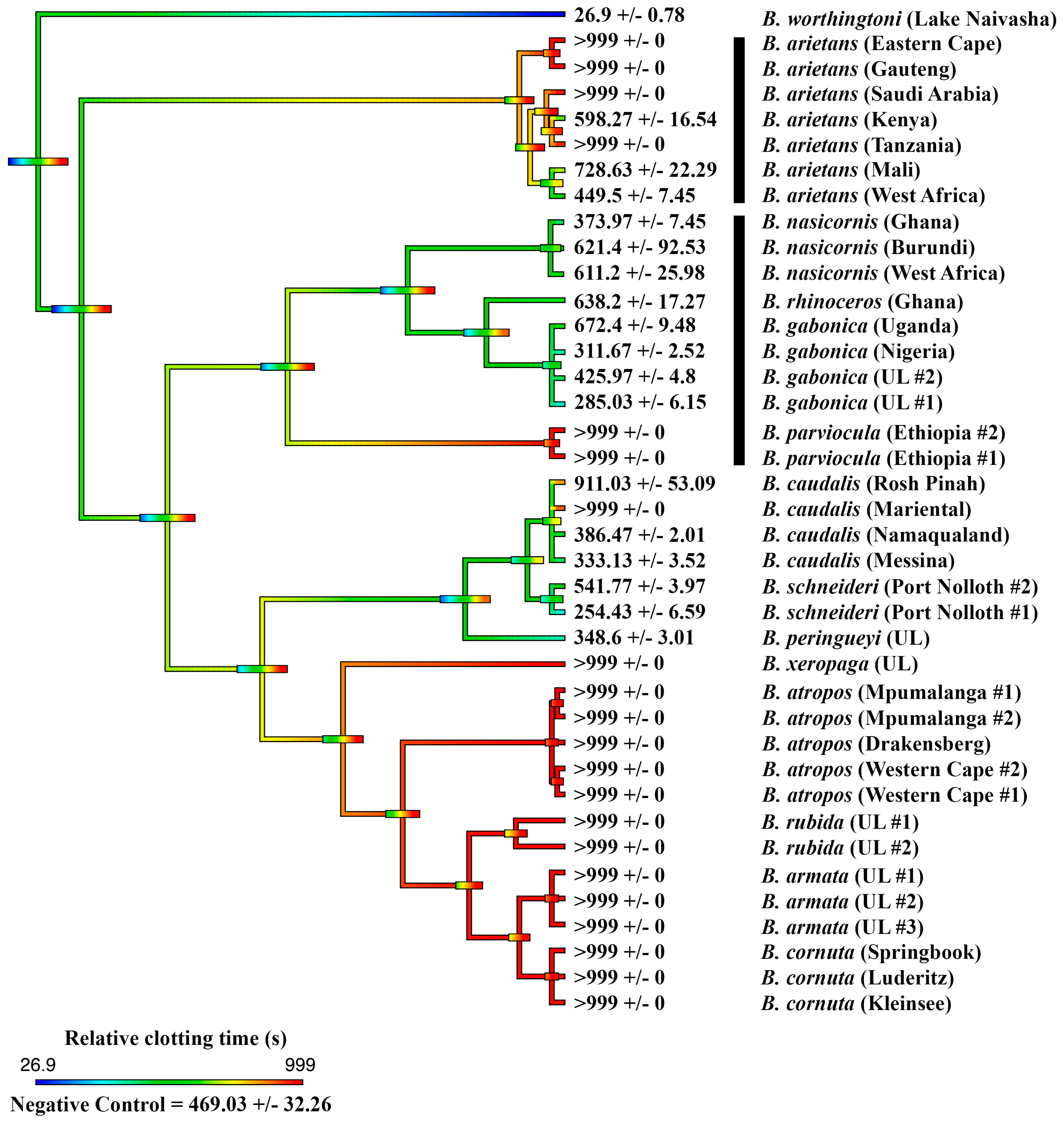
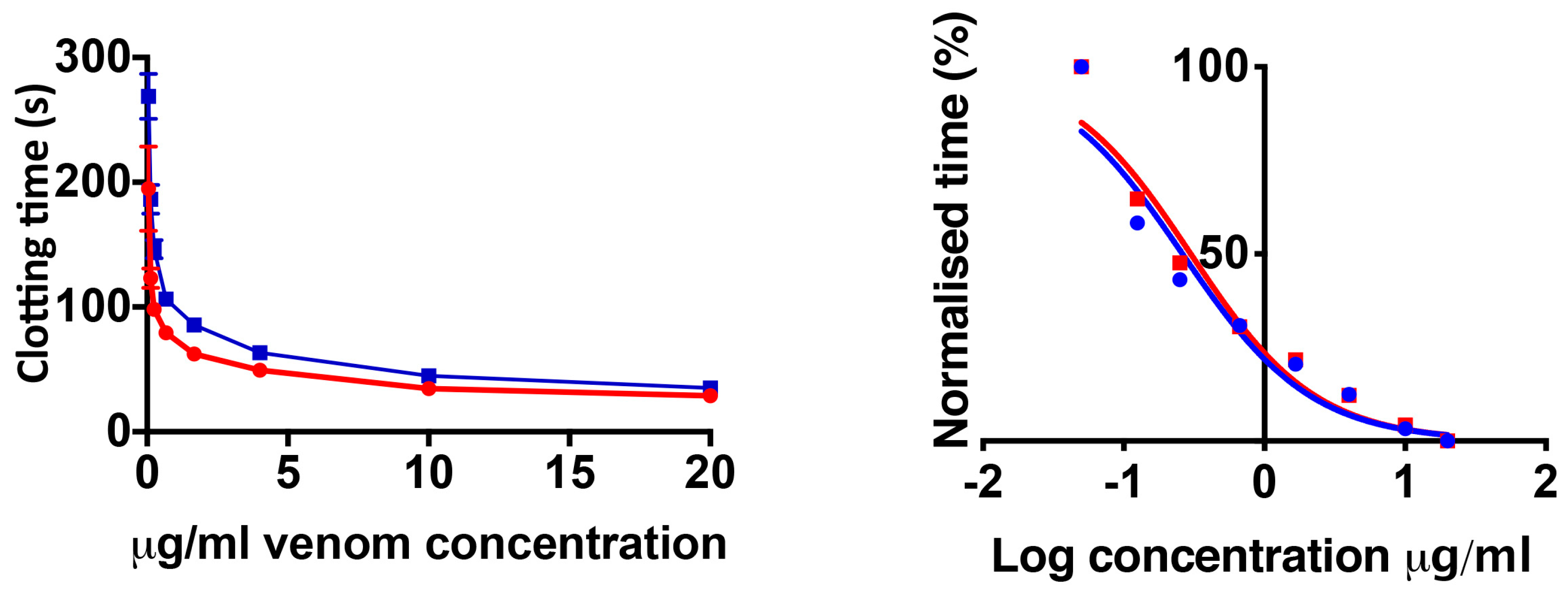
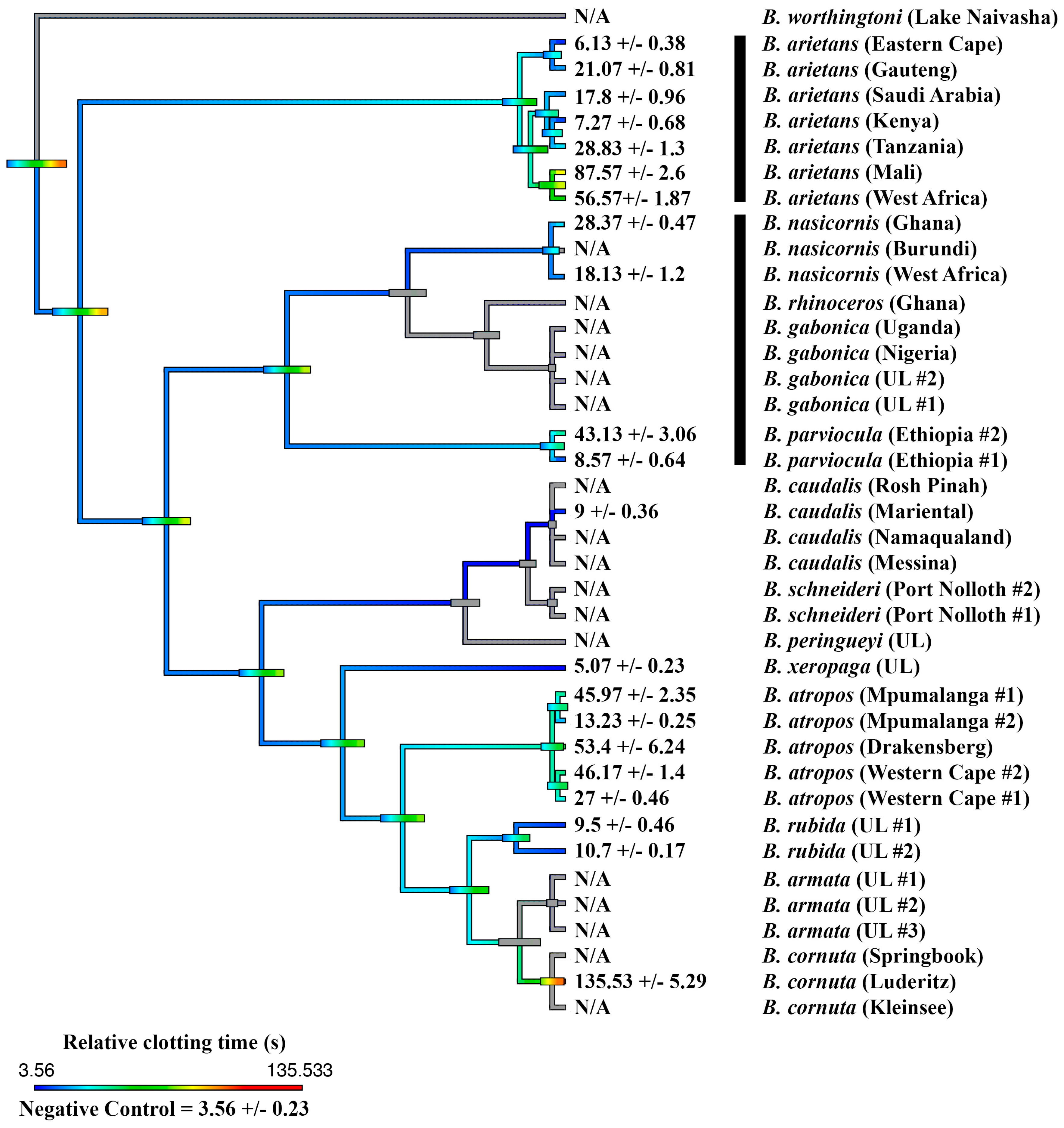
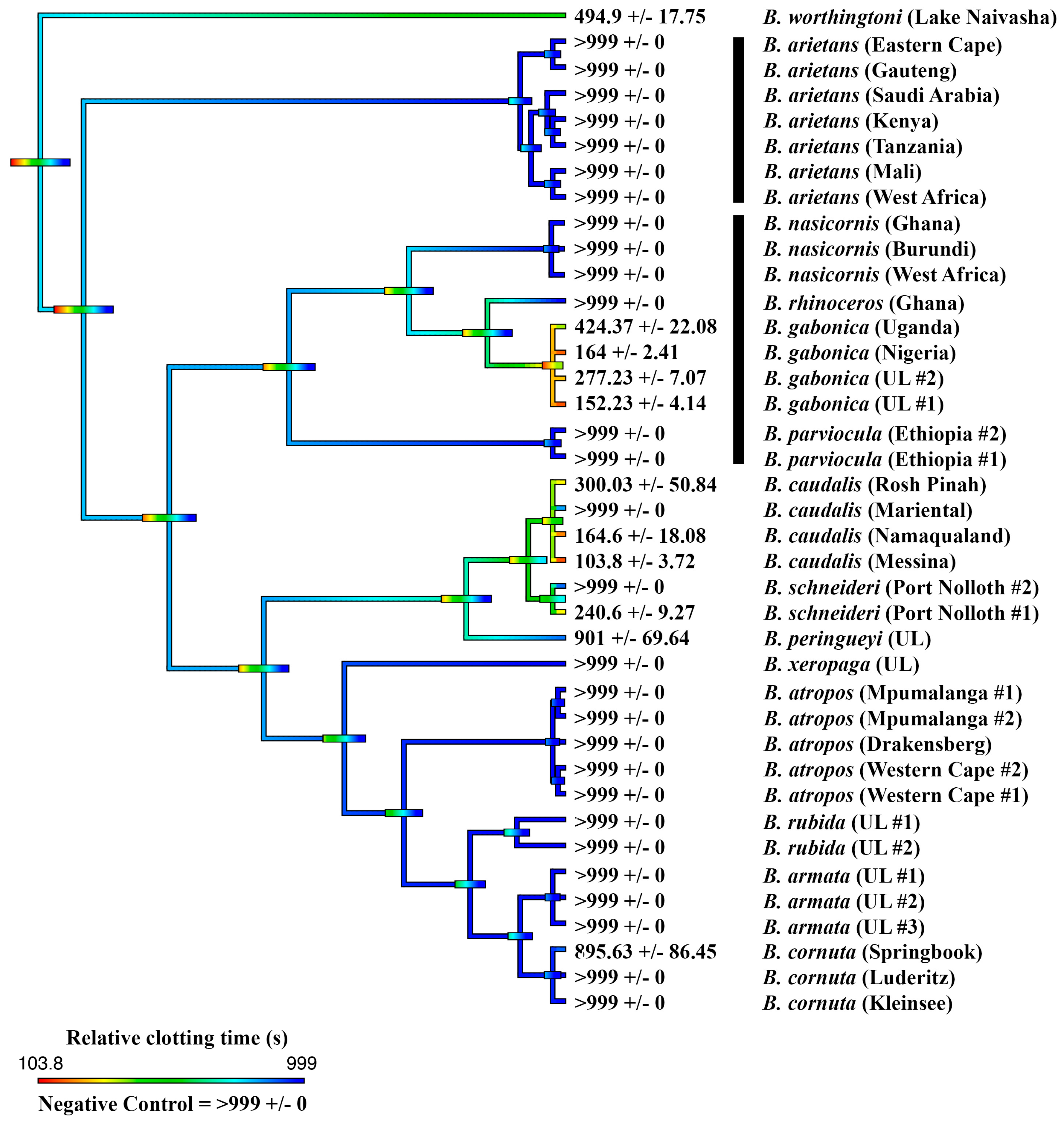
2.1. Coagulation Assays
2.2. Thromboelastography
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Venoms and Reagents
5.2. Coagulation Assays
5.3. Thromboelastography (TEG)
5.4. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fry, B. Snakebite: When the Human Touch Becomes a Bad Touch. Toxins 2018, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The Global Burden of Snakebite: A Literature Analysis and Modelling Based on Regional Estimates of Envenoming and Deaths (Global Burden of Snakebite). PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [PubMed]
- Broadley, D.G. The Venomous Snakes of Central and South Africa. Venomous Animals and Their Venoms; New York Academic Press: New York, NY, USA, 1960; Volume 1. [Google Scholar]
- Mallow, D.; Ludwig, D.; Nilson, G. True Vipers: Natural History and Toxinology of Old World Vipers; Krieger Publishing Company: Malabar, FL, USA, 2003; p. 359. [Google Scholar]
- Phelps, T. Old World Vipers: A Natural History of the Azemiopinae and Viperinae; Chimaira Buchhandelsgesellschaft mbH: Frankfurt, Germany, 2010; p. 558. [Google Scholar]
- Alencar, L.R.V.; Quental, T.B.; Grazziotin, F.G.; Alfaro, M.L.; Martins, M.; Venzon, M.; Zaher, H. Diversification in vipers: Phylogenetic relationships, time of divergence and shifts in speciation rates. Mol. Phylogenet. Evol. 2016, 105, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Lenk, P.; Herrmann, H.W.; Joger, U.; Wink, M. Phylogeny and taxonomic subdivisions of Bitis (Reptilia: Viperidae) based on molecular evidence. Kaupia 1999, 8, 31–38. [Google Scholar]
- Lenk, P.; Kalyabina, S.; Wink, M.; Joger, U. Evolutionary Relationships among the True Vipers (Reptilia: Viperidae) Inferred from Mitochondrial DNA Sequences. Mol. Phylogenet. Evol. 2001, 19, 94–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barlow, A.; Wüster, W.; Kelly, C.M.R.; Branch, W.R.; Phelps, T.; Tolley, K.A. Ancient habitat shifts and organismal diversification are decoupled in the African viper genus Bitis (Serpentes: Viperidae. J. Biogeogr. 2019, 46, 1234–1248. [Google Scholar] [CrossRef]
