Phytotoxic Responses of Soybean (Glycine max L.) to Botryodiplodin, a Toxin Produced by the Charcoal Rot Disease Fungus, Macrophomina phaseolina

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
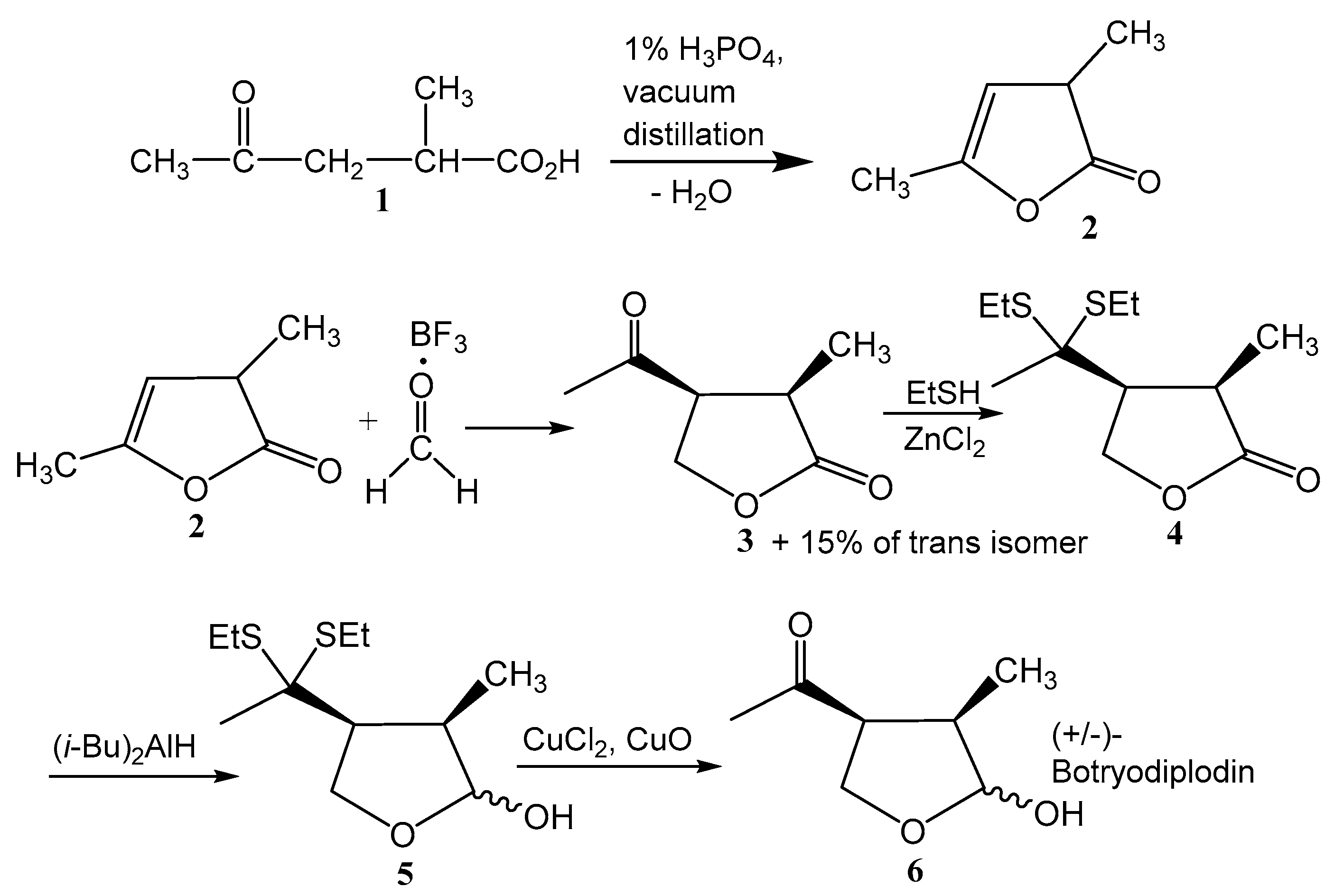
2.1. Synthesis of (±)-Botryodiplodin
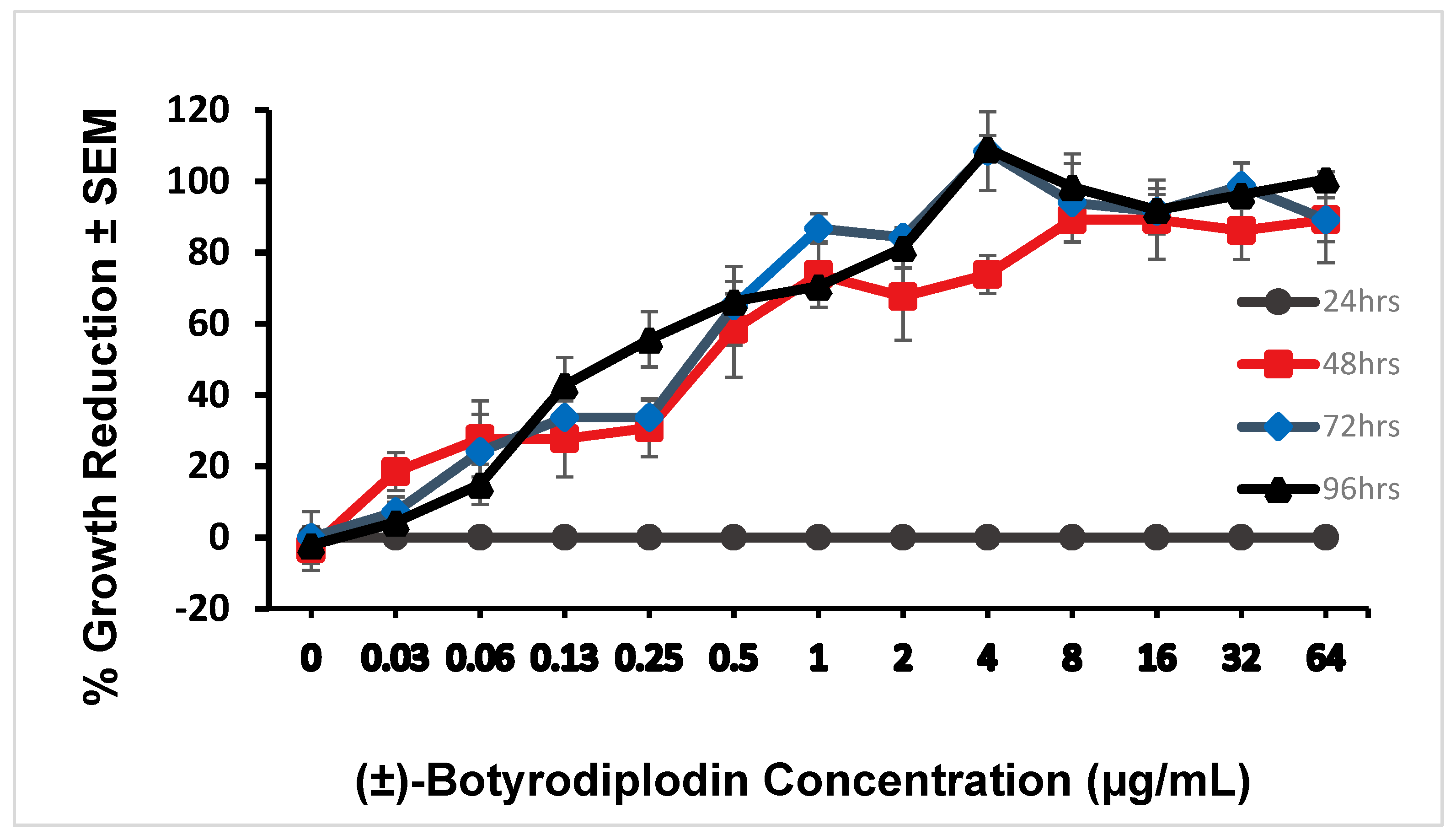
2.2. Phytotoxicity of (±)-Botryodiplodin in Lemna Pausicostata (Duckweed) Cultures
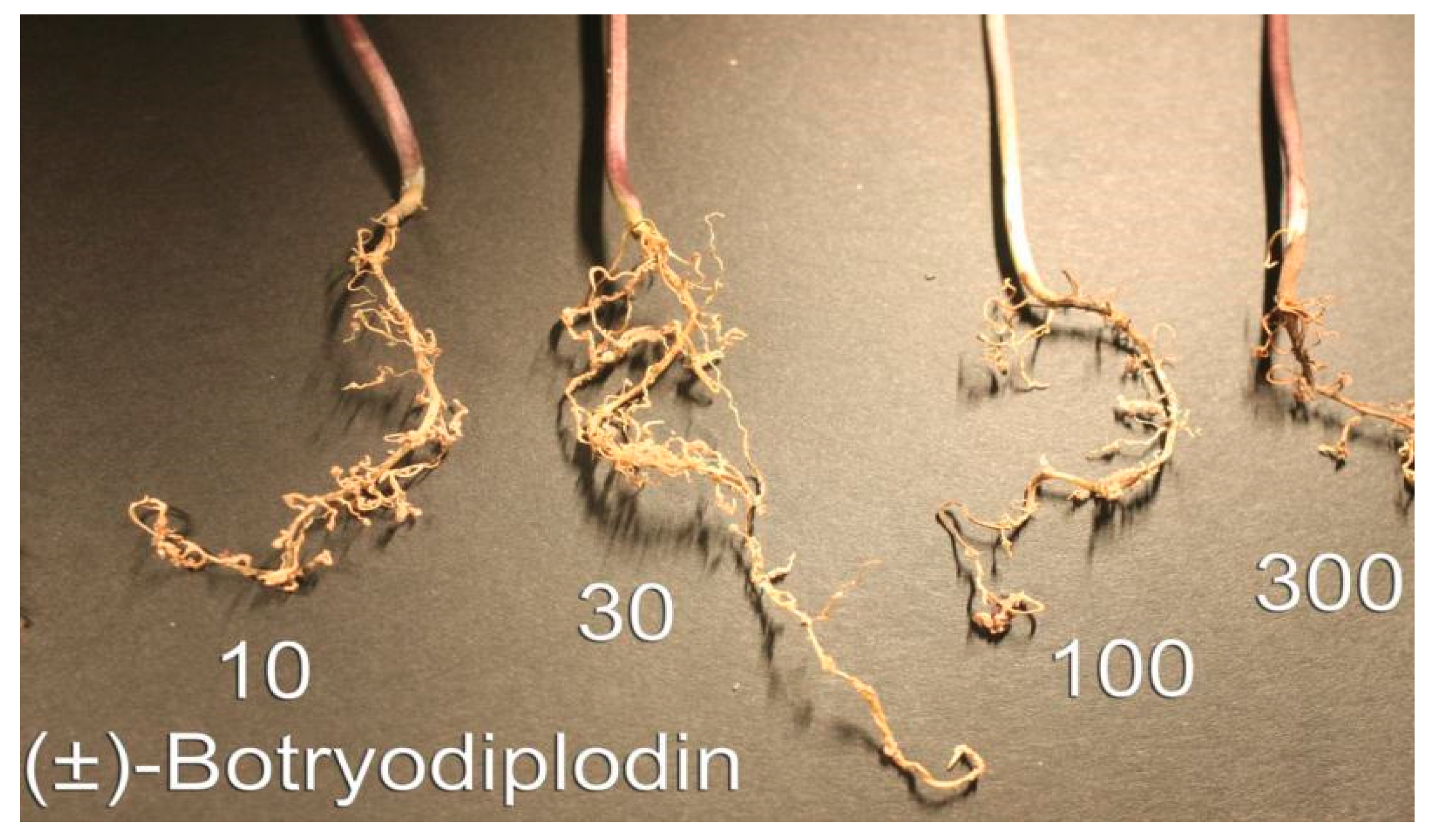
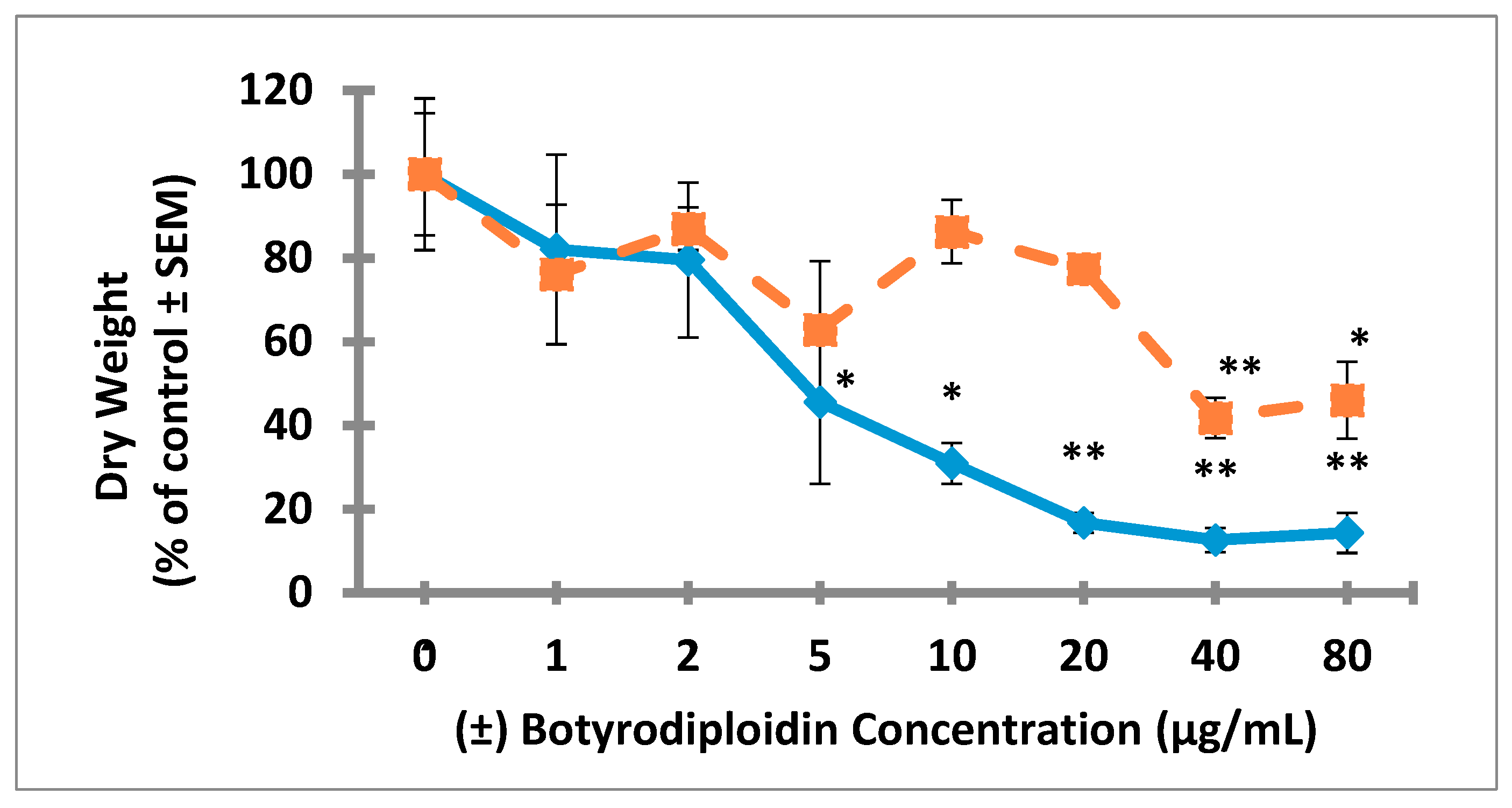
2.3. Root Toxicity of (±)-Botryodiplodin in Soybean Seedlings
3. Conclusions
4. Materials and Methods
4.1. Preparation of (±)-Botryodiplodin
4.2. Assay of Antibacterial Activity of Botryodiplodin
4.3. Plant Growth and Environmental Conditions
4.4. Phytotoxicity of (±)-Botryodiplodin in Soybean Leaf Discs
4.5. Phytotoxicity of (±)-Botryodiplodin in Duckweed Plant Cultures
4.6. Hydroponic Culture of Soybean Seedlings
4.7. Root Toxicity of (±)-Botryodiplodin in Soybean Seedlings in Hydroponic Culture
4.8. Light Micrographs of Soybean Seedling Roots Exposed to (±)-Botryodiplodin in Hydroponic Culture
4.9. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holiday, P.; Punithalingam, E. Macrophomina phaseolina. No. 275 in CMI (Commonweath Mycological Institute), Descriptions of Pathogenic Fungi and Bacteria; Commonweath Mycological Institute: Kew Surrey, UK, 1970. [Google Scholar]
- Ray, D.T.; Coffelt, T.A.; Dierig, D.A. Breeding guayule for commercial production. Ind. Crop. Prod. 2005, 22, 15–25. [Google Scholar] [CrossRef]
- Kuti, J.O.; Schading, R.L.; Latigo, G.V.; Bradford, J.M. Differential responses of guayule (Parthenium argentatum) genotypes to culture filtrate and toxin from Macrophomina Phaseolina. J. Phytopath. 1997, 145, 305–311. [Google Scholar] [CrossRef]
- Wyllie, T.D. Macrophomina phaseolina Charcoal rot. In World Soybean Research, Proceedings of the World Soybean Research Conference; Hill, L.D., Ed.; Interstate Printers and Publishers Inc.: Danville, IL, USA, 1976; pp. 482–484. [Google Scholar]
