Influence of Two Garlic-Derived Compounds, Propyl Propane Thiosulfonate (PTS) and Propyl Propane Thiosulfinate (PTSO), on Growth and Mycotoxin Production by Fusarium Species In Vitro and in Stored Cereals


Abstract
:
1. Introduction
2. Results
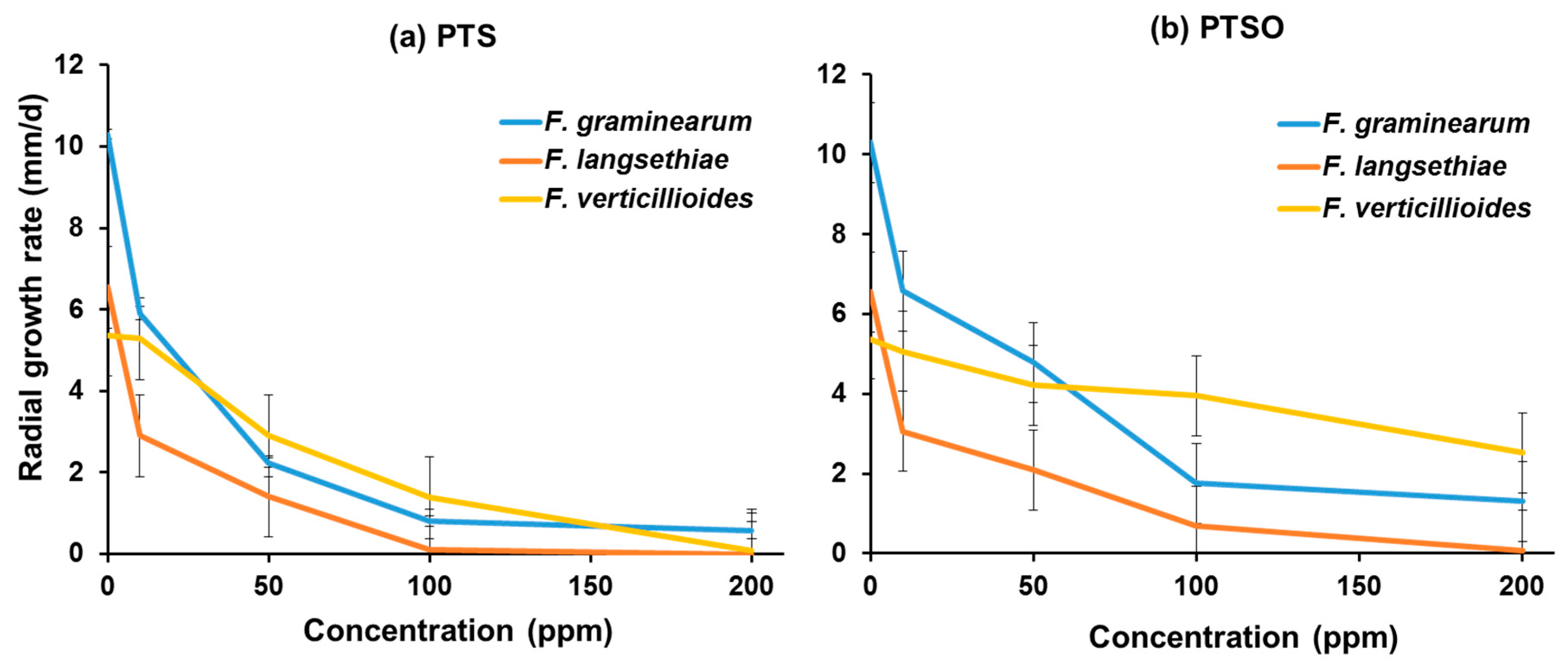
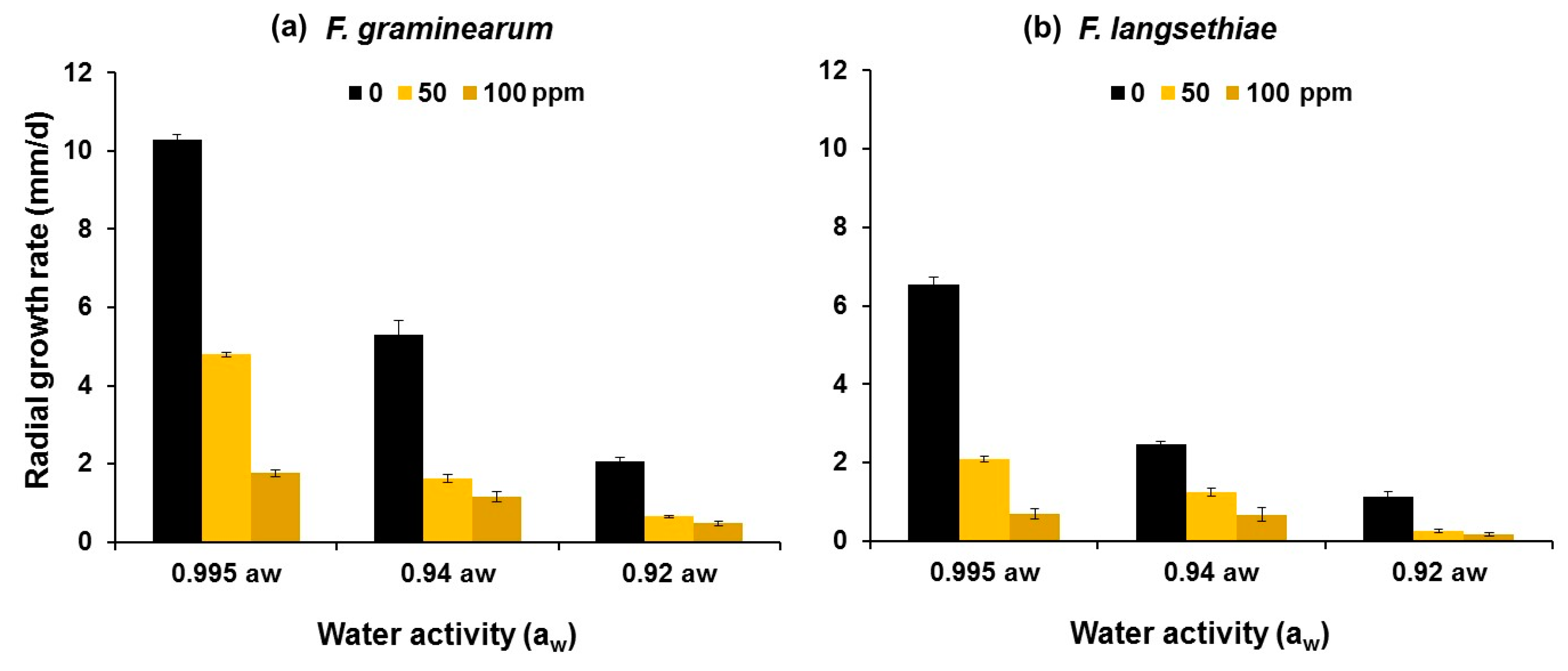
2.1. In Vitro Efficacy of PTS and PTSO Garlic-Derived Compounds against Fungal Growth
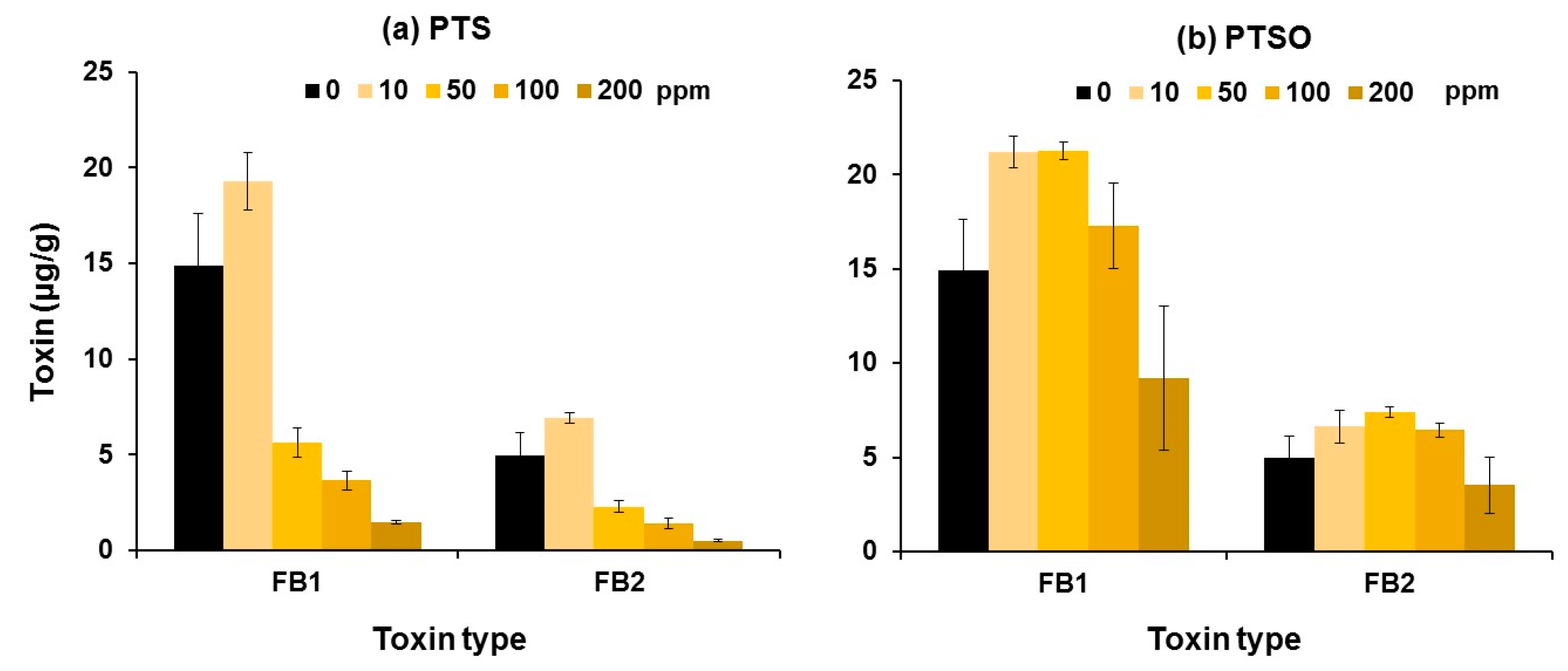
2.2. Effects of the Compounds on In Vitro Mycotoxin Inhibition
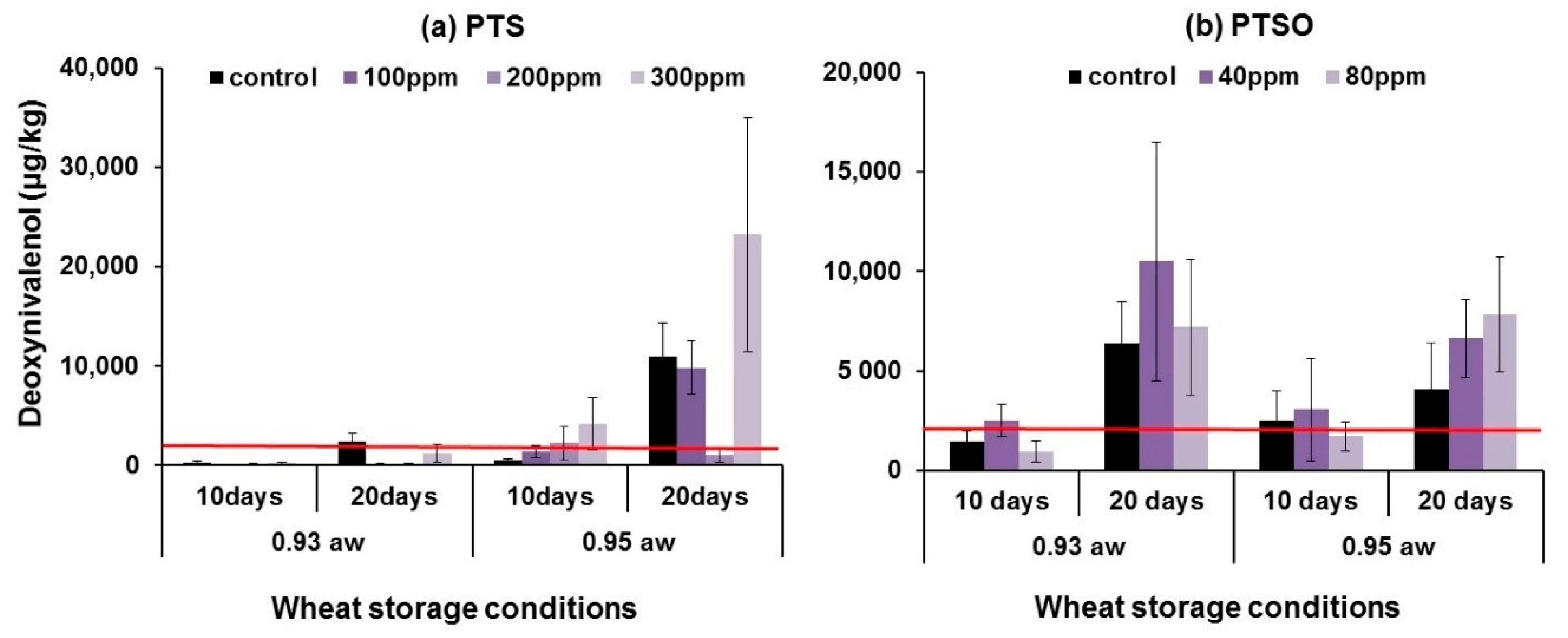
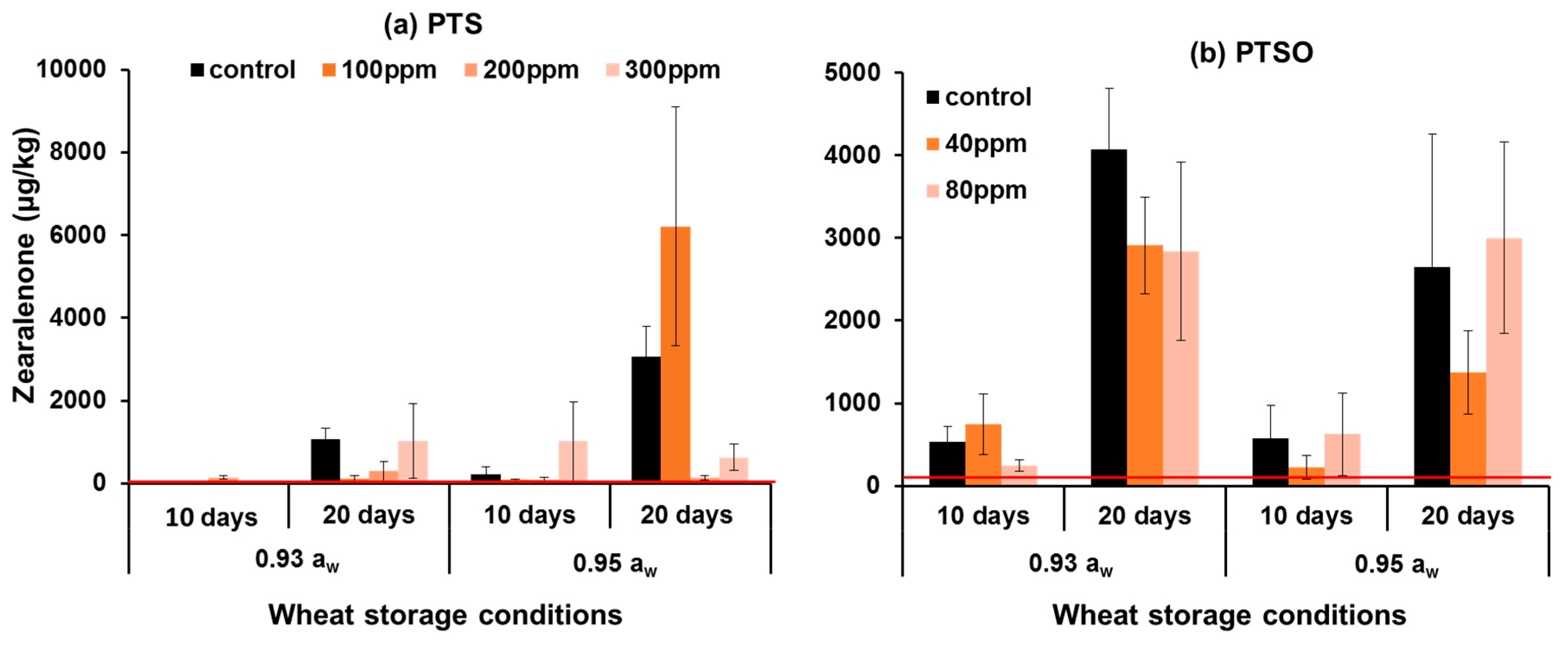
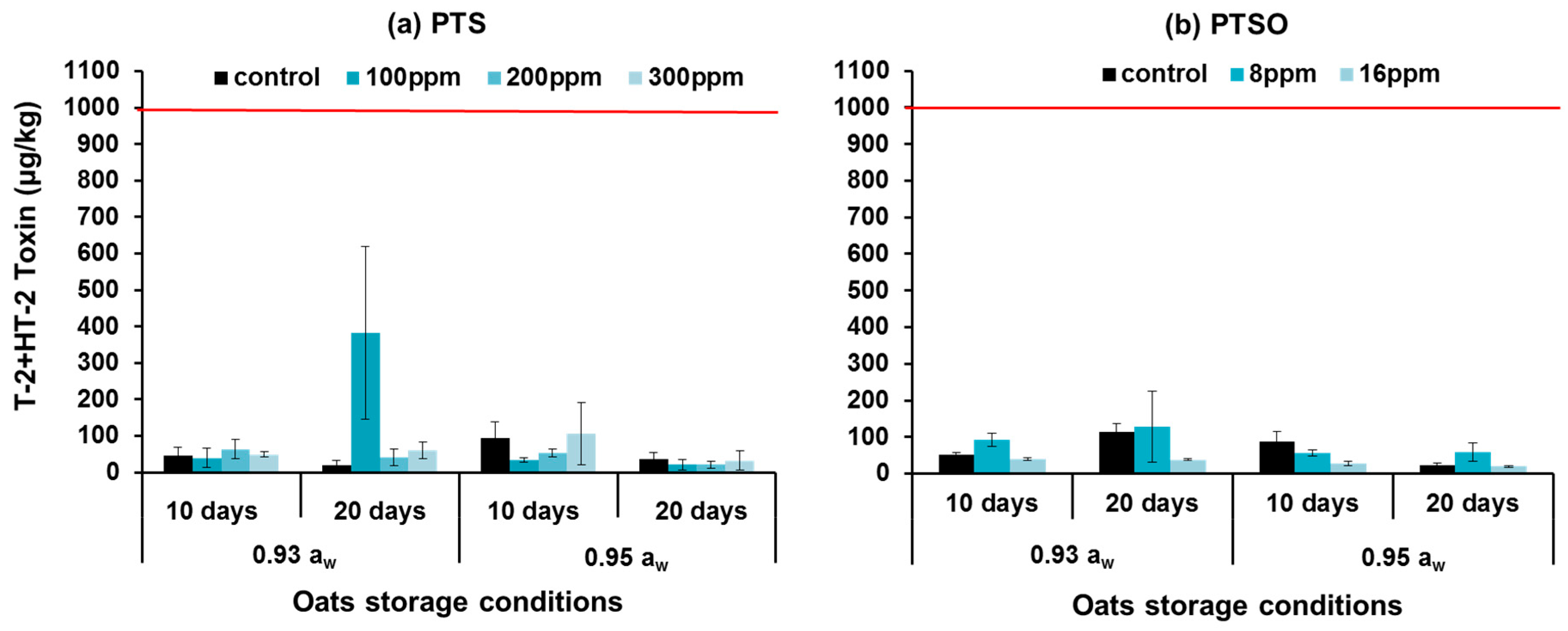
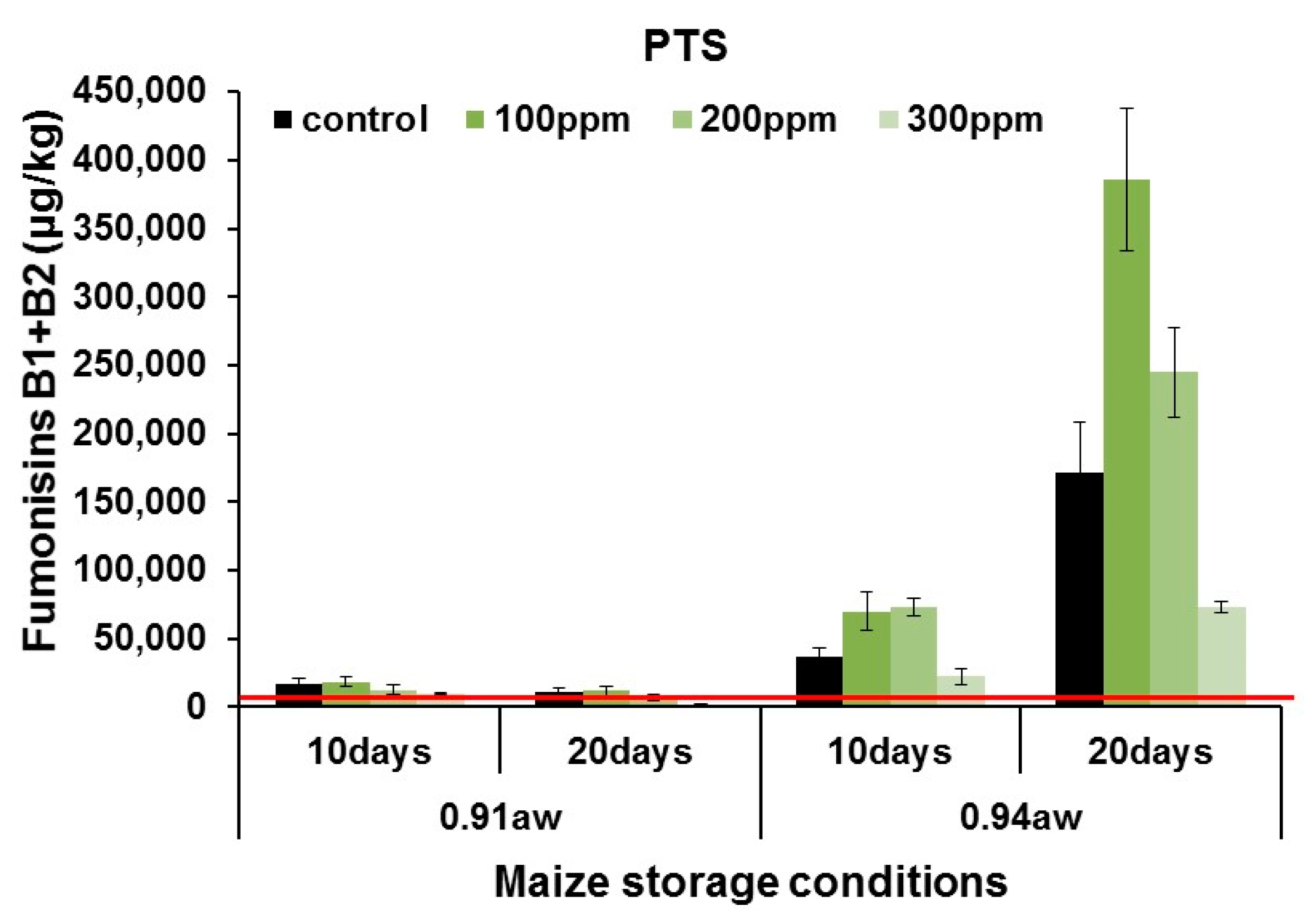
2.3. In Situ Mycotoxin Control in Stored Wheat, Oats and Maize Using PTS and PTSO
3. Discussion
3.1. In Vitro Efficacy of PTS and PTSO
3.2. In Situ Efficacy of PTS and PTSO against Mycotoxin Production in Stored Grain
4. Conclusions
5. Materials and Methods
5.1. Preparation of Stock Solutions of Chemical Compounds
5.2. In Vitro Studies: Fungal Species, Media, Inoculation and Measurements of Growth and Mycotoxin Production
5.3. In Situ Studies with Stored Cereal Grain
5.4. Mycotoxin Analyses
5.4.1. Equipment Description
5.4.2. In Vitro Extraction and Analysis
Deoxynivalenol
T-2 and HT-2
Fumonisins
5.4.3. In Situ Extraction and Analysis
