Chronodisruption: A Poorly Recognized Feature of CKD



Abstract
:1. Introduction: The Growing Global Health Burden of Chronic Kidney Disease
2. Biological Rhythms
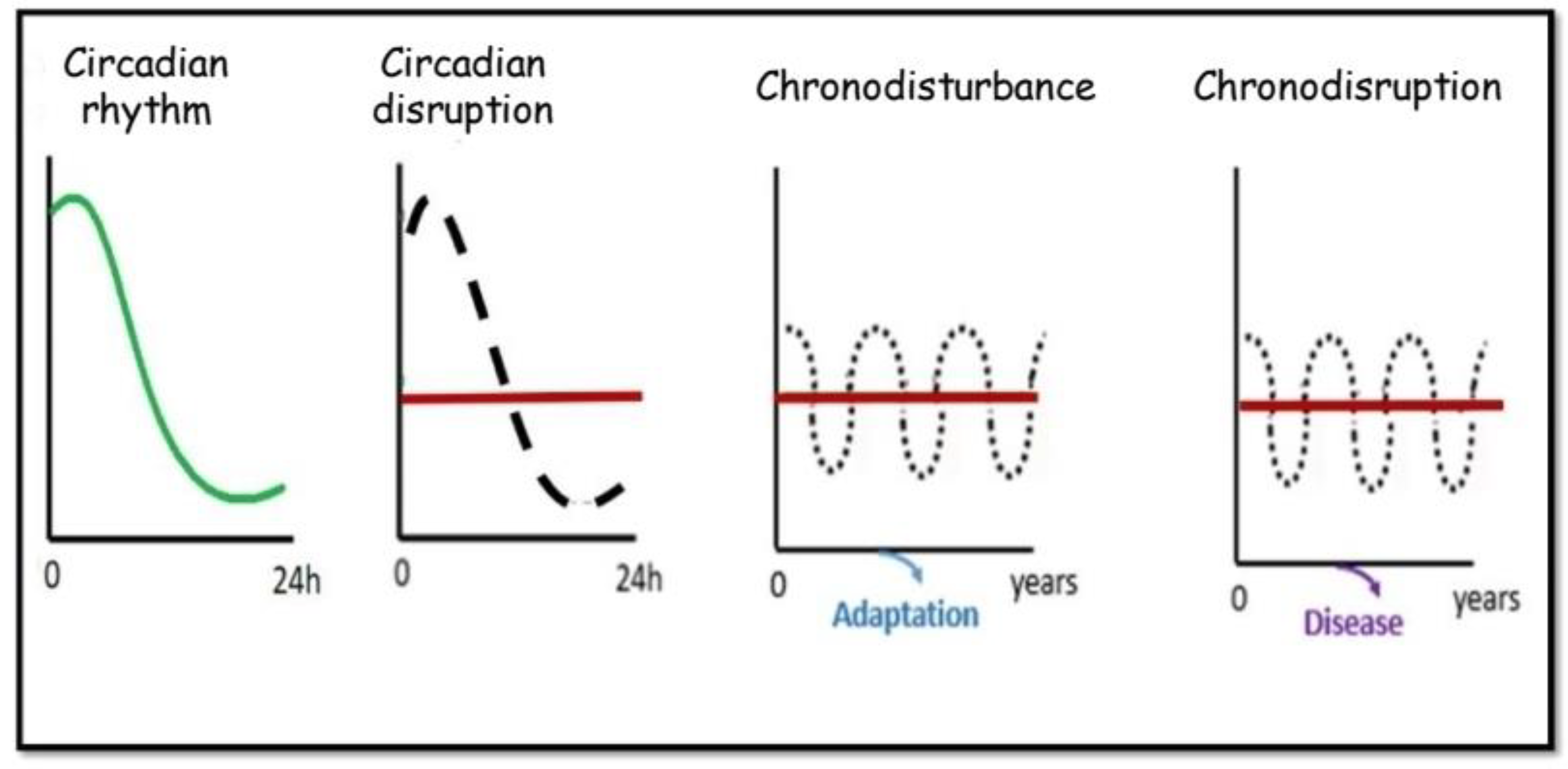
3. Concept of Chronodisruption
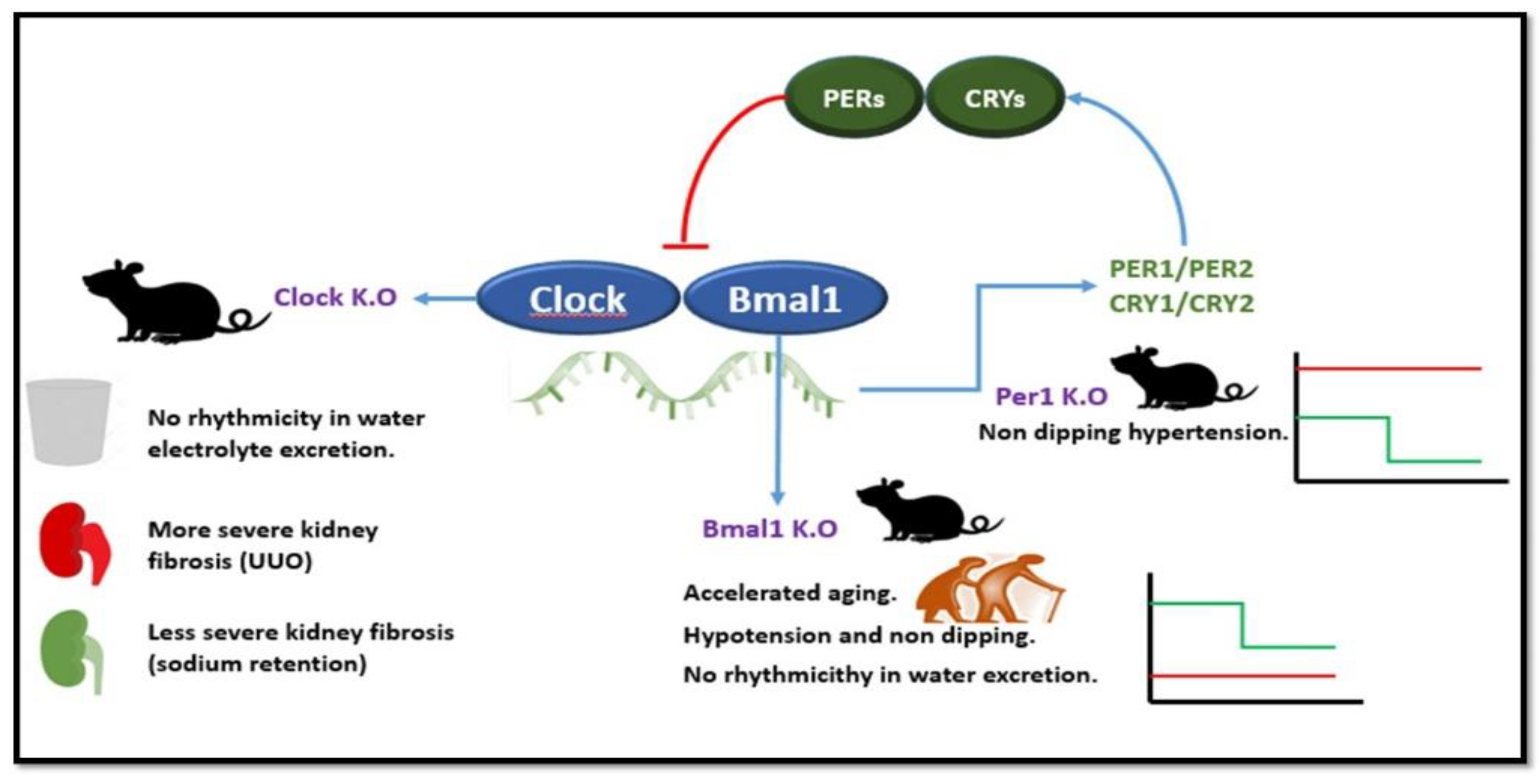
4. Chronodisruption in CKD
5. Chronodisruptors as Therapeutic Targets in CKD
5.1. Dietary Clues
5.2. Light Clues
5.3. Kidney Inflammation
5.4. Uremic Toxins
5.5. Disrupted HIF Activation and EPO Production
5.6. Physical Inactivity
5.7. Integration of Several Chronodisruptors
6. The Way Forward
Author Contributions
Funding
Conflicts of Interest
References
- Perez-Gomez, M.V.; Bartsch, L.A.; Castillo-Rodriguez, E.; Fernandez-Prado, R.; Fernandez-Fernandez, B.; Martin-Cleary, C.; Gracia-Iguacel, C.; Ortiz, A. Clarifying the concept of chronic kidney disease for non-nephrologists. Clin. Kidney J. 2019, 12, 258–261. [Google Scholar] [CrossRef]
- Foreman, K.J.; Marquez, N.; Dolgert, A.; Fukutaki, K.; Fullman, N.; McGaughey, M.; Pletcher, M.A.; Smith, A.E.; Tang, K.; Yuan, C.W.; et al. Forecasting life expectancy, years of life lost, and all-cause and cause-specific mortality for 250 causes of death: Reference and alternative scenarios for 2016-40 for 195 countries and territories. Lancet 2018, 392, 2052–2090. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, A.; Sanchez-Niño, M.D.; Crespo-Barrio, M.; De-Sequera-Ortiz, P.; Fernández-Giráldez, E.; García-Maset, R.; Macía-Heras, M.; Pérez-Fontán, M.; Rodríguez-Portillo, M.; Salgueira-Lazo, M.; et al. The Spanish Society of Nephrology (SENEFRO) commentary to the Spain GBD 2016 report: Keeping chronic kidney disease out of sight of health authorities will only magnify the problem. Nefrologia 2019, 39, 29–34. [Google Scholar] [CrossRef]
- Fernandez-Fernandez, B.; Fernandez-Prado, R.; Górriz, J.L.; Martinez-Castelao, A.; Navarro-González, J.F.; Porrini, E.; Soler, M.J.; Ortiz, A. Canagliflozin and Renal Events in Diabetes with Established Nephropathy Clinical Evaluation and Study of Diabetic Nephropathy with Atrasentan: What was learned about the treatment of diabetic kidney disease with canagliflozin and atrasentan? Clin. Kidney J. 2019, 12, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Sarafidis, P.; Ferro, C.J.; Morales, E.; Ortiz, A.; Malyszko, J.; Hojs, R.; Khazim, K.; Ekart, R.; Valdivielso, J.; Fouque, D.; et al. SGLT-2 inhibitors and GLP-1 receptor agonists for nephroprotection and cardioprotection in patients with diabetes mellitus and chronic kidney disease. A consensus statement by the EURECA-m and the DIABESITY working groups of the ERA-EDTA. Nephrol. Dial. Transplant. 2019, 34, 208–230. [Google Scholar] [CrossRef]
- Herrington, W.G.; Preiss, D.; Haynes, R.; von Eynatten, M.; Staplin, N.; Hauske, S.J.; George, J.T.; Green, J.B.; Landray, M.J.; Baigent, C.; et al. The potential for improving cardio-renal outcomes by sodium-glucose co-transporter-2 inhibition in people with chronic kidney disease: A rationale for the EMPA-KIDNEY study. Clin. Kidney J. 2018, 11, 749–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, B.; Mancia, G.; Spiering, W.; Agabiti Rosei, E.; Azizi, M.; Burnier, M.; Clement, D.; Coca, A.; De Simone, G.; Dominiczak, A.; et al. 2018 Practice Guidelines for the management of arterial hypertension of the European Society of Hypertension and the European Society of Cardiology: ESH/ESC Task Force for the Management of Arterial Hypertension. J. Hypertens. 2018, 36, 2284–2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Niño, M.D.; Sanz, A.B.; Ramos, A.M.; Ruiz-Ortega, M.; Ortiz, A. Translational science in chronic kidney disease. Clin. Sci. (Lond.) 2017, 131, 1617–1629. [Google Scholar] [CrossRef]
- Fernandez-Prado, R.; Esteras, R.; Perez-Gomez, M.V.; Gracia-Iguacel, C.; Gonzalez-Parra, E.; Sanz, A.B.; Ortiz, A.; Sanchez-Niño, M.D. Nutrients Turned into Toxins: Microbiota Modulation of Nutrient Properties in Chronic Kidney Disease. Nutrients 2017, 9, 489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo-Rodríguez, E.; Pizarro-Sánchez, S.; Sanz, A.B.; Ramos, A.M.; Sanchez-Niño, M.D.; Martin-Cleary, C.; Fernandez-Fernandez, B.; Ortiz, A. Inflammatory Cytokines as Uremic Toxins: “Ni Son Todos Los Que Estan, Ni Estan Todos Los Que Son”. Toxins 2017, 9, 114. [Google Scholar]
- Duranton, F.; Cohen, G.; De Smet, R.; Rodriguez, M.; Jankowski, J.; Vanholder, R.; Argiles, A. European Uremic Toxin Work Group. Norm. Pathol. Conc. Urem. Toxins J. Am. Soc. Nephrol. 2012, 23, 1258–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Fernandez, B.; Izquierdo, M.C.; Valiño-Rivas, L.; Nastou, D.; Sanz, A.B.; Ortiz, A.; Sanchez-Niño, M.D. Albumin downregulates Klotho in tubular cells. Nephrol. Dial. Transplant. 2018, 33, 1712–1722. [Google Scholar] [CrossRef] [Green Version]