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2012, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Holding, M.L.; Margres, M.J.; Rokyta, D.R.; Gibbs, H.L. Local prey community composition and genetic distance predict venom divergence among populations of the northern Pacific rattlesnake (Crotalus oreganus). J. Evol. Biol. 2018, 31, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Deuis, J.; Dashevsky, D.; Dobson, J.; Jackson, T.; Brust, A.; Xie, B.; Koludarov, I.; Debono, J.; Hendrikx, I.; et al. The Snake with the Scorpion’s Sting: Novel Three-Finger Toxin Sodium Channel Activators from the Venom of the Long-Glanded Blue Coral Snake (Calliophis bivirgatus). Toxins 2016, 8, 303. [Google Scholar] [CrossRef]
- Cipriani, V.; Debono, J.; Goldenberg, J.; Jackson, T.N.W.; Arbuckle, K.; Dobson, J.; Koludarov, I.; Li, B.; Hay, C.; Dunstan, N.; et al. Correlation between ontogenetic dietary shifts and venom variation in Australian brown snakes (Pseudonaja). Comp. Biochem. Physiol. Part C 2017, 197, 53–60. [Google Scholar] [CrossRef]
- Pawlak, J.; Mackessy, S.P.; Sixberry, N.M.; Stura, E.A.; Le Du, M.H.; Ménez, R.; Foo, C.S.; Ménez, A.; Nirthanan, S.; Kini, R.M. Irditoxin, a novel covalently linked heterodimeric three-finger toxin with high taxon-specific neurotoxicity. FASEB J. Off. Publ. Fed. Am. Soc. Exper. Biol. 2009, 23, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Dobson, J.; Cochran, C.; Dashevsky, D.; Arbuckle, K.; Benard, M.; Boyer, L.; Alagón, A.; Hendrikx, I.; Hodgson, W.; et al. The Bold and the Beautiful: A Neurotoxicity Comparison of New World Coral Snakes in the Micruroides and Micrurus Genera and Relative Neutralization by Antivenom. Neurodegener. Neuroregener. Neurotroph. Action Neuroprot. 