5.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prakash, B.; Mishra, P.K.; Kedia, A.; Dwivedy, A.K.; Dubey, N.K. Efficacy of some essential oil components as food preservatives against food contaminating molds, aflatoxin b1 production and free radical generation. J. Food Qual. 2015, 38, 231–239. [Google Scholar] [CrossRef]
- Yin, M.; Tsao, S. Inhibitory effect of seven Allium plants upon three Aspergillus species. Int. J. Food Microbiol. 1999, 49, 49–56. [Google Scholar] [CrossRef]
- Yoshida, S.; Kasuga, S.; Hayashi, N.; Ushiroguchi, T.; Matsuura, H.; Nakagawa, S. Antifungal Activity of Ajoene Derived from Garlic. Appl. Environ. Microbiol. 1987, 53, 615–617. [Google Scholar]
- Benkeblia, N. Antimicrobial activity of essential oil extracts of various onions (Allium cepa) and garlic (Allium sativum). LWT - Food Sci. Technol. 2004, 37, 263–268. [Google Scholar] [CrossRef]
- Singh, I.; Singh, V.P. Effect of plant extracts on mycelial growth and aflatoxin production by Aspergillus flavus. Indian J. Microbiol. 2005, 45, 139–142. [Google Scholar]
- Aldred, D.; Cairns-Fuller, V.; Magan, N. Environmental factors affect efficacy of some essential oils and resveratrol to control growth and ochratoxin A production by Penicillium verrucosum and Aspergillus westerdijkiae on wheat grain. J. Stored Prod. Res. 2008, 44, 341–346. [Google Scholar] [CrossRef]
- Magan, N.; Aldred, D. Post-harvest control strategies: Minimizing mycotoxins in the food chain. Int. J. Food Microbiol. 2007, 119. [Google Scholar] [CrossRef]