- Rossignol, P.; Massy, Z.A.; Azizi, M.; Bakris, G.; Ritz, E.; Covic, A.; Goldsmith, D.; Heine, G.H.; Jager, K.J.; Kanbay, M.; et al. The double challenge of resistant hypertension and chronic kidney disease. Lancet 2015, 386, 1588–1598. [Google Scholar] [CrossRef]
- Firsov, D.; Bonny, O. Circadian rhythms and the kidney. Nat. Rev. Nephrol. 2018, 14, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Firsov, D.; Bonny, O. Circadian regulation of renal function. Kidney Int. 2010, 78, 640–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Nicolas, A.; Madrid, J.A.; Rol, M.A. Day-night contrast as source of health for the human circadian system. Chronobiol. Int. 2014, 31, 382–393. [Google Scholar] [CrossRef]
- Zhang, D.; Pollock, D.M. Diurnal Regulation of Renal Electrolyte Excretion: The Role of Paracrine Factors. Annu. Rev. Physiol. 2019, 82, 343–363. [Google Scholar] [CrossRef] [Green Version]
- Chiou, Y.Y.; Yang, Y.; Rashid, N.; Ye, R.; Selby, C.P.; Sancar, A. Mammalian Period represses and de-represses transcription by displacing CLOCK-BMAL1 from promoters in a Cryptochrome-dependent manner. Proc. Natl. Acad. Sci. USA 2016, 113, E6072–E6079. [Google Scholar] [CrossRef] [Green Version]
- Mészáros, K.; Pruess, L.; Szabó, A.J.; Gondan, M.; Ritz, E.; Schaefer, F. Development of the circadian clockwork in the kidney. Kidney Int. 2014, 86, 915–922. [Google Scholar] [CrossRef] [Green Version]
- Becker, B.K.; Zhang, D.; Soliman, R.; Pollock, D.M. Autonomic nerves and circadian control of renal function. Auton. Neurosci. 2019, 217, 58–65. [Google Scholar] [CrossRef]
- Motohashi, H.; Tahara, Y.; Whittaker, D.S.; Wang, H.B.; Yamaji, T.; Wakui, H.; Haraguchi, A.; Yamazaki, M.; Miyakawa, H.; Hama, K.; et al. The circadian clock is disrupted in mice with adenine-induced tubulointerstitial nephropathy. Kidney Int. 2020. [Google Scholar] [CrossRef] [Green Version]