2017, 32, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Wüster, W.; Kini, R.M.; Brusic, V.; Khan, A.; Venkataraman, D.; Rooney, A.P. Molecular Evolution and Phylogeny of Elapid Snake Venom Three-Finger Toxins. J. Mol. Evol. 2003, 57, 110–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatesh, M.; Prasad, N.; Sing, T.; Gowda, V. Purification, Characterization, and Chemical Modification of Neurotoxic Peptide from Daboia russelii Snake Venom of India. J. Biochem. Mol. Toxicol. 2013, 27, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Jackson, T.; Koludarov, I.; Ali, S.; Dobson, J.; Zdenek, C.; Dashevsky, D.; Brouw, B.; Masci, P.; Nouwens, A.; Josh, P.; et al. Rapid Radiations and the Race to Redundancy: An Investigation of the Evolution of Australian Elapid Snake Venoms. Toxins 2016, 8, 309. [Google Scholar] [CrossRef] [PubMed]
- Zdenek, C.N.; Hay, C.; Arbuckle, K.; Jackson, T.N.W.; Bos, M.H.A.; Op Den Brouw, B.; Debono, J.; Allen, L.; Dunstan, N.; Morley, T.; et al. Coagulotoxic effects by brown snake (Pseudonaja) and taipan (Oxyuranus) venoms, and the efficacy of a new antivenom. Toxicol. In Vitro 2019, 58, 97–109. [Google Scholar] [CrossRef]
- Jin, N.Z.; Gopinath, S.C.B. Potential blood clotting factors and anticoagulants. Biomed. Pharmacother. 2016, 84, 356–365. [Google Scholar] [CrossRef]
- Kini, R.; Koh, C. Metalloproteases Affecting Blood Coagulation, Fibrinolysis and Platelet Aggregation from Snake Venoms: Definition and Nomenclature of Interaction Sites. Toxins 2016, 8, 284. [Google Scholar] [CrossRef]
- Coimbra, F.C.P.; Dobson, J.; Zdenek, C.N.; Op Den Brouw, B.; Hamilton, B.; Debono, J.; Masci, P.; Frank, N.; Ge, L.; Kwok, H.F.; et al. Does size matter? Venom proteomic and functional comparison between night adder species (Viperidae: Causus) with short and long venom glands. Comp. Biochem. Physiol. Part C 2018, 211, 7–14. [Google Scholar] [CrossRef]
- Debono, J.; Bos, M.H.A.; Coimbra, F.; Ge, L.; Frank, N.; Kwok, H.F.; Fry, B.G. Basal but divergent: Clinical implications of differential coagulotoxicity in a clade of Asian vipers. Toxicol. In Vitro 2019, 58, 195–206. [Google Scholar] [CrossRef]