- Garcia, D.; Garcia-Cela, E.; Ramos, A.J.; Sanchis, V.; Marín, S. Mould growth and mycotoxin production as affected by Equisetum arvense and Stevia rebaudiana extracts. Food Control 2011, 22, 1378–1384. [Google Scholar] [CrossRef]
- Garcia-Cela, E.; Gil-Serna, J.; Marin, S.; Acevedo, H.; Patino, B.; Ramos, A.J. Effect of preharvest anti-fungal compounds on Aspergillus steynii and A. carbonarius under fluctuating and extreme environmental conditions. Int. J. Food Microbiol. 2012, 159, 167–176. [Google Scholar] [CrossRef]
- Magan, N.; Hope, R.; Colleate, A.; Baxter, E. Relationship Between Growth and Mycotoxin Production by Fusarium species, Biocides and Environment. Eur. J. Plant Pathol. 2002, 108, 685–690. [Google Scholar] [CrossRef]
- Small, L.D.; Bailey, J.H.; Cavallito, C.T. Alkyl thiosulfinates. J. Am. Chem. Soc. 1947, 69, 1710–1713. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No. 1881/2006 of 19 December 2006, Setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 364, 5–24. [Google Scholar]
- European Commission. Commission Recommendation No 2013/165/EU of 27 March 2013 on the presence of T-2 and HT-2 toxin in cereals and cereal products. Off. J. Eur. Union 2013, 91, 12–15. [Google Scholar] [CrossRef]
- Aly, S.E.; Sabry, B.A.; Shaheen, M.S.; Hathout, A.S. Assessment of antimycotoxigenic and antioxidant activity of star anise (Illicium verum) in vitro. J. Saudi Soc. Agric. Sci. 2016, 15, 20–27. [Google Scholar] [CrossRef]
