Neurotrophic Properties of C-Terminal Domain of the Heavy Chain of Tetanus Toxin on Motor Neuron Disease


Abstract
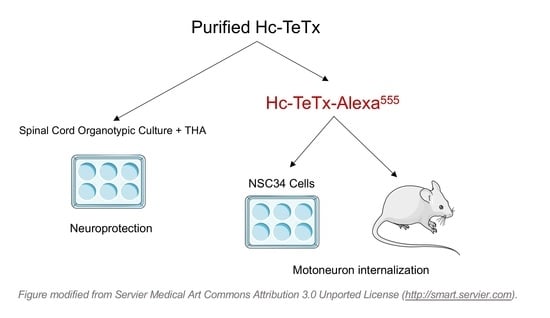
:
{kind=link}
{kind=link}
{kind=link}
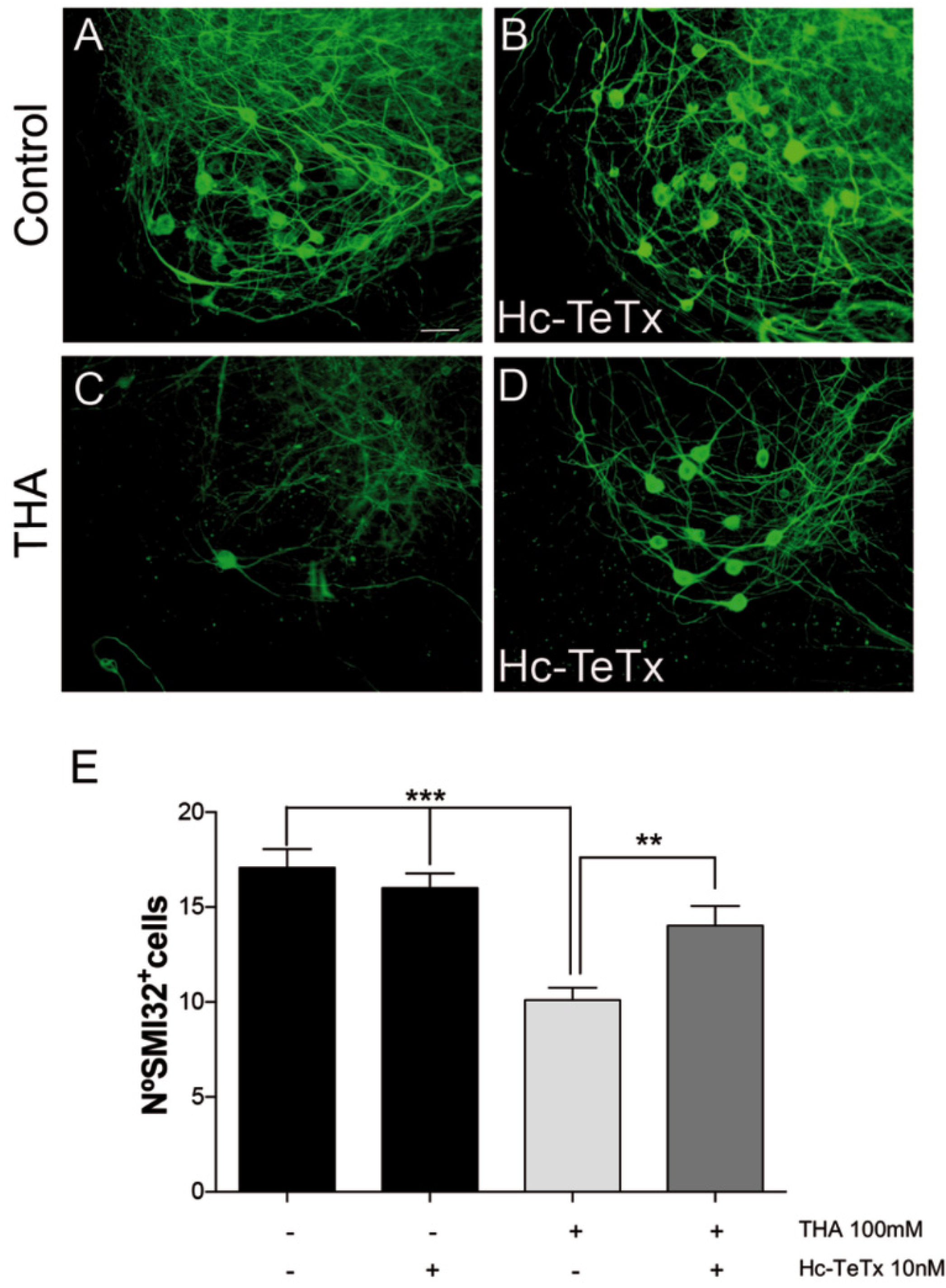{kind=link}
{kind=link}
1. Introduction
2. Results
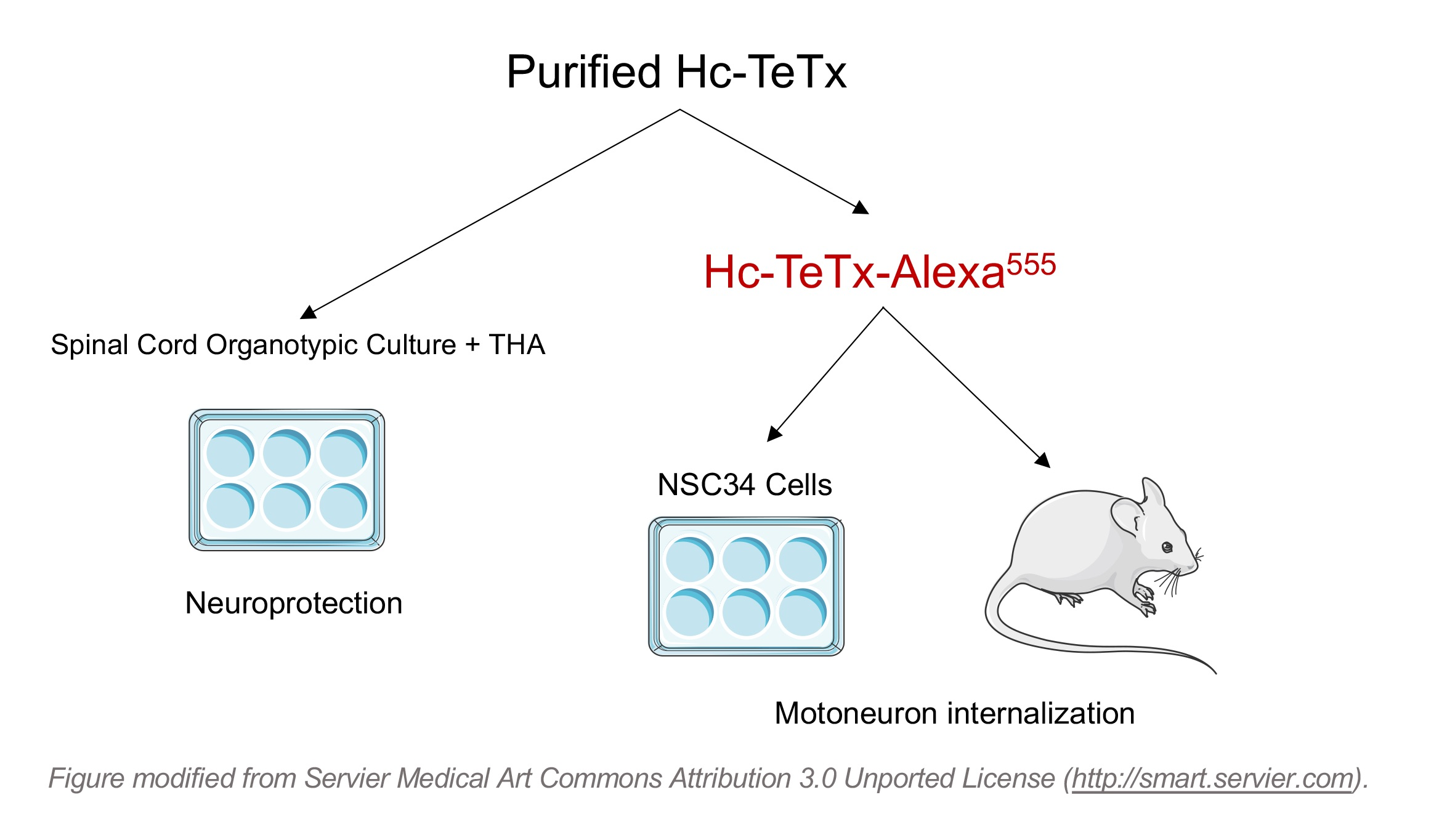
2.1. Purification and Fluorescence Labeling of Recombinant Protein Hc-TeTx
2.2. Membrane Motoneuron Binding and Retrograde Transport of Recombinant Hc-TeTx Protein
2.3. Motor Neuron Preservation under Chronic Excitotoxicity
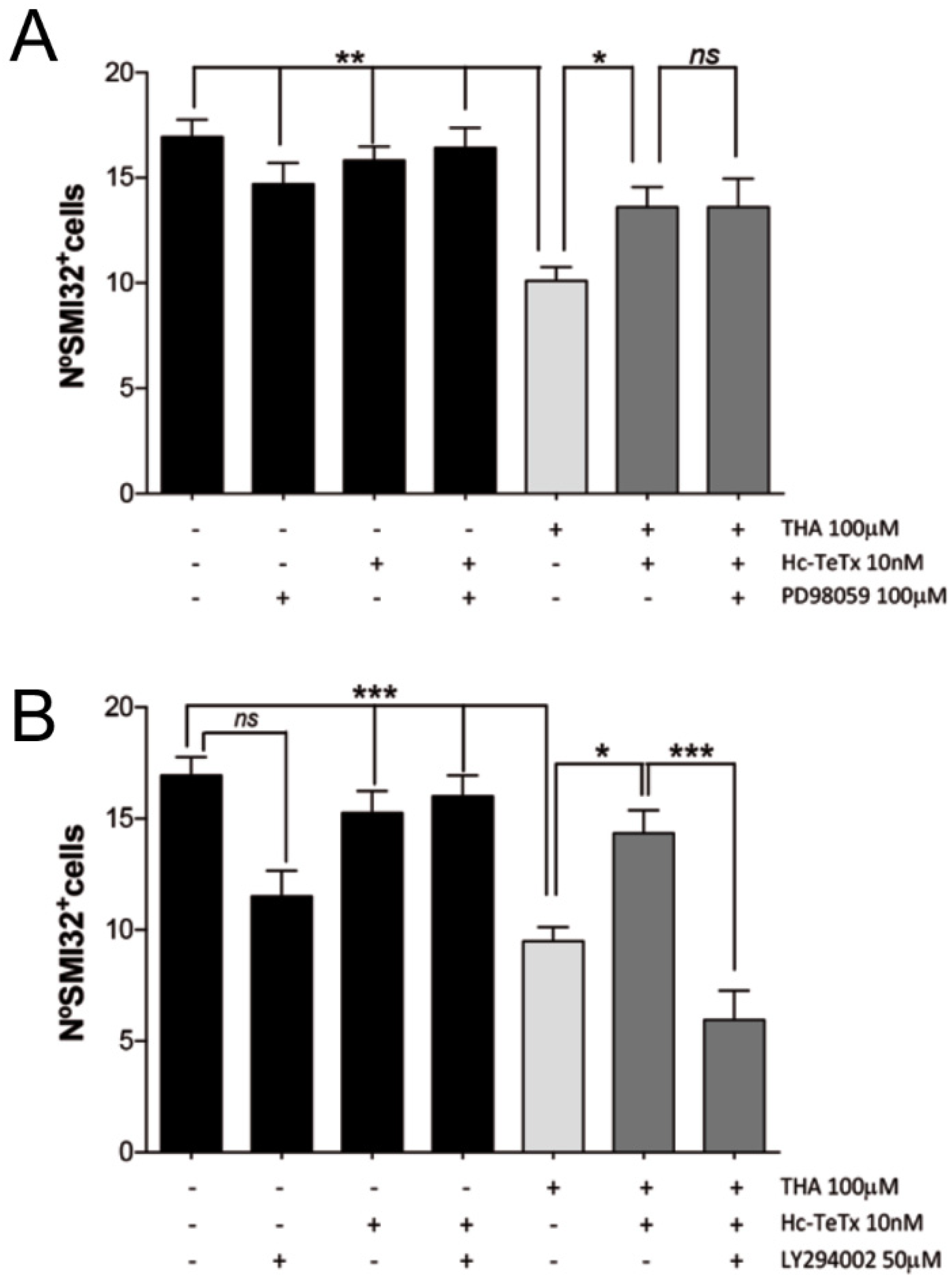
2.4. Signaling Cascade Induced by Hc-TeTx under Chronic Excitotoxicity
3. Discussion
4. Materials and Methods
4.1. Recombinant Hc-TeTx Protein Purification
4.2. Hc-TeTx Retrogade Transport
4.3. NSC-34 Cell Culture
4.4. Spinal Cord Organotypic Culture
4.5. Motor Neuron Counting
4.6. Immunofluorescence
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wijesekera, L.C.; Leigh, P.N. Amyotrophic lateral sclerosis. Orphanet J. Rare Dis. 2009, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludolph, A.C.; Jesse, S. Evidence-based drug treatment in amyotrophic lateral sclerosis and upcoming clinical trials. Ther. Adv. Neurol. Disord. 2009, 2, 319–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, K.; Aoki, M.; Tsuji, S.; Itoyama, Y.; Sobue, G.; Togo, M.; Hamada, C.; Tanaka, M.; Akimoto, M.; Nakamura, K.; et al. Safety and efficacy of edaravone in well defined patients with amyotrophic lateral sclerosis: A randomised, double-blind, placebo-controlled trial. Lancet Neurol. 2017, 16, 505–512. [Google Scholar] [CrossRef]