- Debono, J.; Bos, M.H.A.; Nouwens, A.; Ge, L.; Frank, N.; Kwok, H.F.; Fry, B.G. Habu coagulotoxicity: Clinical implications of the functional diversification of Protobothrops snake venoms upon blood clotting factors. Toxicol. In Vitro 2019, 55, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Youngman, N.J.; Zdenek, C.N.; Dobson, J.S.; Bittenbinder, M.A.; Gillett, A.; Hamilton, B.; Dunstan, N.; Allen, L.; Veary, A.; Veary, E.; et al. Mud in the blood: Novel potent anticoagulant coagulotoxicity in the venoms of the Australian elapid snake genus Denisonia (mud adders) and relative antivenom efficacy. Toxicol. Lett. 2019, 302, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lister, C.; Arbuckle, K.; Jackson, T.N.W.; Debono, J.; Zdenek, C.N.; Dashevsky, D.; Dunstan, N.; Allen, L.; Hay, C.; Bush, B.; et al. Catch a tiger snake by its tail: Differential toxicity, co-factor dependence and antivenom efficacy in a procoagulant clade of Australian venomous snakes. Comp. Biochem. Physiol. Part C 2017, 202, 39–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oulion, B.; Dobson, J.S.; Zdenek, C.N.; Arbuckle, K.; Lister, C.; Coimbra, F.C.P.; Op Den Brouw, B.; Debono, J.; Rogalski, A.; Violette, A.; et al. Factor X activating Atractaspis snake venoms and the relative coagulotoxicity neutralising efficacy of African antivenoms. Toxicol. Lett. 2018, 288, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Rogalski, A.; Soerensen, C.; Op Den Brouw, B.; Lister, C.; Dashevsky, D.; Arbuckle, K.; Gloria, A.; Zdenek, C.N.; Casewell, N.R.; Gutiérrez, J.M.; et al. Differential procoagulant effects of saw-scaled viper (Serpentes: Viperidae: Echis) snake venoms on human plasma and the narrow taxonomic ranges of antivenom efficacies. Toxicol. Lett. 2017, 280, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Sousa, L.F.; Zdenek, C.N.; Dobson, J.S.; Op Den