Biotransformation of the Mycotoxin Zearalenone to its Metabolites Hydrolyzed Zearalenone (HZEN) and Decarboxylated Hydrolyzed Zearalenone (DHZEN) Diminishes its Estrogenicity In Vitro and In Vivo


 , ,
, ,
Abstract
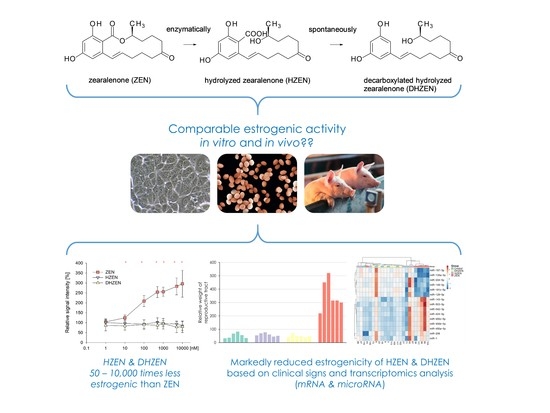
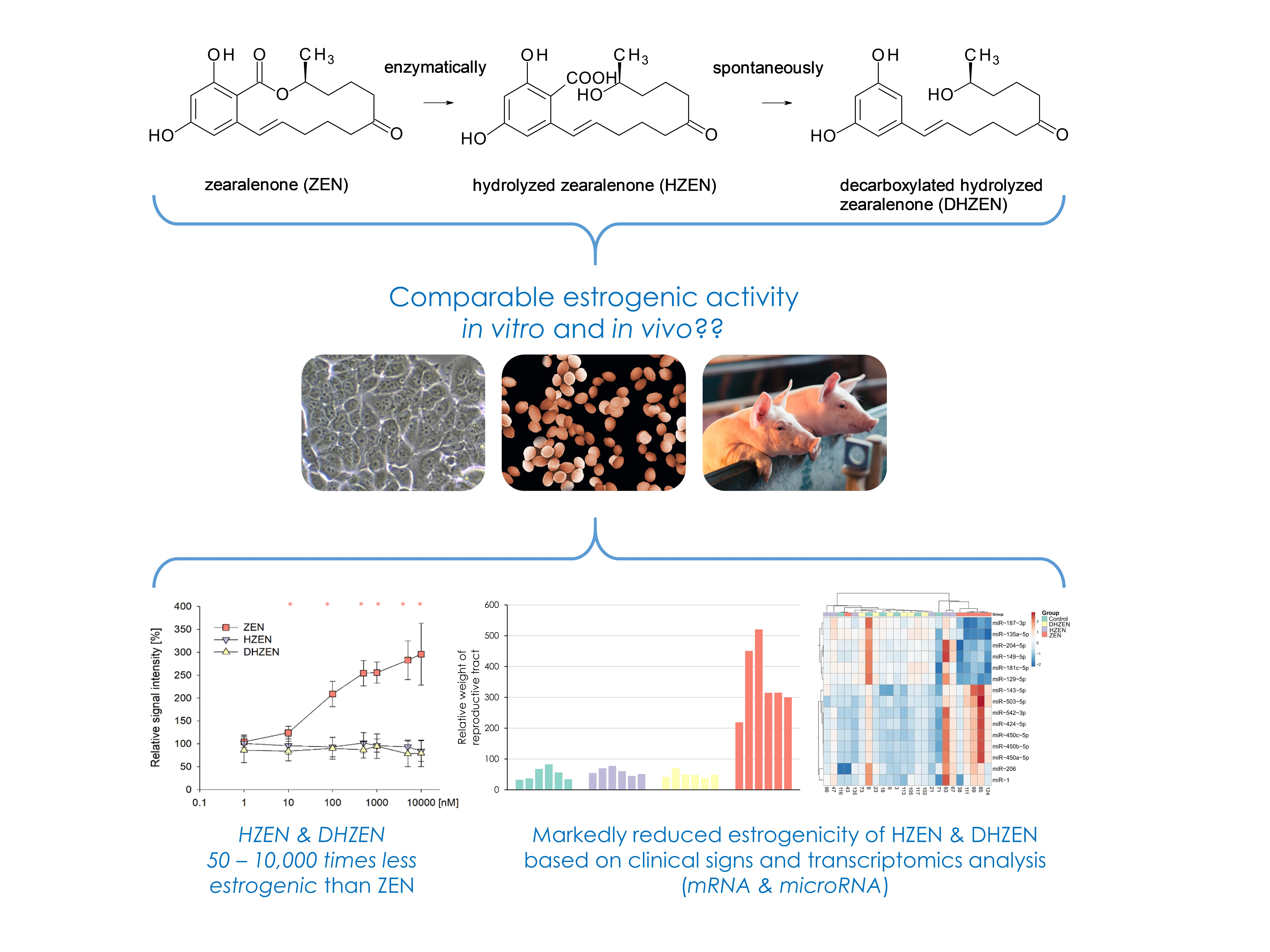
:
1. Introduction
2. Results
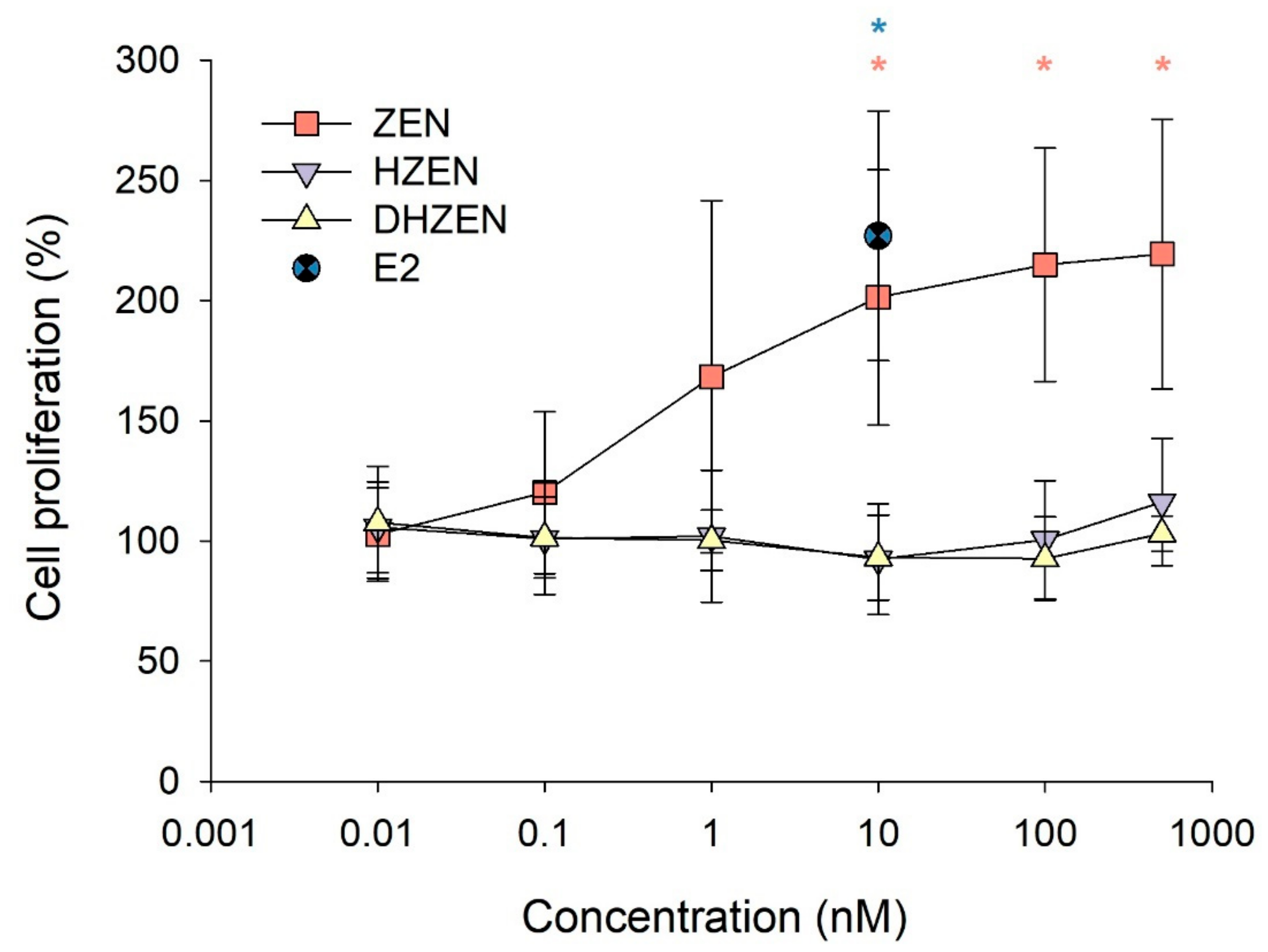
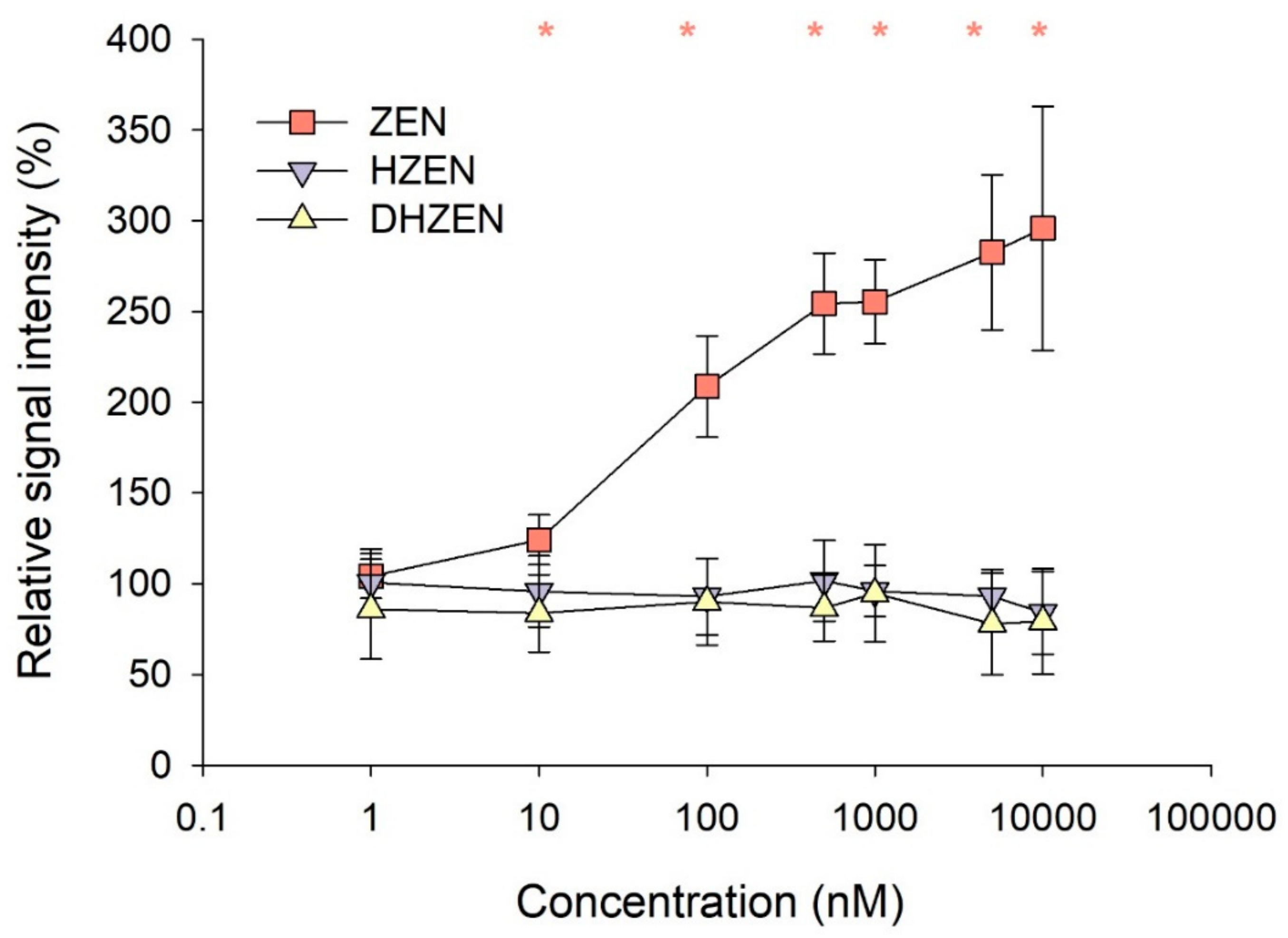
2.1. In Vitro Experiments
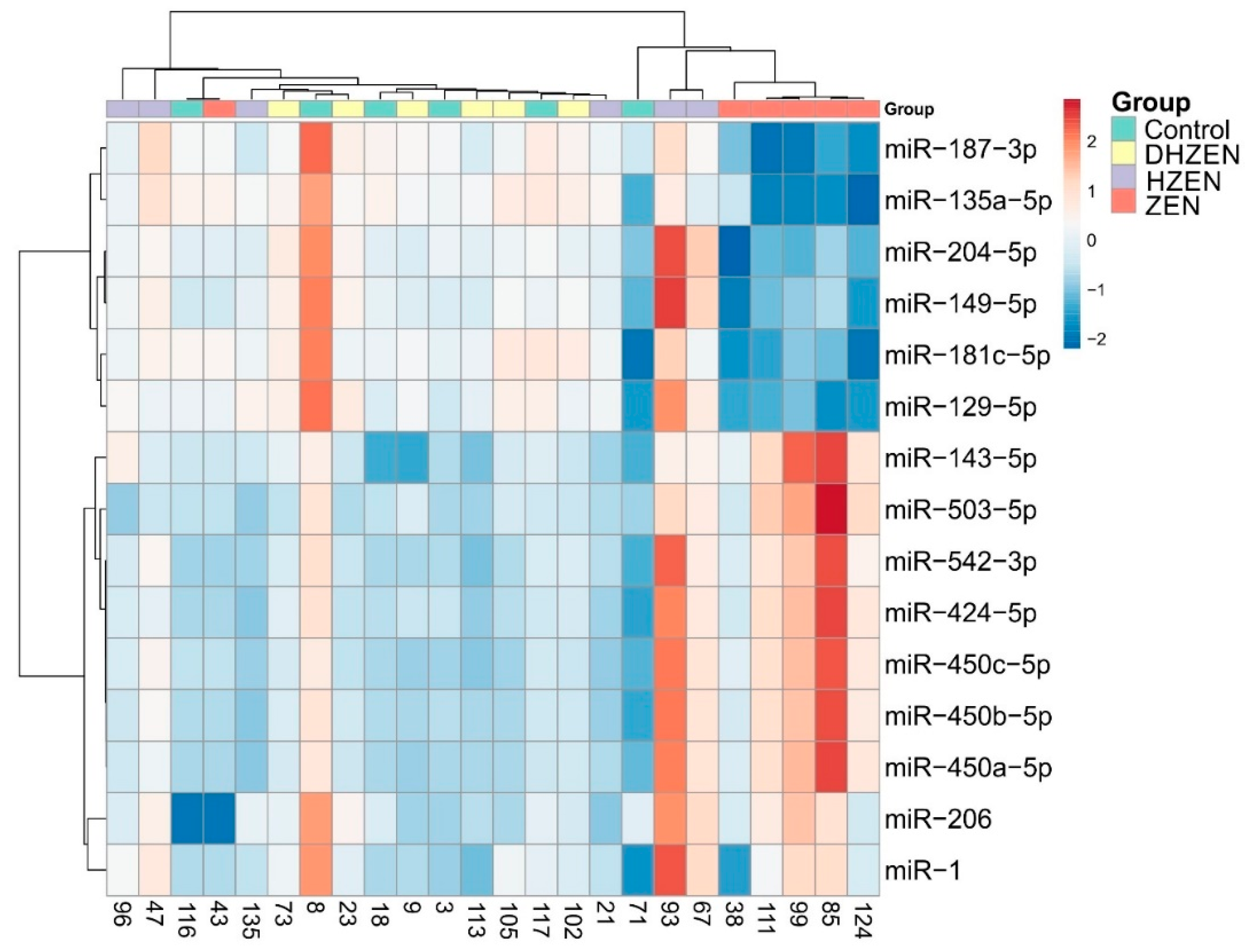
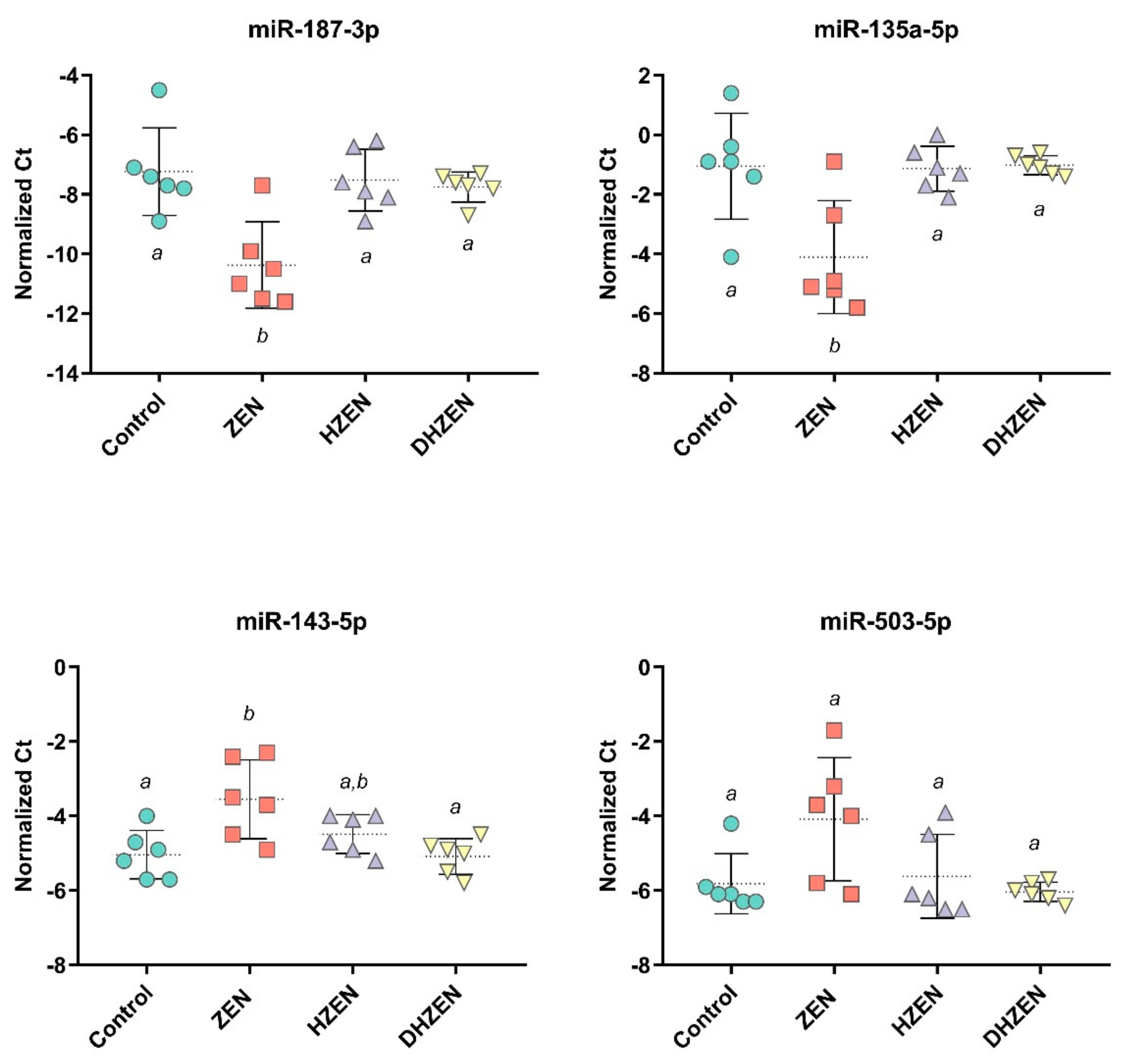
2.2. In Vivo Experiment
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. In Vitro Experiments
5.1.1. Chemicals, Reagents and Materials
5.1.2. Production of HZEN and DHZEN
5.1.3. MCF-7 Proliferation Assay
5.1.4. Yeast Bioassay
5.2. In Vivo Experiment
5.2.1. Animals and Study Design
5.2.2. qPCR Analysis of mRNAs From Genes with Potential ERE or Associated with Estrogen Response
5.2.3. qPCR Analysis of microRNAs
5.3. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zinedine, A.; Soriano, J.M.; Molto, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Rodrigues, I.; Naehrer, K. Prevalence of mycotoxins in feedstuffs and feed surveyed worldwide in 2009 and 2010. Phytopathol. Mediterr. 2012, 51, 175–192. [Google Scholar]
- Kovalsky, P.; Kos, G.; Nährer, K.; Schwab, C.; Jenkins, T.; Schatzmayr, G.; Sulyok, M.; Krska, R. Co-Occurrence of Regulated, Masked and Emerging Mycotoxins and Secondary Metabolites in Finished Feed and Maize—An Extensive Survey. Toxins 2016, 8, 363. [Google Scholar] [CrossRef] [PubMed]
- Fink-Gremmels, J.; Malekinejad, H. Clinical effects and biochemical mechanisms associated with exposure to the mycoestrogen zearalenone. Anim. Feed Sci. Technol. 2007, 137, 326–341. [Google Scholar] [CrossRef]
- Vrtačnik, P.; Ostanek, B.; Mencej-Bedrač, S.; Marc, J. The many faces of estrogen signaling. Biochem. Med. 2014, 24, 329–342. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Piastowska-Ciesielska, A.W. Zearalenone as an endocrine disruptor in humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Hennig-Pauka, I.; Koch, F.-J.; Schaumberger, S.; Woechtl, B.; Novak, J.; Sulyok, M.; Nagl, V. Current challenges in the diagnosis of zearalenone toxicosis as illustrated by a field case of hyperestrogenism in suckling piglets. Porc. Health Manag. 2018, 4, 18. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wu, W.; Li, L.; He, J.; Huang, S.; Chen, S.; Chen, J.; Long, M.; Yang, S.; Li, P. Analysis of the miRNA Expression Profiles in the Zearalenone-Exposed TM3 Leydig Cell Line. Int. J. Mol. Sci. 2019, 20, 635. [Google Scholar] [CrossRef] [PubMed]