- Medhaug, I.; Stolpe, M.B.; Fischer, E.M.; Knutti, R. Reconciling controversies about the ‘global warming hiatus’. Nature 2017, 545, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Dubey, M.K.; Aamir, M.; Kaushik, M.S.; Khare, S.; Meena, M.; Singh, S.; Upadhyay, R.S. PR toxin-biosynthesis, genetic regulation, toxicological potential, prevention and control measures, Overview and challenges. Front. Pharmacol. 2018, 9, 288. [Google Scholar] [CrossRef] [Green Version]
- Moreau, S.; Lablache-Combier, A.; Biguet, J.; Foulon, C.; Delfosse, C. Botryodiploidin, a mycotoxin produced by a strain of Penicillium roqueforti. J. Org. Chem. 1982, 47, 2358–2359. [Google Scholar] [CrossRef]
- Nielsen, K.F.; Sumarah, M.W.; Frisvad, J.C.; Miller, J.D. Production of metabolites from the Penicillium roqueforti complex. J. Agric. Food Chem. 2006, 54, 3756–3763. [Google Scholar] [CrossRef]
- O’Brien, M.; Nielsen, K.F.; O’Kiely, P.; Forristal, P.D.; Fuller, H.T.; Frisvad, J.C. Mycotoxins and other secondary metabolites produced in vitro by Penicillium paneum Frisvad and Penicillium roqueforti Thom isolated from baled grass silage in Ireland. J. Agric. Food Chem. 2006, 54, 9268–9276. [Google Scholar] [CrossRef] [Green Version]
- Shier, W.T.; Abbas, H.K.; Baird, R.E.; Ramezani, M.; Sciumbato, G.L. (-)-Botryodiplodin, a unique ribose-analog toxin. Toxin Rev. 2007, 26, 343–386. [Google Scholar] [CrossRef]
- Wrather, J.A.; Anderson, T.R.; Arsyad, D.M.; Tan, Y.; Ploper, L.D.; Porta-Puglia, A.; Ram, H.H.; Yorinori, J.T. Soybean disease loss estimates for the top ten soybean-producing countries in 1998. Can. J. Plant. Pathol. 2001, 23, 115–121. [Google Scholar] [CrossRef]
- Wrather, J.A.; Koenning, S.R.; Anderson, T.R. Effect of diseases on soybean yields in the United States and Ontario (1999–2002). Plant. Health Prog. 2003, 4, 24. [Google Scholar] [CrossRef] [Green Version]
- Wrather, J.A.; Koenning, S.R. Estimates of disease effects on soybean yields in the United States 2003–2005. J. Nematol. 2006, 38, 173–180. [Google Scholar] [PubMed]
- Wrather, J.A.; Stienstra, W.C.; Koenning, S.R. Soybean disease loss estimates for the United States from 1996 to 1998. Can. J. Plant. Pathol. 2001, 23, 122–131. [Google Scholar] [CrossRef]