- Morcia, C.; Tumino, G.; Ghizzoni, R.; Bara, A.; Salhi, N.; Terzi, V.; Morcia, C.; Tumino, G.; Ghizzoni, R.; Bara, A.; et al. In vitro evaluation of sub-lethal concentrations of plant-derived antifungal compounds on FUSARIA growth and mycotoxin production. Molecules 2017, 22, 1271. [Google Scholar] [CrossRef]
- Soliman, K.; Badeaa, R. Effect of oil extracted from some medicinal plants on different mycotoxigenic fungi. Food Chem. Toxicol. 2002, 40, 1669–1675. [Google Scholar] [CrossRef]
- Yamamoto-Ribeiro, M.M.G.; Grespan, R.; Kohiyama, C.Y.; Ferreira, F.D.; Mossini, S.A.G.; Silva, E.L.; de Abreu Filho, B.A.; Mikcha, J.M.G.; Machinski Junior, M. Effect of Zingiber officinale essential oil on Fusarium verticillioides and fumonisin production. Food Chem. 2013, 141, 3147–3152. [Google Scholar] [CrossRef]
- Nguefack, J.; Leth, V.; Amvam Zollo, P.H.; Mathur, S.B. Evaluation of five essential oils from aromatic plants of Cameroon for controlling food spoilage and mycotoxin producing fungi. Int. J. Food Microbiol. 2004, 94, 329–334. [Google Scholar] [CrossRef]
- Xing, F.; Hua, H.; Selvaraj, J.N.; Zhao, Y.; Zhou, L.; Liu, X.; Liu, Y. Growth inhibition and morphological alterations of Fusarium verticillioides by cinnamon oil and cinnamaldehyde. Food Control 2014, 46, 343–350. [Google Scholar] [CrossRef]