- Brzuzan, P.; Woźny, M.; Wolińska-Nizioł, L.; Piasecka, A.; Florczyk, M.; Jakimiuk, E.; Góra, M.; Łuczyński, M.; Gajęcki, M. MicroRNA expression profiles in liver and colon of sexually immature gilts after exposure to Fusarium mycotoxins. Pol. J. Vet. Sci. 2015, 18, 29–38. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Zhang, J.; Wang, Y.; Liu, W.; Gou, K.; Liu, Z.; Cui, S. MiR-7 Mediates the Zearalenone Signaling Pathway Regulating FSH Synthesis and Secretion by Targeting FOS in Female Pigs. Endocrinology 2018, 159, 2993–3006. [Google Scholar] [CrossRef]
- Grenier, B.; Hackl, M.; Skalicky, S.; Thamhesl, M.; Moll, W.-D.; Berrios, R.; Schatzmayr, G.; Nagl, V. MicroRNAs in porcine uterus and serum are affected by zearalenone and represent a new target for mycotoxin biomarker discovery. Sci. Rep. 2019, 9, 9408. [Google Scholar] [CrossRef] [PubMed]
- Schraml, E.; Hackl, M.; Grillari, J. Micrornas and toxicology: A love marriage micrornas in liquid biopsies are minimal-invasive biomarkers for tissue-specific toxicity. Toxicol. Rep. 2017, 4, 634–636. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Wei, C.; Li, Y.; Liu, Y.; Wang, Y.; Pan, J.; Liu, J.; Wu, Y.; Cui, S. Zearalenone and alpha-zearalenol inhibit the synthesis and secretion of pig follicle stimulating hormone via the non-classical estrogen membrane receptor GPR30. Mol. Cell. Endocrinol. 2018, 461, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Ayed-Boussema, I.; Pascussi, J.M.; Maurel, P.; Bacha, H.; Hassen, W. Zearalenone activates pregnane X receptor, constitutive androstane receptor and aryl hydrocarbon receptor and corresponding phase I target genes mRNA in primary cultures of human hepatocytes. Environ. Toxicol. Pharmacol. 2011, 31, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. Activity of Zearalenone in the Porcine Intestinal Tract. Molecules 2016, 22, 18. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Wang, B.; Li, X.; Wang, T.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Bai, J.; Bian, J.; et al. Zearalenone Promotes Cell Proliferation or Causes Cell Death? Toxins 2018, 10, 184. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission recommendation of of 17 august 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin a, t-2 and ht-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, L 229, 7–9. [Google Scholar]
- Alassane-Kpembi, I.; Schatzmayr, G.; Taranu, I.; Marin, D.; Puel, O.; Oswald, I.P. Mycotoxins co-contamination: Methodological aspects and biological relevance of combined toxicity studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 3489–3507. [Google Scholar] [CrossRef]
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarc’H, A.; Lebrihi, A. Review of mycotoxin reduction in food and feed: From prevention in the field to detoxification by adsorption or transformation. Food Addit. Contam. Part A 2011, 28, 1590–1609. [Google Scholar] [CrossRef]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: Effect on human health, metabolism and neutralisation methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef]