- Wrather, J.A. Soybean disease loss estimates for the southern United States, 1974 to 1994. Plant. Dis. 1995, 79, 1076–1079. [Google Scholar]
- Mengistu, A.; Wrather, A.; Rupe, J.C. Charcoal Rot. Compendium of Soybean Diseases and Pests, 5th ed.; Hartman, G.L., Rupe, J.C., Sikora, E.J., Domier, L.L., Davis, J.A., Steffey, K.L., Eds.; American Phytopathology Society: Saint Paul, MN, USA, 2015; pp. 67–69. [Google Scholar]
- Mengistu, A.; Reddy, K.N.; Zablotowicz, R.M.; Wrather, A.J. Propagule densities of Macrophomina phaseolina in soybean tissue and soil as affected by tillage, cover crop, and herbicides. Plant. Health Prog. 2009, 10, 28. Available online: https://www.plantmanagementnetwork.org/pub/php/research/2009/soybean/ (accessed on 23 December 2019). [CrossRef] [Green Version]
- Paris, R.L.; Mengistu, A.; Tyler, J.M.; Smith, J.R. Registration of soybean germplasm line DT97-4290 with moderate resistance to charcoal rot. Crop. Sci. 2006, 46, 2324–2325. [Google Scholar] [CrossRef] [Green Version]
- Bellaloui, N.; Mengistu, A.; Zobiole, L.H.S.; Shier, W.T. Resistance to toxin-mediated fungal infection, Role of lignins, isoflavones, other seed phenolics, sugars and boron in the mechanism of resistance to charcoal rot disease in soybean. Toxin Rev. 2012, 31, 16–26. [Google Scholar] [CrossRef]
- Bellaloui, N.; Mengistu, A.; Paris, R.L. Soybean seed composition in cultivars differing in resistance to charcoal rot (Macrophomina phaseolina). J. Agric. Sci. 2008, 146, 667–675. [Google Scholar] [CrossRef] [Green Version]
- Gangopadhyay, S.; Wyllie, T.D.; Luedders, V.D. Charcoal rot disease of soybean transmitted by seeds. Plant. Dis. Rep. 1970, 54, 1088–1091. [Google Scholar]
- Ichinose, Y.; Taguchi, F.; Mukaihara, T. Pathogenicity and virulence factors of Pseudomonas syringae. J. Gen. Plant. Pathol. 2013, 79, 285–296. [Google Scholar] [CrossRef]
- Mihail, J.D. Macrophomina. Methods for Research on Soilborne Phytopathogenic Fungi; Singleton, L., Mihail, J.D., Rush, C.M., Eds.; The American Phytopathological Society, APS Press: Paul, MN, USA, 1992; pp. 134–136. [Google Scholar]