Brouw, B.; Coimbra, F.; Gillett, A.; Del-Rei, T.H.M.; Chalkidis, H.d.M.; Sant’Anna, S.; Teixeira-Da-Rocha, M.M.; et al. Coagulotoxicity of Bothrops (Lancehead Pit-Vipers) Venoms from Brazil: Differential Biochemistry and Antivenom Efficacy Resulting from Prey-Driven Venom Variation. Toxins 2018, 10, 411. [Google Scholar] [CrossRef] [PubMed]
- Brink, S.; Steytler, J.G. Effects of puff-adder venom on coagulation, fibrinolysis and platelet aggregation in the baboon. S. Afr. Med. J. 1974, 48, 1205–1213. [Google Scholar] [PubMed]
- Fernandez, S.; Hodgson, W.; Chaisakul, J.; Kornhauser, R.; Konstantakopoulos, N.; Smith, A.I.; Kuruppu, S. In vitro toxic effects of puff adder (Bitis arietans) venom, and their neutralization by antivenom. Toxins 2014, 6, 1586–1597. [Google Scholar] [CrossRef] [PubMed]
- Morné, A.S.; Janette, B.; Sthembile, M.; Etheresia, P. The effect of physiological levels of South African puff adder (Bitis arietans) snake venom on blood cells: An in vitro model. Sci. Rep. 2016, 35988. [Google Scholar] [CrossRef]
- Visser, J.; Chapman, D.S. Snakes and Snakebite: Venomous Snakes and Management of Snakebite in Southern Africa; Purnell and Sons Ltd.: Cape Town, South Africa, 1978; p. 152. [Google Scholar]
- Marsh, N.A.; Whaler, B.C. Separation and Partial Characterization of a Coagulant Enzyme from Bitis gabonica Venom. Br. J. Haematol. 1974, 26, 295–306. [Google Scholar] [CrossRef]
- Branch, W.R. Field Guide to the Snakes and Other Reptiles of Southern Africa; Struik Publications: Cape Town, South Africa, 1992; p. 328. [Google Scholar]
- Sánchez, A.; Coto, J.; Segura, Á.; Vargas, M.; Solano, G.; Herrera, M.; Villalta, M.; Estrada, R.; Gutiérrez, J.M.; León, G. Effect of geographical variation of Echis ocellatus, Naja nigricollis and Bitis arietans venoms on their neutralization by homologous and heterologous antivenoms. Toxicon 2015, 108, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Viljoen, C.C.; Meehan, C.M.; Botes, D.P. Separation of Bitis gabonica (Gaboon adder) venom arginine esterases into kinin-releasing, clotting and fibrinolytic factors. Toxicon 1979, 17, 145–154. [Google Scholar] [CrossRef]