- Ji, C.; Fan, Y.; Zhao, L. Review on biological degradation of mycotoxins. Anim. Nutr. 2016, 2, 127–133. [Google Scholar] [CrossRef]
- Di Gregorio, M.C.; De Neeff, D.V.; Jager, A.V.; Corassin, C.H.; Carão, Á.C.D.P.; De Albuquerque, R.; De Azevedo, A.C.; Oliveira, C.A.F. Mineral adsorbents for prevention of mycotoxins in animal feeds. Toxin Rev. 2014, 33, 1–11. [Google Scholar] [CrossRef]
- Fruhauf, S.; Schwartz, H.; Ottner, F.; Krska, R.; Vekiru, E. Yeast cell based feed additives: Studies on aflatoxin B1and zearalenone. Food Addit. Contam. Part A 2012, 29, 217–231. [Google Scholar] [CrossRef]
- De Baere, S.; De Mil, T.; Antonissen, G.; Devreese, M.; Croubels, S. In vitro model to assess the adsorption of oral veterinary drugs to mycotoxin binders in a feed- and aflatoxin B1-containing buffered matrix. Food Addit. Contam. Part A 2018, 35, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kiessling, K.H.; Pettersson, H.; Sandholm, K.; Olsen, M. Metabolism of aflatoxin, ochratoxin, zearalenone, and three trichothecenes by intact rumen fluid, rumen protozoa, and rumen bacteria. Appl. Environ. Microbiol. 1984, 47, 1070–1073. [Google Scholar] [PubMed]
- Fitzpatrick, D.; Picken, C.; Murphy, L.C.; Buhr, M. Measurement of the relative binding affinity of zearalenone, α-zearalenol and β-zearalenol for uterine and oviduct estrogen receptors in swine, rats and chickens: An indicator of estrogenic potencies. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1989, 94, 691–694. [Google Scholar] [CrossRef]
- El-Sharkawy, S.; Abul-Hajj, Y. Microbial Transformation of Zearalenone, I. Formation of Zearalenone-4-O-β-glucoside. J. Nat. Prod. 1987, 50, 520–521. [Google Scholar] [CrossRef]
- Brodehl, A.; Möller, A.; Kunte, H.-J.; Koch, M.; Maul, R. Biotransformation of the mycotoxin zearalenone by fungi of the genera Rhizopus and Aspergillus. FEMS Microbiol. Lett. 2014, 359, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Binder, S.B.; Schwartz-Zimmermann, H.E.; Varga, E.; Bichl, G.; Michlmayr, H.; Adam, G.; Berthiller, F. Metabolism of Zearalenone and Its Major Modified Forms in Pigs. Toxins 2017, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Vekiru, E.; Frühauf, S.; Hametner, C.; Schatzmayr, G.; Krska, R.; Moll, W.; Schuhmacher, R. Isolation and characterisation of enzymatic zearalenone hydrolysis reaction products. World Mycotoxin J. 2016, 9, 353–363. [Google Scholar] [CrossRef]
- Vekiru, E.; Hametner, C.; Mitterbauer, R.; Rechthaler, J.; Adam, G.; Schatzmayr, G.; Krska, R.; Schuhmacher, R. Cleavage of Zearalenone by Trichosporon mycotoxinivorans to a Novel Nonestrogenic Metabolite. Appl. Environ. Microbiol. 2010, 76, 2353–2359. [Google Scholar] [CrossRef] [PubMed]
- Kakeya, H.; Takahashi-Ando, N.; Kimura, M.; Onose, R.; Yamaguchi, I.; Osada, H. Biotransformation of the Mycotoxin, Zearalenone, to a Non-estrogenic Compound by a Fungal Strain of Clonostachys sp. Biosci. Biotechnol. Biochem. 2002, 66, 2723–2726. [Google Scholar] [CrossRef]