- Sumalan, R.-M.; Alexa, E.; Poiana, M.-A. Assessment of inhibitory potential of essential oils on natural mycoflora and Fusarium mycotoxins production in wheat. Chem. Cent. J. 2013, 7, 32. [Google Scholar] [CrossRef]
- Venkatesh, H.N.; Sudharshana, T.N.; Abhishek, R.U.; Thippeswamy, S.; Manjunath, K.; Mohana, D.C. Antifungal and antimycotoxigenic properties of chemically characterised essential oil of Boswellia serrata Roxb. ex Colebr. Int. J. Food Prop. 2017, 1–13. [Google Scholar] [CrossRef]
- Ferreira, F.M.D.; Hirooka, E.Y.; Ferreira, F.D.; Silva, M.V.; Mossini, S.A.G.; Machinski, M., Jr. Effect of Zingiber officinale Roscoe essential oil in fungus control and deoxynivalenol production of Fusarium graminearum Schwabe in vitro. Food Addit. Contam. Part A 2018, 35, 2168–2174. [Google Scholar] [CrossRef]
- Elhouiti, F.; Tahri, D.; Takhi, D.; Ouinten, M.; Barreau, C.; Verdal-Bonnin, M.-N.; Bombarda, I.; Yousfi, M. Variability of composition and effects of essential oils from Rhanterium adpressum Coss. & Durieu against mycotoxinogenic Fusarium strains. Arch. Microbiol. 2017, 199, 1345–1356. [Google Scholar] [CrossRef]
- Kurita, N.; Koike, S. Synergistic antimicrobial effect of ethanol, sodium chloride, acetic acid and essential oil components. Agric. Biol. Chem. 1983, 47, 67–75. [Google Scholar] [CrossRef]
- Marin, S.; Magan, N.; Ramos, A.J.; Sanchis, V. Fumonisin-producing strains of Fusarium: A review of their ecophysiology. J. Food Prot. 2004, 67, 1792–1805. [Google Scholar] [CrossRef]