- Nielsen, V.; Cerruti, M.; Valencia, O.; Amos, Q. Decreased snake venom metalloproteinase effects via inhibition of enzyme and modification of fibrinogen. Int. J. Role Metal Ions Biol. Biochem. Med. 2016, 29, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Mackay, N.; Ferguson, J.C.; McNicol, G.P. Effects of the venom of the rhinoceros horned viper (Bitis nasicornis) on blood coagulation, platelet aggregation, and fibrinolysis. J. Clin. Pathol. 1970, 23, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.E.; Hotle, D.; Rodríguez-Acosta, A. Neutralization of Bitis parviocula (Ethiopian mountain adder) venom by the South African Institute of Medical Research (SAIMR) antivenom. Revista do Instituto de Medicina Tropical de Sao Paulo 2011, 53, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Ho, C.L.; Botes, D.P. Site of action of caudoxin, a neurotoxic phospholipase A 2 from the horned puff adder (Bitis caudalis) venom. Toxicon 1982, 20, 637–647. [Google Scholar] [CrossRef]
- Van Der Walt, A.J.; Muller, G.J. Berg adder (Bitis atropos) envenoming: An analysis of 14 cases. Clin. Toxicol. 2018, 1–6. [Google Scholar] [CrossRef]
- Van Zyl, J.M.; Müller, G.J. The berg adder (Bitis atropos) and its venom. Toxicon 1999, 37, 1335–1339. [Google Scholar]
- Van Zyl, J.M.; Müller, G.J.; Van der Merwe, M.J. Purification and properties of two phospholipase A 2 enzymes from berg adder (Bitis atropos) venom. S. Afr. J. Sci. 2001, 97, 437–444. [Google Scholar]
- Pourreau, F.; Pinsard, M.; Goyffon, M.; Plasse, F.; Desport, E.; Thierry, A.; Touchard, G.; Bridoux, F. Bilateral renal cortical necrosis with end-stage renal failure following envenoming by Proatheris superciliaris: A case report. Toxicon 2014, 84, 36–40. [Google Scholar] [CrossRef]