- El-Sharkawy, S.; Abul-Hajj, Y.J. Microbial cleavage of zearalenone. Xenobiotica 1988, 18, 365–371. [Google Scholar] [CrossRef]
- Takahashi-Ando, N.; Ohsato, S.; Shibata, T.; Hamamoto, H.; Yamaguchi, I.; Kimura, M. Metabolism of Zearalenone by Genetically Modified Organisms Expressing the Detoxification Gene from Clonostachys rosea. Appl. Environ. Microbiol. 2004, 70, 3239–3245. [Google Scholar] [CrossRef] [PubMed]
- Bajic, V.B.; Tan, S.L.; Chong, A.; Tang, S.; Ström, A.; Gustafsson, J.-A.; Lin, C.-Y.; Liu, E.T. Dragon ERE Finder version 2: A tool for accurate detection and analysis of estrogen response elements in vertebrate genomes. Nucleic Acids Res. 2003, 31, 3605–3607. [Google Scholar] [CrossRef]
- Tatay, E.; Espín, S.; García-Fernández, A.-J.; Ruiz, M.-J. Estrogenic activity of zearalenone, α-zearalenol and β-zearalenol assessed using the e-screen assay in mcf-7 cells. Toxicol. Mech. Methods 2018, 28, 239–242. [Google Scholar] [CrossRef]
- LeComte, S.; Lelong, M.; Bourgine, G.; Efstathiou, T.; Saligaut, C.; Pakdel, F. Assessment of the potential activity of major dietary compounds as selective estrogen receptor modulators in two distinct cell models for proliferation and differentiation. Toxicol. Appl. Pharmacol. 2017, 325, 61–70. [Google Scholar] [CrossRef]
- Hessenberger, S.; Botzi, K.; Degrassi, C.; Kovalsky, P.; Schwab, C.; Schatzmayr, D.; Schatzmayr, G.; Fink-Gremmels, J. Interactions between plant-derived oestrogenic substances and the mycoestrogen zearalenone in a bioassay with MCF-7 cells. Pol. J. Vet. Sci. 2017, 20, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosrokhavar, R.; Rahimifard, N.; Shoeibi, S.; Hamedani, M.P.; Hosseini, M.-J. Effects of zearalenone and α-Zearalenol in comparison with Raloxifene on T47D cells. Toxicol. Mech. Methods 2009, 19, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Andersen, H.R.; Andersson, A.-M.; Arnold, S.F.; Autrup, H.; Barfoed, M.; Beresford, N.A.; Bjerregaard, P.; Christiansen, L.B.; Gissel, B.; Hummel, R.; et al. Comparison of Short-Term Estrogenicity Tests for Identification of Hormone-Disrupting Chemicals. Environ. Health Perspect. 1999, 107, 89–108. [Google Scholar] [CrossRef]
- Doll, S.; Dänicke, S.; Ueberschär, K.-H.; Valenta, H.; Schnurrbusch, U.; Ganter, M.; Klobasa, F.; Flachowsky, G. Effects of graded levels of Fusarium toxin contaminated maize in diets for female weaned piglets. Arch. Anim. Nutr. 2003, 57, 311–334. [Google Scholar] [CrossRef]
- Zöllner, P.; Jodlbauer, J.; Kleinova, M.; Kahlbacher, H.; Kuhn, T.; Hochsteiner, W.; Lindner, W. Concentration Levels of Zearalenone and Its Metabolites in Urine, Muscle Tissue, and Liver Samples of Pigs Fed with Mycotoxin-Contaminated Oats. J. Agric. Food Chem. 2002, 50, 2494–2501. [Google Scholar] [CrossRef] [PubMed]