- Talbot, K.; Feneberg, E.; Scaber, J.; Thompson, A.G.; Turner, M.R. Amyotrophic lateral sclerosis: The complex path to precision medicine. J. Neurol. 2018, 265, 2454–2462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bizzini, B.; Stoeckel, K.; Schwab, M. An antigenic polypeptide fragment isolated from tetanus toxin: Chemical characterization, binding to gangliosides and retrograde axonal transport in various neuron systems. J. Neurochem. 1977, 28, 529–542. [Google Scholar] [CrossRef]
- Manning, K.A.; Erichsen, J.T.; Evinger, C. Retrograde transneuronal transport properties of fragment C of tetanus toxin. Neuroscience 1990, 34, 251–263. [Google Scholar] [CrossRef]
- Price, D.L.; Griffin, J.; Young, A.; Peck, K.; Stocks, A. Tetanus toxin: Direct evidence for retrograde intraaxonal transport. Science 1975, 30, 945–947. [Google Scholar] [CrossRef]
- Lalli, G.; Schiavo, G. Analysis of retrograde transport in motor neurons reveals common endocytic carriers for tetanus toxin and neurotrophin receptor p75NTR. J. Cell Biol. 2002, 156, 233–239. [Google Scholar] [CrossRef]
- Lalli, G.; Bohnert, S.; Deinhardt, K.; Verastegui, C.; Schiavo, G. The journey of tetanus and botulinum neurotoxins in neurons. Trends Microbiol. 2003, 11, 431–437. [Google Scholar] [CrossRef]
- Roux, S.; Colasante, C.; Saint Cloment, C.; Barbier, J.; Curie, T.; Girard, E.; Molgó, J.; Brûlet, P. Internalization of a GFP-tetanus toxin C-terminal fragment fusion protein at mature mouse neuromuscular junctions. Mol. Cell Neurosci. 2005, 30, 572–582. [Google Scholar] [CrossRef]
- Herrando-Grabulosa, M.; Casas, C.; Aguilera, J. The C-terminal domain of tetanus toxin protects motor neurons against acute excitotoxic damage on spinal cord organotypic cultures. J. Neurochem. 