- Hope, R.; Aldred, D.; Magan, N. Comparison of the effect of environmental factors on deoxynivalenol production by F. culmorum and F. graminearum on wheat grain. Lett. Appl. Microbiol. 2005, 40, 295–300. [Google Scholar] [CrossRef]
- Medina, A.; Magan, N. Water availability and temperature affects production of T-2 and HT-2 by Fusarium langsethiae strains from north European countries. Food Microbiol. 2011, 28, 392–398. [Google Scholar] [CrossRef]
- Tracz, B.L.; Bordin, K.; de Melo Nazareth, T.; Costa, L.B.; de Macedo, R.E.F.; Meca, G.; Luciano, F.B. dAssessment of allyl isothiocyanate as a fumigant to avoid mycotoxin production during corn storage. LWT 2017, 75, 692–696. [Google Scholar] [CrossRef]
- Giorni, P.; Bertuzzi, T.; Battilani, P. Impact of fungi co-occurrence on mycotoxin contamination in maize during the growing season. Front. Microbiol. 2019, 10, 1265. [Google Scholar] [CrossRef]
- Magan, N.; Aldred, D.; Hope, R.; Mitchell, D. Environmental factors and interactions with mycoflora of grain and grapes: effects on growth and deoxynivalenol and ochratoxin production by Fusarium culmorum and Aspergillus carbonarius. Toxins 2010, 2, 353–366. [Google Scholar] [CrossRef]
- Mylona, K.; Sulyok, M.; Magan, N. Fusarium graminearum and Fusarium verticillioides colonisation of wheat and maize, environmental factors, dry matter losses and mycotoxin production relevant to the EU legislative limits. Food Addit. Contam. 2012, 29, 1118–1128. [Google Scholar] [CrossRef]