- Frizzell, C.; Uhlig, S.; Miles, C.O.; Verhaegen, S.; Elliott, C.T.; Eriksen, G.S.; Sørlie, M.; Ropstad, E.; Connolly, L. Biotransformation of zearalenone and zearalenols to their major glucuronide metabolites reduces estrogenic activity. Toxicol. In Vitro 2015, 29, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Faisal, Z.; Lemli, B.; Szerencsés, D.; Kunsági-Máté, S.; Bálint, M.; Hetényi, C.; Kuzma, M.; Mayer, M.; Poór, M. Interactions of zearalenone and its reduced metabolites α-zearalenol and β-zearalenol with serum albumins: Species differences, binding sites, and thermodynamics. Mycotoxin Res. 2018, 34, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Yang, Z.; Yang, W.; Yao, B.; Zhao, H.; Liu, F.; Chen, C.; Chi, F. Effects of feeding purified zearalenone contaminated diets with or without clay enterosorbent on growth, nutrient availability, and genital organs in post-weaning female pigs. Asian-Aust. J. Anim. Sci. 2010, 23, 74–81. [Google Scholar] [CrossRef]
- Grenier, B.; Nagl, V.; Lutz, A.; Aleschko, M.; Schatzmayr, G.; Moll, W.-D.; Thamhesl, M. Targeted approaches for zen biomarker discovery in prepubertal gilts. In Proceedings of the 10th World Mycotoxin Forum, Amsterdam, The Netherlands, 12–14 March 2018. [Google Scholar]
- Li, Y.; Burns, K.A.; Arao, Y.; Luh, C.J.; Korach, K.S. Differential Estrogenic Actions of Endocrine-Disrupting Chemicals Bisphenol A, Bisphenol AF, and Zearalenone through Estrogen Receptor α and β in Vitro. Environ. Health Perspect. 2012, 120, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Bourdeau, V.; Deschênes, J.; Métivier, R.; Nagai, Y.; Nguyen, D.; Bretschneider, N.; Gannon, F.; White, J.H.; Mader, S. Genome-Wide Identification of High-Affinity Estrogen Response Elements in Human and Mouse. Mol. Endocrinol. 2004, 18, 1411–1427. [Google Scholar] [CrossRef] [Green Version]
- Jung, E.-M.; An, B.-S.; Yang, H.; Choi, K.-C.; Jeung, E.-B. Biomarker Genes for Detecting Estrogenic Activity of Endocrine Disruptors via Estrogen Receptors. Int. J. Environ. Res. Public Health 2012, 9, 698–711. [Google Scholar] [CrossRef] [Green Version]
- Zou, K.; Ing, N.H. Oestradiol up-regulates oestrogen receptor, cyclophilin, and glyceraldehyde phosphate dehydrogenase mRNA concentrations in endometrium, but down-regulates them in liver. J. Steroid Biochem. Mol. Boil. 1998, 64, 231–237. [Google Scholar] [CrossRef]
- LeComte, S.; DeMay, F.; Pham, T.H.; Moulis, S.; Efstathiou, T.; Chalmel, F.; Pakdel, F. Deciphering the Molecular Mechanisms Sustaining the Estrogenic Activity of the Two Major Dietary Compounds Zearalenone and Apigenin in ER-Positive Breast Cancer Cell Lines. Nutrients 2019, 11, 237. [Google Scholar] [CrossRef]