2013, 124, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Herreros, J.; Ng, T.; Schiavo, G. Lipid rafts act as specialized domains for tetanus toxin binding and internalization into neurons. Mol. Biol. Cell 2001, 12, 2947–2960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpern, J.L.; Loftus, A. Characterization of the receptor-binding domain of tetanus toxin. J. Biol. Chem. 1993, 268, 11188–11192. [Google Scholar] [PubMed]
- Benson, M.A.; Fu, Z.; Kim, J.J.; Baldwin, M.R. Unique ganglioside recognition strategies for clostridial neurotoxins. J. Biol. Chem. 2011, 286, 34015–34022. [Google Scholar] [CrossRef] [Green Version]
- Yavin, E.; Nathan, A. Tetanus toxin receptors on nerve cells contain a trypsin-sensitive component. Eur. J. Biochem. 1986, 154, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Herreros, J.; Lalli, G.; Montecucco, C.; Schiavo, G. Tetanus toxin fragment C binds to a protein present in neuronal cell lines and motor neurons. J. Neurochem. 2000, 74, 1941–1950. [Google Scholar] [CrossRef]
- Deinhardt, K.; Salinas, S.; Verastegui, C.; Watson, R.; Worth, D.; Hanrahan, S.; Bucci, C.; Schiavo, G. Rab5 and Rab7 control endocytic sorting along the axonal retrograde transport pathway. Neuron 2006, 52, 293–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaïb-Oukadour, I.; Gil, C.; Aguilera, J. The C-terminal domain of the heavy chain of tetanus toxin rescues cerebellar granule neurones from apoptotic death: Involvement of phosphatidylinositol 3-kinase and mitogen-activated protein kinase pathways. J. Neurochem. 2004, 90, 1227–1236. [Google Scholar] [CrossRef]
- Chaïb-Oukadour, I.; Gil, C.; Rodríguez-Alvarez, J.; Ortega, A.; Aguilera, J. Tetanus toxin H(C) fragment reduces neuronal MPP+ toxicity. Mol. Cell Neurosci. 2009, 41, 297–303. [Google Scholar] [CrossRef]