- Valenta, J.; Stach, Z.; Fricova, D.; Zak, J.; Balik, M. Envenoming by the viperid snake Proatheris superciliaris: A case report. Toxicon 2008, 52, 392–394. [Google Scholar] [CrossRef] [PubMed]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of Phylogenetics and Evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revell, L.J. Phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor Xa inhibition assay | Step 1: 50 µL 0.1 µg/mL venom (1 mg/mL 50% glycerol stock diluted with Owren–Koller (OK) Buffer (Stago catalog # 00360) + 50 µL 0.025 M calcium (Stago catalog # 00367) + 50 µL phospholipid (Stago catalog #00597) + 25 µL Factor Xa (Stago catalog #00311). Step 2: 120 s incubation. Step 3: Addition of 75 µL plasma. |
| Thrombin inhibition assay | Step 1: 50 µL 0.1 µg/mL venom (1 mg/mL 50% glycerol stock diluted with OK Buffer + 50 µL 0.025 M calcium + 50 µL phospholipid + 25 µL thrombin. Step 2: 120 s incubation. Step 3: Addition of 75 µL 4 mg/mL fibrinogen. |
| Fibrinogen destruction assay | Step 1: 50 µL 0.1 µg/mL venom (1 mg/mL 50% glycerol stock diluted with OK Buffer + 50 µL 0.025 M calcium + 50 µL phospholipid + 75 µL of 4 mg/mL fibrinogen. Step 2: 1-h incubation. Step 3: Addition of 25 µL thrombin (Stago Liquid Fib kit catalog # 00673). |
| Fibrinogen clotting assay | Step 1: 50 µL 0.1 µg/mL venom (1 mg/mL 50% glycerol stock diluted with OK Buffer + 50 µL 0.025 M calcium + 50 µL phospholipid + 25 µL OK Buffer. Step 2: 120 s incubation. Step 3: Addition of 75 µL of 4 mg/mL fibrinogen. |
| Fibrinogen clotting assay (antivenom assay conditions) | Step 1: 50 µL 0.1 µg/mL venom (1 mg/mL 50% glycerol stock diluted with OK Buffer + 50 µL 0.025 M calcium + 50 µL phospholipid + 25 µL of 5% concentration of SAIMR polyvalent antivenom diluted with OK Buffer. Step 2: 120 s incubation. Step 3: Addition of 75 µL 4 mg/mL fibrinogen. |
| Prothrombinase complex inhibition assay | Step 1: 50 µL 0.1 µg/mL venom (1 mg/mL 50% glycerol stock diluted with OK Buffer + 50 µL 0.025 M calcium + 50 µL phospholipid + 75 µL plasma. Step 2: 120 s incubation. Step 3: Addition of 25 µL Factor Xa (Stago catalog # 00311). |