- Vo, T.T.B.; Jeung, E.-B. An Evaluation of Estrogenic Activity of Parabens Using Uterine Calbindin-D9k Gene in an Immature Rat Model. Toxicol. Sci. 2009, 112, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Krisinger, J. Porcine calbindin-D9k gene: Expression in endometrium, myometrium, and placenta in the absence of a functional estrogen response element in intron A. Boil. Reprod. 1995, 52, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Oliver, W.; Miles, J.; Diaz, D.; Dibner, J.; Rottinghaus, G.; Harrell, R. Zearalenone enhances reproductive tract development, but does not alter skeletal muscle signaling in prepubertal gilts. Anim. Feed Sci. Technol. 2012, 174, 79–85. [Google Scholar] [CrossRef]
- Klinge, C.M. miRNAs regulated by estrogens, tamoxifen, and endocrine disruptors and their downstream gene targets. Mol. Cell. Endocrinol. 2015, 418, 273–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Madak-Erdogan, Z.; Ventrella, R.; Katzenellenbogen, B.S. A microrna196a2* and tp63 circuit regulated by estrogen receptor-α and erk2 that controls breast cancer proliferation and invasiveness properties. Horm. Cancer 2013, 4, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, L.; Ravo, M.; Nassa, G.; Tarallo, R.; De Filippo, M.R.; Giurato, G.; Cirillo, F.; Stellato, C.; Silvestro, S.; Cantarella, C.; et al. Effects of Oestrogen on MicroRNA Expression in Hormone-Responsive Breast Cancer Cells. Horm. Cancer 2012, 3, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Tribollet, V.; Barenton, B.; Kroiss, A.; Vincent, S.; Zhang, L.; Forcet, C.; Cerutti, C.; Périan, S.; Allioli, N.; Samarut, J.; et al. miR-135a Inhibits the Invasion of Cancer Cells via Suppression of ERRα. PLoS ONE 2016, 11, e0156445. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, J.; Wang, H.; Zhao, J.; Xu, C.; Du, Y.; Luo, X.; Zheng, F.; Liu, R.; Zhang, H.; et al. miRNA-135a promotes breast cancer cell migration and invasion by targeting HOXA10. BMC Cancer 2012, 12, 111. [Google Scholar] [CrossRef]
- Guo, S.; Yang, J.; Wu, M.; Xiao, G. Clinical value screening, prognostic significance and key pathway identification of miR-204-5p in endometrial carcinoma: A study based on the Cancer Genome Atlas (TCGA), and bioinformatics analysis. Pathol. Res. Pr. 2019, 215, 1003–1011. [Google Scholar] [CrossRef]
- Nothnick, W.B.; Healy, C. Estrogen induces distinct patterns of microRNA expression within the mouse uterus. Reprod. Sci. 2010, 17, 987–994. [Google Scholar] [CrossRef]
- Baran-Gale, J.; Purvis, J.E.; Sethupathy, P. An integrative transcriptomics approach identifies miR-503 as a candidate master regulator of the estrogen response in MCF-7 breast cancer cells. RNA 2016, 22, 1592–1603. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Scientific Opinion Guidance for the preparation of dossiers for technological additives. EFSA J. 2012, 10, 2528. [Google Scholar] [CrossRef]
- Reg (EC) No 1831/2003. European Union Register of Feed Additives. Edition 6/2019 (272). Appendixes 3e, 4–26 June 2019. Available online: https://ec.europa.eu/food/safety/animal-feed/feed-additives_en (accessed on 7 August 2019).