- Medina, A.; Valle-Algarra, F.M.; Jiménez, M.; Magan, N. Different sample treatment approaches for the analysis of T-2 and HT-2 toxins from oats-based media. J. Chromatogr. B 2010, 878, 2145–2149. [Google Scholar] [CrossRef] [Green Version]
- Sanchis, V.; Magan, N. Mycotoxins in food: Detection and control. In Environmental Profiles for Growth and Mycotoxin Production; Magan, N., Olsen, M., Eds.; Woodhead Publishing: Cambridge, UK, 2004; pp. 174–189. [Google Scholar]
- Krska, R.; Welzig, E.; Boudra, H. Analysis of Fusarium toxins in feed. Anim. Feed Sci. Technol. 2007, 137, 241–264. [Google Scholar] [CrossRef]
- Vishwanath, V.; Sulyok, M.; Labuda, R.; Bicker, W.; Krska, R. Simultaneous determination of 186 fungal and bacterial metabolites in indoor matrices by liquid chromatography/tandem mass spectrometry. Anal. Bioanal. Chem. 2009, 395, 1355–1372. [Google Scholar] [CrossRef]
- Sulyok, M.; Berthiller, F.; Krska, R.; Schuhmacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment (ppm) | PTS | PTSO | ||
|---|---|---|---|---|
| Species | ED50 | ED90 | ED50 | ED90 |
| F. graminearum | 33 | 144 | 52 | 186 |
| F. langsethiae | 12 | 80 | 21 | 113 |
| F. verticilloides0 | 64 | 172 | >200 | >200 |
| Treatments | F. graminearum | F. verticillioides | F. langsethiae |
|---|---|---|---|
| PTSO (0.98 aw) | 42 | 189 | 9.5 |
| PTSO (0.94 aw) | 36 | >100 | 50 |