- Oku, H. Plant. Pathogenesis and Disease Control; Lewis Publishers: Boca Raton, FL, USA, 1994. [Google Scholar]
- Siddiqui, K.A.I.; Gupta, A.K.; Paul, A.K.; Banerjee, A.K. Purification and properties of a heat-resistant exotoxin produced by Macrophomina phaseolina (Tassi) Gold in culture. Experentia 1979, 35, 1222–1223. [Google Scholar] [CrossRef]
- Sen Gupta, R.; Chandran, R.R.; Divekar, P.V. Botryodiplodin, a new antibiotic from Botryodiplodia theobromae. I. Production, isolation, and biological properties. Indian J. Exp. Biol. 1966, 4, 152–153. [Google Scholar]
- Islam, M.S.; Haque, M.S.; Islam, M.M.; Emdad, E.M.; Halim, H.; Hossen, Q.M.M.; Hossain, M.Z.; Ahmed, B.; Rahim, S.; Rahman, M.S.; et al. Tools to kill, Genome of one of the most destructive plant pathogenic fungi Macrophomina phaseolina. BMC Genom. 2012, 13, 493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhar, T.K.; Siddiqui, K.A.I.; Ali, E. Structure of phaseolinone, a novel phytotoxin from Macrophomina phaseolina. Tetrahedron Lett. 1982, 23, 5459–5462. [Google Scholar]
- Bunkers, G.; Kenfield, D.; Strobel, G.; Sugawara, F. Structure-activity relationships of the eremophilanes produced by Drechslera gigantea. Phytochem 1990, 29, 1471–1474. [Google Scholar] [CrossRef]
- Kitahara, T.; Kiyota, H.; Kurata, H.; Mori, K. Synthesis of oxygenated eremophilanes, gigantenone, phomenone and phaseolinone, phytotoxins from pathogenic fungi. Tetrahedron 1911, 47, 1649–1654. [Google Scholar] [CrossRef]
- Sugawara, F.; Hallock, Y.F.; Bunkers, G.D.; Kenfield, D.S.; Strobel, G.; Yoshida, S. Phytoactive eremophilanes produced by the weed pathogen Drechslera gigantea. Biosci. Biotechnol. Biochem. 1993, 57, 236–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramezani, M.; Shier, W.T.; Abbas, H.K.; Tonos, J.L.; Baird, R.E.; Sciumbato, G.L. Soybean charcoal rot disease fungus Macrophomina phaseolina in Mississippi produces the phytotoxin, (-)-botryodiplodin, but no detectable phaseolinone. J. Nat. Prod. 2007, 70, 128–129. [Google Scholar] [CrossRef]