| Prothrombinase complex inhibition assay (antivenom assay conditions) | Step 1: 25 µL 0.2 µg/mL venom (1 mg/mL 50% glycerol stock diluted with OK Buffer + 75 µL of [50 µL 0.025 M calcium +25 µL of 5% concentration of SAIMR polyvalent antivenom diluted in OK Buffer] + 50 µL phospholipid + 75 µL plasma. Step 2: 120 s incubation. Step 3: Addition of 25 µL Factor Xa. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youngman, N.J.; Debono, J.; Dobson, J.S.; Zdenek, C.N.; Harris, R.J.; Brouw, B.o.d.; Coimbra, F.C.P.; Naude, A.; Coster, K.; Sundman, E.; et al. Venomous Landmines: Clinical Implications of Extreme Coagulotoxic Diversification and Differential Neutralization by Antivenom of Venoms within the Viperid Snake Genus Bitis. Toxins 2019, 11, 422. https://doi.org/10.3390/toxins11070422
Youngman NJ, Debono J, Dobson JS, Zdenek CN, Harris RJ, Brouw Bod, Coimbra FCP, Naude A, Coster K, Sundman E, et al. Venomous Landmines: Clinical Implications of Extreme Coagulotoxic Diversification and Differential Neutralization by Antivenom of Venoms within the Viperid Snake Genus Bitis. Toxins. 2019; 11(7):422. https://doi.org/10.3390/toxins11070422
Chicago/Turabian StyleYoungman, Nicholas J., Jordan Debono, James S. Dobson, Christina N. Zdenek, Richard J. Harris, Bianca op den Brouw, Francisco C. P. Coimbra, Arno Naude, Kristian Coster, Eric Sundman, and et al. 2019. "Venomous Landmines: Clinical Implications of Extreme Coagulotoxic Diversification and Differential Neutralization by Antivenom of Venoms within the Viperid Snake Genus Bitis" Toxins 11, no. 7: 422. https://doi.org/10.3390/toxins11070422
APA StyleYoungman, N. J., Debono, J., Dobson, J. S., Zdenek, C. N., Harris, R. J., Brouw, B. o. d., Coimbra, F. C. P., Naude, A., Coster, K., Sundman, E., Braun, R., Hendrikx, I., & Fry, B. G. (2019). Venomous Landmines: Clinical Implications of Extreme Coagulotoxic Diversification and Differential Neutralization by Antivenom of Venoms within the Viperid Snake Genus Bitis. Toxins, 11(7), 422. https://doi.org/10.3390/toxins11070422