- Mendieta, L.; Venegas, B.; Moreno, N.; Patricio, A.; Martínez, I.; Aguilera, J.; Limon, I.D. The carboxyl-terminal domain of the heavy chain of tetanus toxin prevents dopaminergic degeneration and improves motor behavior in rats with striatal MPP(+)-lesions. Neurosci. Res. 2009, 65, 98–106. [Google Scholar] [CrossRef]
- Oliván, S.; Calvo, A.C.; Rando, A.; Herrando-Grabulosa, M.; Manzano, R.; Zaragoza, P.; Tizzano, E.F.; Aquilera, J.; Osta, R. Neuroprotective Effect of Non-viral Gene Therapy Treatment Based on Tetanus Toxin C-fragment in a Severe Mouse Model of Spinal Muscular Atrophy. Front. Mol. Neurosci. 2016, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.C.; Moreno-Igoa, M.; Mancuso, R.; Manzano, R.; Oliván, S.; Muñoz, M.J.; Penas, C.; Zaragoza, P.; Navarro, X.; Osta, R. Lack of a synergistic effect of a non-viral ALS gene therapy based on BDNF and a TTC fusion molecule. Orphanet J. Rare Dis. 2011, 6, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Igoa, M.; Calvo, A.C.; Penas, C.; Manzano, R.; Oliván, S.; Muñoz, M.J.; Mancuso, R.; Zaragoza, P.; Aguilera, J.; Navarro, X.; et al. Fragment C of tetanus toxin, more than a carrier. Novel perspectives in non-viral ALS gene therapy. J. Mol. Med. 2010, 88, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Igoa, M.; Calvo, A.C.; Ciriza, J.; Muñoz, M.J.; Zaragoza, P.; Osta, R. Non-viral gene delivery of the GDNF, either alone or fused to the C-fragment of tetanus toxin protein, prolongs survival in a mouse ALS model. Restor. Neurol. Neurosci. 2012, 30, 69–80. [Google Scholar] [CrossRef]
- Gil, C.; Chaib-Oukadour, I.; Blasi, J.; Aguilera, J. HC fragment (C-terminal portion of the heavy chain) of tetanus toxin activates protein kinase C isoforms and phosphoproteins involved in signal transduction. Biochem. J. 2001, 356 Pt 1, 97–103. [Google Scholar] [CrossRef]