- Abbas, H.K.; Bellaloui, N.; Accinelli, C.; Smith, J.R.; Shier, W.T. Toxin production in soybean (Glycine max L.) plants with charcoal rot disease and by Macrophomina phaseolina, the fungus that causes the disease. Toxins 2019, 11, 645. [Google Scholar] [CrossRef] [Green Version]
- Goos, R.D.; Cox, E.A.; Stotzky, G. Botryodiplodia theobromae and its association with Musa species. Mycologia 1961, 53, 262–277. [Google Scholar] [CrossRef]
- McCurry, P.M.; Abe, K. Stereochemistry and synthesis of the antileukemic agent botryodiplodin. J. Am. Chem. Soc. 1973, 95, 5824–5825. [Google Scholar] [CrossRef]
- Moule, Y.; Decloitre, F.; Hamon, G. Mutagenicity of the mycotoxin botryodiplodin in the Salmonella typhimurium/microsomal activation test. Environ. Mutagen. 1981, 3, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Moule, Y.; Douce, C.; Moreau, S.; Darracq, N. Effects of the mycotoxin botryodiplodin on mammalian cells in culture. Chem.-Biol. Interact. 1981, 37, 155–164. [Google Scholar] [CrossRef]
- Moule, Y.; Renauld, F.; Darracq, N.; Douce, C. DNA-protein cross-linking by the mycotoxin, botryodiplodin, in mammalian cells. Carcinogen 1982, 3, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Rehnberg, N.; Magnusson, G. Total synthesis of (-)- and (+)-botryodiplodin and (+)- and (-)-epibotryodiplodin. Acta Chem. Scand. 1990, 44, 377–383. [Google Scholar] [CrossRef]
- Fujimoto, Y.; Kamiya, M.; Tsunoda, H.; Ohtsubo, K.; Tatsuno, T. Recherche toxicologique des substances métaboliques de Penicillium carneo-lutescens. Chem. Pharm. Bull. 1980, 28, 1062–1066. [Google Scholar] [CrossRef]
- Félix, C.; Salvatore, M.M.; DellaGreca, M.; Ferreira, V.; Duarte, A.S.; Salvatore, F.; Naviglio, D.; Gallo, M.; Alves, A.; Esteves, A.C.; et al. Secondary metabolites produced by grapevine strains of Lasiodiplodia theobromae grown at two different temperatures. Mycologia 2019, 111, 466–476. [Google Scholar] [CrossRef]
- Bhatnagar, D.; Payne, G.A.; Cleveland, T.E.; Robens, J.F. Mycotoxins, Current Issues in USA. In Meeting the Mycotoxin Menace; Barug, H., van Egmond, H.P., Lopez-Garcia, R., van Osenbruggen, W.A., Visconti, A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2004; pp. 17–47. [Google Scholar]