- Hartinger, D.; Moll, W.-D. Fumonisin elimination and prospects for detoxification by enzymatic transformation. World Mycotoxin J. 2011, 4, 271–283. [Google Scholar] [CrossRef]
- Alberts, J.; Lilly, M.; Rheeder, J.; Burger, H.; Shephard, G.S.; Gelderblom, W. Technological and community-based methods to reduce mycotoxin exposure. Food Control 2017, 73, 101–109. [Google Scholar] [CrossRef]
- Hahn, I.; Kunz-Vekiru, E.; Twaruzek, M.; Grajewski, J.; Krska, R.; Berthiller, F. Aerobic and anaerobic in vitro testing of feed additives claiming to detoxify deoxynivalenol and zearalenone. Food Addit. Contam. Part A 2015, 32, 922–933. [Google Scholar] [CrossRef] [PubMed]
- James, P.; Halladay, J.; Craig, E.A. Genomic Libraries and a Host Strain Designed for Highly Efficient Two-Hybrid Selection in Yeast. Genetics 1996, 144, 1425–1436. [Google Scholar] [PubMed]
- Mitterbauer, R.; Weindorfer, H.; Safaie, N.; Krska, R.; Lemmens, M.; Ruckenbauer, P.; Kuchler, K.; Adam, G. A Sensitive and Inexpensive Yeast Bioassay for the Mycotoxin Zearalenone and Other Compounds with Estrogenic Activity. Appl. Environ. Microbiol. 2003, 69, 805–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachmann, H. Phenotypic Detection of Zearalenone in Saccharomyces Cerevisiae. Master’s Thesis, University of Natural Resources and Life Sciences, Vienna (BOKU), Vienna, Austria, 2001. [Google Scholar]
- Sherman, F. Getting started with yeast. Methods Enzymol 2002, 350, 3–41. [Google Scholar] [PubMed]
- European Commission. Directive 2010/63/eu of the European Parliament and of the Council of of 22 September 2010 on the Protection of Animals Used for Scientific Purposes. Off. J. Eur. Union 2010, L 276, 33–79. [Google Scholar]
- Klinge, C.M. Estrogen receptor interaction with estrogen response elements. Nucleic Acids Res. 2001, 29, 2905–2919. [Google Scholar] [CrossRef] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Boil. 2002, 3, research0034.1. [Google Scholar]
- Nygard, A.-B.; Jorgensen, C.B.; Cirera, S.; Fredholm, M. Selection of reference genes for gene expression studies in pig tissues using SYBR green qPCR. BMC Mol. Boil. 2007, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Loureiro-Bracarense, A.-P.; Lucioli, J.; Pacheco, G.D.; Cossalter, A.-M.; Moll, W.-D.; Schatzmayr, G.; Oswald, I.P.; Loureiro-Bracarense, A. Individual and combined effects of subclinical doses of deoxynivalenol and fumonisins in piglets. Mol. Nutr. Food Res. 2011, 55, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Gessner, D.K.; Fiesel, A.; Most, E.; Dinges, J.; Wen, G.; Ringseis, R.; Eder, K. Supplementation of a grape seed and grape marc meal extract decreases activities of the oxidative stress-responsive transcription factors nf-κb and nrf2 in the duodenal mucosa of pigs. Acta Vet. Scand. 2013, 55, 18. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Body Weight (kg) | Vulva Size 1 (cm2) | Reproductive Tract Weight 2 |
|---|---|---|---|
| Control | 20.5 ± 2.8 | 1.47 ± 0.42a | 51.8 ± 20.6a |
| ZEN | 21.0 ± 2.2 | 4.84 ± 0.66b | 353.4 ± 45.2b |
| HZEN | 20.7 ± 2.5 | 1.70 ± 0.24a | 59.8 ± 12.0a |
| DHZEN | 20.5 ± 4.0 | 1.41 ± 0.27a | 49.9 ± 11.1a |
| Gene Name | Protein Name | Relative Gene Expression Mean (and Range) Fold-Change Compared to Control | |||
|---|---|---|---|---|---|
| Control | ZEN | HZEN | DHZEN | ||
| EBAG9 | Receptor-binding cancer antigen expressed on SiSo cells | 1.00 a (0.61–1.65) | 1.47 b (1.01–2.16) | 1.04 a (0.67–1.61) | 1.05 a (0.92–1.20) |
| OVGP1 | Oviduct-specific glycoprotein or mucin-9 | 1.00 (0.54–1.85) | 2.19 (1.03–4.70) | 1.35 (0.68–2.67) | 1.79 (1.07–3.00) |
| IGFBP4 | Insulin-like growth factor-binding protein 4 | 1.00 (0.59–1.71) | 0.57 (0.29–1.15) | 1.16 (0.70–1.94) | 1.23 (0.93–1.63) |
| GJA1 | Gap junction alpha-1 protein or connexin 43 | 1.00 a (0.62–1.62) | 1.93 b (1.48–2.52) | 1.12 a,b (0.65–1.93) | 1.26 a,b (0.98–1.62) |
| GAPDH | Glyceraldehyde 3phosphate dehydrogenase | 1.00 a,b (0.46–2.15) | 1.97 a (1.15–3.37) | 1.08 a,b (0.48–2.43) | 0.92 b (0.58–1.45) |
| C3 | Complement component 3 | 1.00 (0.41–2.42) | 0.78 (0.40–1.53) | 0.86 (0.44–1.69) | 1.40 (1.11–1.76) |
| S100G | S100 calciumbinding protein or calbindin D9K | 1.00 a,b (0.13–7.72) | 0.22 a (0.02–2.44) | 2.48 a,b (1.68–3.66) | 4.32 b (2.18–8.54) |
| CLU | Clusterin or apolipoprotein J | 1.00 (0.53–1.90) | 1.27 (0.65–2.50) | 0.85 (0.53–1.35) | 1.02 (0.73–1.44) |
| ODC | Ornithine decarboxylase | 1.00 (0.55–1.82) | 0.95 (0.71–1.28) | 0.95 (0.62–1.45) | 1.09 (0.89–1.33) |
| ESR1 | Estrogen receptor alpha | 1.00 (0.57–1.74) | 1.11 (0.73–1.68) | 1.18 (0.66–2.10) | 1.24 (1.06–1.45) |
| ESR2 | Estrogen receptor beta | 1.00 (0.39–2.54) | 1.22 (0.33–4.51) | 0.40 (0.09–1.90) | 1.62 (0.53–4.94) |
| IL-1β | Interleukin 1 beta | 1.00 (0.26–3.90) | 3.36 (0.47–24.12) | 1.23 (0.43–3.52) | 1.83 (1.12–3.01) |
| IL-6 | Interleukin 6 | 1.00 (0.56–1.78) | 0.89 (0.34–2.36) | 1.40 (0.85–2.30) | 1.90 (1.25–2.92) |
| microRNA | Relative microRNA Expression Mean (and Range) Fold-Change Compared to Control | |||
|---|---|---|---|---|
| Control | ZEN | HZEN | DHZEN | |