- Gil, C.; Chaib-Oukadour, I.; Aguilera, J. C-terminal fragment of tetanus toxin heavy chain activates Akt and MEK/ERK signalling pathways in a Trk receptor-dependent manner in cultured cortical neurons. Biochem. J. 2003, 373 Pt 2, 613–620. [Google Scholar] [CrossRef]
- Sozbilen, M.C.; Ozturk, M.; Kaftan, G.; Dagci, T.; Ozyalcin, H.; Armagan, G. Neuroprotective Effects of C-terminal Domain of Tetanus Toxin on Rat Brain Against Motorneuron Damages After Experimental Spinal Cord Injury. Spine 2018, 43, E327–E333. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, J.D.; Jin, L.I.N.; Dykes-Hoberg, M.; Kuncl, R.W. Chronic inhibition of glutamate uptake produces a model of slow neurotoxicity. Proc. Natl. Acad. Sci. USA 1993, 90, 6591–6595. [Google Scholar] [CrossRef] [Green Version]
- Ferraiuolo, L.; Kirby, J.; Grierson, A.J.; Sendtner, M.; Shaw, P.J. Molecular pathways of motor neuron injury in amyotrophic lateral sclerosis. Nat. Rev. Neurol. 2011, 7, 616–630. [Google Scholar] [CrossRef]
- Rothstein, J.D.; Martin, L.J.; Kuncl, R.W. Decreased glutamate transport by the brain and spinal cord in amyotrophic lateral sclerosis. N. Engl. J. Med. 1992, 326, 1464–1468. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, J.D.; Van Kammen, M.; Levey, A.I.; Martin, L.J.; Kuncl, R.W. Selective loss of glial glutamate transporter GLT-1 in amyotrophic lateral sclerosis. Ann. Neurol. 1995, 38, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Fray, A.E.; Ince, P.G.; Banner, S.J.; Milton, I.D.; Usher, P.A.; Cookson, M.R.; Shaw, P.J. The expression of the glial glutamate transporter protein EAAT2 in motor neuron disease: An immunohistochemical study. Eur. J. Neurosci. 1998, 10, 2481–2489. [Google Scholar] [CrossRef] [PubMed]