- Hara, M.; Minami, Y.; Ohashi, M.; Tsuchiya, Y.; Kusaba, T.; Tamagaki, K.; Koike, N.; Umemura, Y.; Inokawa, H.; Yagita, K. Robust circadian clock oscillation and osmotic rhythms in inner medulla reflecting cortico-medullary osmotic gradient rhythm in rodent kidney. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nikolaeva, S.; Ansermet, C.; Centeno, G.; Pradervand, S.; Bize, V.; Mordasini, D.; Henry, H.; Koesters, R.; Maillard, M.; Bonny, O.; et al. Nephron-Specific Deletion of Circadian Clock Gene Bmal1 Alters the Plasma and Renal Metabolome and Impairs Drug Disposition. J. Am. Soc. Nephrol. 2016, 27, 2997–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erren, T.C.; Reiter, R.J.; Piekarski, C. Light, timing of biological rhythms, and chronodisruption in man. Naturwissenschaften 2003, 90, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Erren, T.C.; Reiter, R.J. Defining chronodisruption. J. Pineal Res. 2009, 46, 245–247. [Google Scholar] [CrossRef]
- Bonny, O.; Vinciguerra, M.; Gumz, M.L.; Mazzoccoli, G. Molecular bases of circadian rhythmicity in renal physiology and pathology. Nephrol. Dial. Transplant. 2013, 28, 2421–2431. [Google Scholar] [CrossRef] [Green Version]
- Mendez, N.; Torres-Farfan, C.; Salazar, E.; Bascur, P.; Bastidas, C.; Vergara, K.; Spichiger, C.; Halabi, D.; Vio, C.P.; Richter, H.G. Fetal Programming of Renal Dysfunction and High Blood Pressure by Chronodisruption. Front. Endocrinol. 2019, 10, 362. [Google Scholar] [CrossRef] [Green Version]
- Wuerzner, G.; Firsov, D.; Bonny, O. Circadian glomerular function: From physiology to molecular and therapeutical aspects. Nephrol. Dial. Transplant. 2014, 29, 1475–1480. [Google Scholar] [CrossRef] [Green Version]
- Firsov, D.; Tokonami, N.; Bonny, O. Role of the renal circadian timing system in maintaining water and electrolytes homeostasis. Mol. Cell. Endocrinol. 2012, 349, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Argilés, A.; Mourad, G.; Mion, C. Seasonal changes in blood pressure in patients with end-stage renal disease treated with hemodialysis. N. Engl. J. Med. 1998, 339, 1364–1370. [Google Scholar] [CrossRef]
- Knutson, K.L.; Lash, J.; Ricardo, A.C.; Herdegen, J.; Thornton, J.D.; Rahman, M.; Turek, N.; Cohan, J.; Lawrence, J.; Bazzano, L.; et al. Habitual sleep and kidney function in chronic kidney disease: The Chronic Renal Insufficiency Cohort study. J. Sleep Res. 2018, 27, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, N.; Ishigaki, S.; Isobe, S. The pivotal role of melatonin in ameliorating chronic kidney disease by suppression of the renin-angiotensin system in the kidney. Hypertens. Res. 2019, 42, 761–768. [Google Scholar] [CrossRef]