- Horn, B.W.; Dorner, J.W. Regional differences in production of aflatoxin B1 and cyclopiazonic acid by soil isolates of Aspergillus flavus along a transect within the United States. Appl. Environ. Microbiol. 1999, 65, 1444–1449. [Google Scholar] [CrossRef] [Green Version]
- Shier, W.T. On the origin of antibiotics and mycotoxins. Toxin Rev. 2011, 30, 6–30. [Google Scholar] [CrossRef]
- Ryder, L.S.; Talbot, N.J. Regulation of appressorium development in pathogenic fungi. Curr. Opin. Plant. Biol. 2015, 26, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Dunlap, J.R.; Bruton, D.B. Pigment biosynthesis by Macrophomina phaseolina. The glycine-specific requirement. Trans. Br. Mycol. Soc. 1986, 86, 111–115. [Google Scholar] [CrossRef]
- Renault, F.; Moreau, S.; Lablache-Combier, A.; Tiffon, B. Botryodiplodin: A mycotoxin from Penicillium roqueforti, Reaction with amino-pyrimidines, amino-purines and 2’-deoxynucleosides. Tetrahedron 1985, 41, 955–962. [Google Scholar] [CrossRef]
- Alam, S.; Abbas, H.K.; Okunowu, W.O.; Kotowicz, J.; Butler, A.M.; Shier, W.T. Development of an in-culture assay for detecting production of the mycotoxin (-)-botryodiplodin by Macrophomina phaseolina. In preparation.
- Brechenmacher, L.; Lee, J.; Sachdev, S.; Song, Z.; Nguyen, T.H.N.; Joshi, T.; Oehrle, N.; Libault, M.; Mooney, B.; Xu, D.; et al. Establishment of a protein reference map for soybean root hair cells. Plant. Physiol. 2009, 149, 670–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, D.R.; Negi, S.; Sukumar, P.; Muday, G.K. Ethylene inhibits lateral root development, increases IAA transport and expression of PIN3 and PIN7 auxin efflux carriers. Development 2011, 138, 3485–3495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negi, S.; Ivanchenko, M.G.; Muday, G.K. Ethylene regulates lateral root formation and auxin transport in Arabidopsis thaliana. Plant. J. 2008, 55, 175–187. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.; Chen, M.; Xu, Z.; Li, L.; Chen, X.; Ma, Y. Characterization of ethylene receptors and their interactions with GmTPR-A novel tetratricopeptide repeat protein (TPR) in soybean (Glycine max L.). J. Integr. Agric. 2013, 12, 571–581. [Google Scholar] [CrossRef]