- Foran, E.; Trotti, D. Glutamate transporters and the excitotoxic path to motor neuron degeneration in amyotrophic lateral sclerosis. Antioxid. Redox Signal. 2009, 11, 1587–1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matyja, E.; Nagańska, E.; Taraszewska, A.; Rafatowska, J. The mode of spinal motor neurons degeneration in a model of slow glutamate excitotoxicity in vitro. Folia Neuropathol. 2005, 43, 7–13. [Google Scholar]
- Matyja, E.; Taraszewska, A.; Nagańska, E.; Rafałowska, J. Autophagic degeneration of motor neurons in a model of slow glutamate excitotoxicity in vitro. Ultrastruct. Pathol. 2005, 29, 331–339. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrando-Grabulosa, M.; Casas, C.; Talbot, K.; Aguilera, J. Neurotrophic Properties of C-Terminal Domain of the Heavy Chain of Tetanus Toxin on Motor Neuron Disease. Toxins 2020, 12, 666. https://doi.org/10.3390/toxins12100666
Herrando-Grabulosa M, Casas C, Talbot K, Aguilera J. Neurotrophic Properties of C-Terminal Domain of the Heavy Chain of Tetanus Toxin on Motor Neuron Disease. Toxins. 2020; 12(10):666. https://doi.org/10.3390/toxins12100666
Chicago/Turabian StyleHerrando-Grabulosa, Mireia, Caty Casas, Kevin Talbot, and José Aguilera. 2020. "Neurotrophic Properties of C-Terminal Domain of the Heavy Chain of Tetanus Toxin on Motor Neuron Disease" Toxins 12, no. 10: 666. https://doi.org/10.3390/toxins12100666
APA StyleHerrando-Grabulosa, M., Casas, C., Talbot, K., & Aguilera, J. (2020). Neurotrophic Properties of C-Terminal Domain of the Heavy Chain of Tetanus Toxin on Motor Neuron Disease. Toxins, 12(10), 666. https://doi.org/10.3390/toxins12100666