| PTSO (0.92 aw) | 37 | >100 | 32 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mylona, K.; Garcia-Cela, E.; Sulyok, M.; Medina, A.; Magan, N. Influence of Two Garlic-Derived Compounds, Propyl Propane Thiosulfonate (PTS) and Propyl Propane Thiosulfinate (PTSO), on Growth and Mycotoxin Production by Fusarium Species In Vitro and in Stored Cereals. Toxins 2019, 11, 495. https://doi.org/10.3390/toxins11090495
Mylona K, Garcia-Cela E, Sulyok M, Medina A, Magan N. Influence of Two Garlic-Derived Compounds, Propyl Propane Thiosulfonate (PTS) and Propyl Propane Thiosulfinate (PTSO), on Growth and Mycotoxin Production by Fusarium Species In Vitro and in Stored Cereals. Toxins. 2019; 11(9):495. https://doi.org/10.3390/toxins11090495
Chicago/Turabian StyleMylona, Kalliopi, Esther Garcia-Cela, Michael Sulyok, Angel Medina, and Naresh Magan. 2019. "Influence of Two Garlic-Derived Compounds, Propyl Propane Thiosulfonate (PTS) and Propyl Propane Thiosulfinate (PTSO), on Growth and Mycotoxin Production by Fusarium Species In Vitro and in Stored Cereals" Toxins 11, no. 9: 495. https://doi.org/10.3390/toxins11090495
APA StyleMylona, K., Garcia-Cela, E., Sulyok, M., Medina, A., & Magan, N. (2019). Influence of Two Garlic-Derived Compounds, Propyl Propane Thiosulfonate (PTS) and Propyl Propane Thiosulfinate (PTSO), on Growth and Mycotoxin Production by Fusarium Species In Vitro and in Stored Cereals. Toxins, 11(9), 495. https://doi.org/10.3390/toxins11090495