- Drew, M.C. Soil aeration and plant root metabolism. Soil Sci. 1992, 154, 259–268. [Google Scholar] [CrossRef]
- Jackson, M.B.; Drew, M.C. The effect of flooding on growth and metabolism of herbaceous plants. In Physiological Ecology, A Series of Monographs, Texts and Treatises; Kozlowski, T.T., Ed.; Academic Press: Cambridge, MA, USA, 1984; pp. 47–128. [Google Scholar]
- Roy, R.; Bhattacharya, G.; Siddiqui, K.A.I.; Bhadra, R. A new antileishmanial compound, phaseolinone. BioChem. Biophys. Res. Commun. 1990, 168, 43–50. [Google Scholar] [CrossRef]
- Sturdikova, M.; Fuskova, A.; Proksa, B.; Fuska, J. Relationships between structure and biological activities of the antibiotic PSX-1 (botryodiplodine). Biology (Bratisl. Slovak.) 1988, 43, 233–238. [Google Scholar]
- Fuska, J.; Fuskova, A. The in vitro-in vivo effect of antibiotic PSX-1 on lympholeukemia L-5178. J. Antibiot. (Tokyo) 1976, 29, 981–982. [Google Scholar] [CrossRef]
- Chakraborty, S.; Lala, S. Assessment of the antifertility effect of phaseolinone, an antileishmanial agent, in male rats. Contraception 1998, 58, 183–191. [Google Scholar] [CrossRef]
- Fuska, J.; Kuhr, I.; Nemec, P.; Fuskova, A. Antitumor antibiotics produced by Penicillium stipitatum. J. Antibiot (Tokyo) 1974, 27, 123–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Helberger, J.H.; Ulubay, S.; Civelekoglu, H. EIN EINFACHES VERFAHREN ZUR GEWINNUNG VON ALPHA-ANGELICALACTON UND UBER DIE HYDRIERENDE SPALTUNG SAUERSTOFFHALTIGER RINGE. ANNALEN DER CHEMIE-JUSTUS LIEBIG. 1949, 561, 215–220. (In German) [Google Scholar] [CrossRef]
- Mukaiyama, T.; Wada, M.; Hanna, J. A convenient synthesis of the antibiotic botryodiplodin. Chem. Lett. 1974, 3, 1181–1184. [Google Scholar] [CrossRef] [Green Version]
- Nouguier, R.; Gastaldi, S.; Stien, D.; Bertrand, M.; Villar, F.; Andrey, O.; Renaud, P. Synthesis of (±)- and (-)-botryodiplodin using stereoselective radical cyclizations of acyclic esters and acetals. Tetrahedron Asymmetry 2003, 14, 3005–3018. [Google Scholar] [CrossRef]