| miR-1 | 1.00 (0.47–2.11) | 1.27 (0.72–2.22) | 1.89 (0.63–5.66) | 1.03 (0.67–1.60) |
| miR-129-5p | 1.00 a,b (0.45–2.24) | 0.42 a (0.18–0.99) | 1.44 b (0.40–5.21) | 1.19 b (0.78–1.82) |
| miR-135a-5p | 1.00 a (0.32–3.11) | 0.12 b (0.02–0.68) | 0.94 a (0.21–4.27) | 1.01 a (0.63–1.62) |
| miR-143-5p | 1.00 a (0.47–2.14) | 2.78 b (1.44–5.36) | 1.45 a,b (0.38–5.48) | 0.96 a (0.65–1.43) |
| miR-149-5p | 1.00 a,b (0.46–2.18) | 0.37 a (0.19–0.74) | 1.77 b (0.59–5.31) | 1.11 a,b (0.76–1.63) |
| miR-181c-5p | 1.00 (0.40–2.52) | 0.40 (0.17–0.96) | 1.12 (0.30–4.24) | 1.14 (0.70–1.87) |
| miR-187-3p | 1.00 a (0.44–2.29) | 0.11 b (0.03–0.48) | 0.81 a (0.23–2.85) | 0.70 a (0.42–1.16) |
| miR-204-5p | 1.00 a (0.45–2.22) | 0.29 b (0.13–0.66) | 1.58 a (0.54–4.65) | 0.99 a (0.66–1.47) |
| miR-206 | 1.00 (0.36–2.79) | 1.34 (0.45–4.04) | 1.88 (0.61–5.81) | 0.88 (0.56–1.39) |
| miR-424-5p | 1.00 (0.47–2.15) | 3.69 (1.31–10.41) | 1.86 (0.63–5.47) | 0.95 (0.57–1.57) |
| miR-450a-5p | 1.00 (0.50–1.98) | 4.25 (1.45–12.42) | 2.11 (0.72–6.21) | 0.92 (0.60–1.42) |
| miR-450b-5p | 1.00 (0.48–2.09) | 3.63 (1.34–9.84) | 2.00 (0.68–5.86) | 0.94 (0.61–1.44) |
| miR-450c-5p | 1.00 a,b (0.49–2.06) | 4.52 a (1.60–12.75) | 2.36 a,b (0.78–7.12) | 0.84 b (0.54–1.30) |
| miR-503-5p | 1.00 (0.53–1.89) | 3.36 (1.29–8.74) | 1.16 (0.38–3.51) | 0.87 (0.49–1.54) |
| miR-542-3p | 1.00 (0.47–2.12) | 3.45 (1.20–9.89) | 2.19 (0.72–6.63) | 0.88 (0.54–1.42) |
| Gene Name | Primer Sequence | Amplicon Size | Ensembl Access # |
|---|---|---|---|
| Housekeeping Genes | |||
| HPRT1 1 | F (300 nM) GGACTTGAATCATGTTTGTG R (300 nM) CAGATGTTTCCAAACTCAAC | 91 bp | ENSSSCG00000034896 |
| RPL32 1 | F (300 nM) AGTTCATCCGGCACCAGTCA R (300 nM) GAACCTTCTCCGCACCCTGT | 92 bp | ENSSSCG00000035811 |
| RPL4 1 | F (300 nM) CAAGAGTAACTACAACCTTC R (300 nM) GAACTCTACGATGAATCTTC | 122 bp | ENSSSCG00000004945 |
| TBP 1 | F (300 nM) AACAGTTCAGTAGTTATGAGCCAGA R (300 nM) AGATGTTCTCAAACGCTTCG | 153 bp | ENSSSCG00000037372 |
| Genes with Identified ERE Motif | |||
| EBAG9 | F (300 nM) GCACAGGTTTCTCTAGTAGGCT R (300 nM) TCCCTGTCTGCTATCTTCTGC | 175 bp | ENSSSCG00000006024 |
| OVGP1 | F (300 nM) GGGTCGGCTATGATGATGACA R (300 nM) CCGGTGAAGGAGTTGAGCTA | 198 bp | ENSSSCG00000006791 |
| IGFBP4 | F (300 nM) CATCCCCATCCCTAACTGCG R (300 nM) CTCACTCTCGGAAGCTGTCG | 185 bp | ENSSSCG00000017472 |
| GJA1 | F (300 nM) TCTGAGTGCCTGAACTTGCT R (300 nM) CAGCGGTGGAATAGGCTTGA | 154 bp | ENSSSCG00000004241 |
| GAPDH 1,2 | F (300 nM) AGGGGCTCTCCAGAACATCATCC R (300 nM) TCGCGTGCTCTTGCTGGGGTTGG | 446 bp | ENSSSCG00000000694 |
| C3 2 | F (300 nM) GGGCAGATCTTGAGTGTCCG R (300 nM) ATGCTGGATGAACTGAGCCC | 179 bp | ENSSSCG00000013551 |
| Other Genes Associated With Estrogenic Response or Inflammation | |||
| S100G | F (300 nM) GGAGTTGAACTTGGACGTGC R (300 nM) CGCATCCCTTCCAGTCCTTA | 184 bp | ENSSSCG00000012147 |
| CLU | F (300 nM) CCATGACATGTTCCAGCCCT R (300 nM) TCTGAGAGGAATTGCTGGCC | 239 bp | ENSSSCG00000009668 |
| ODC | F (300 nM) CGGCGATTGGATGCTCTTTG R (300 nM) AAGTCGTGGTTCCGGATCTG | 144 bp | ENSSSCG00000027121 |
| ESR1 | F (300 nM) CCTGGAGAATGAGCCGAGC R (300 nM) CTTCCCTTGTCACTGGTGCT | 92 bp | ENSSSCG00000025777 |
| ESR2 | F (300 nM) TGCAGTGATTATGCGTCAGGA R (300 nM) CAGCTTTTACGCCGGTTCTT | 149 bp | ENSSSCG00000005109 |
| IL-1β | F (300 nM) CCATAGTACCTGAACCCGCC R (300 nM) GCTGGTGAGAGATTTGCAGC | 165 bp | ENSSSCG00000039214 |
| IL-6 1 | F (300 nM) GGCAAAAGGGAAAGAATCCAG R (300 nM) CGTTCTGTGACTGCAGCTTATCC | 87 bp | ENSSSCG00000020970 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fruhauf, S.; Novak, B.; Nagl, V.; Hackl, M.; Hartinger, D.; Rainer, V.; Labudová, S.; Adam, G.; Aleschko, M.; Moll, W.-D.; et al. Biotransformation of the Mycotoxin Zearalenone to its Metabolites Hydrolyzed Zearalenone (HZEN) and Decarboxylated Hydrolyzed Zearalenone (DHZEN) Diminishes its Estrogenicity In Vitro and In Vivo. Toxins 2019, 11, 481. https://doi.org/10.3390/toxins11080481
Fruhauf S, Novak B, Nagl V, Hackl M, Hartinger D, Rainer V, Labudová S, Adam G, Aleschko M, Moll W-D, et al. Biotransformation of the Mycotoxin Zearalenone to its Metabolites Hydrolyzed Zearalenone (HZEN) and Decarboxylated Hydrolyzed Zearalenone (DHZEN) Diminishes its Estrogenicity In Vitro and In Vivo. Toxins. 2019; 11(8):481. https://doi.org/10.3390/toxins11080481
Chicago/Turabian StyleFruhauf, Sebastian, Barbara Novak, Veronika Nagl, Matthias Hackl, Doris Hartinger, Valentina Rainer, Silvia Labudová, Gerhard Adam, Markus Aleschko, Wulf-Dieter Moll, and et al. 2019. "Biotransformation of the Mycotoxin Zearalenone to its Metabolites Hydrolyzed Zearalenone (HZEN) and Decarboxylated Hydrolyzed Zearalenone (DHZEN) Diminishes its Estrogenicity In Vitro and In Vivo" Toxins 11, no. 8: 481. https://doi.org/10.3390/toxins11080481
APA StyleFruhauf, S., Novak, B., Nagl, V., Hackl, M., Hartinger, D., Rainer, V., Labudová, S., Adam, G., Aleschko, M., Moll, W. -D., Thamhesl, M., & Grenier, B. (2019). Biotransformation of the Mycotoxin Zearalenone to its Metabolites Hydrolyzed Zearalenone (HZEN) and Decarboxylated Hydrolyzed Zearalenone (DHZEN) Diminishes its Estrogenicity In Vitro and In Vivo. Toxins, 11(8), 481. https://doi.org/10.3390/toxins11080481