- Owen, P.A.; Nickell, C.D.; Noel, G.R.; Thomas, D.J.; Frey, K. Registration of ‘Saline’ Soybean. Crop. Sci. 1994, 34, 1689. [Google Scholar] [CrossRef]
- Tanaka, T.; Abbas, H.K.; Duke, S.O. Structure-dependent phytotoxicity of fumonisins and related compounds in a duckweed bioassay. Phytochem 1993, 33, 779–785. [Google Scholar] [CrossRef]
- Villagarcia, M.R.; Carter, T.E.; Rufty, T.W.; Niewoehner, A.S.; Jennette, M.W.; Arrellano, C. Genotypic rankings for aluminum tolerance of soybean roots grown in hydroponics and sand culture. Crop. Sci. 2001, 41, 1499–1507. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, D.; Hrapchak, B. Theory and Practice of Histotechnology, 2nd ed.; Battelle Press: Columbus, OH, USA, 1980; pp. 330–331. [Google Scholar]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbas, H.K.; Bellaloui, N.; Butler, A.M.; Nelson, J.L.; Abou-Karam, M.; Shier, W.T. Phytotoxic Responses of Soybean (Glycine max L.) to Botryodiplodin, a Toxin Produced by the Charcoal Rot Disease Fungus, Macrophomina phaseolina. Toxins 2020, 12, 25. https://doi.org/10.3390/toxins12010025
Abbas HK, Bellaloui N, Butler AM, Nelson JL, Abou-Karam M, Shier WT. Phytotoxic Responses of Soybean (Glycine max L.) to Botryodiplodin, a Toxin Produced by the Charcoal Rot Disease Fungus, Macrophomina phaseolina. Toxins. 2020; 12(1):25. https://doi.org/10.3390/toxins12010025
Chicago/Turabian StyleAbbas, Hamed K., Nacer Bellaloui, Alemah M. Butler, Justin L. Nelson, Mohamed Abou-Karam, and W. Thomas Shier. 2020. "Phytotoxic Responses of Soybean (Glycine max L.) to Botryodiplodin, a Toxin Produced by the Charcoal Rot Disease Fungus, Macrophomina phaseolina" Toxins 12, no. 1: 25. https://doi.org/10.3390/toxins12010025
APA StyleAbbas, H. K., Bellaloui, N., Butler, A. M., Nelson, J. L., Abou-Karam, M., & Shier, W. T. (2020). Phytotoxic Responses of Soybean (Glycine max L.) to Botryodiplodin, a Toxin Produced by the Charcoal Rot Disease Fungus, Macrophomina phaseolina. Toxins, 12(1), 25. https://doi.org/10.3390/toxins12010025