- Voulgaris, A.; Marrone, O.; Bonsignore, M.R.; Steiropoulos, P. Chronic kidney disease in patients with obstructive sleep apnea. Narrat. Rev. Sleep Med. Rev. 2019, 47, 74–89. [Google Scholar] [CrossRef] [PubMed]
- Sarafidis, P.A.; Persu, A.; Agarwal, R.; Burnier, M.; de Leeuw, P.; Ferro, C.J.; Halimi, J.M.; Heine, G.H.; Jadoul, M.; Jarraya, F.; et al. Hypertension in dialysis patients: A consensus document by the European Renal and Cardiovascular Medicine (EURECA-m) working group of the European Renal Association-European Dialysis and Transplant Association (ERA-EDTA) and the Hypertension and the Kidney working group of the European Society of Hypertension (ESH). Nephrol. Dial. Transplant. 2017, 32, 620–640. [Google Scholar] [PubMed]
- Bolignano, D.; Rastelli, S.; Agarwal, R.; Fliser, D.; Massy, Z.; Ortiz, A.; Wiecek, A.; Martinez-Castelao, A.; Covic, A.; Goldsmith, D.; et al. Pulmonary hypertension in CKD. Am. J. Kidney Dis. 2013, 61, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Di Daniele, N.; Fegatelli, D.A.; Rovella, V.; Castagnola, V.; Gabriele, M.; Scuteri, A. Circadian blood pressure patterns and blood pressure control in patients with chronic kidney disease. Atherosclerosis 2017, 267, 139–145. [Google Scholar] [CrossRef]
- Ruiz-Hurtado, G.; Ruilope, L.; De la Sierra, A.; Sarafidis, P.; De la Cruz, J.; Gorostidi, M.; Segura, J.; Vinyoles, E.; Banegas, J. Association between High and Very High Albuminuria and Nighttime Blood Pressure: Influence of Diabetes and Chronic Kidney Disease. Diabetes Care 2016, 39, 1729–1737. [Google Scholar] [CrossRef] [Green Version]
- Ando, D.; Yasuda, G. Circadian Blood Pressure Rhythm Is Changed by Improvement in Hypoalbuminemia and Massive Proteinuria in Patients with Minimal Change Nephrotic Syndrome. Cardiorenal. Med. 2016, 6, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Ohkubo, T.; Hozawa, A.; Yamaguchi, J.; Kikuya, M.; Ohmori, K.; Michimata, M.; Matsubara, M.; Hashimoto, J.; Hoshi, H.; Araki, T.; et al. Prognostic significance of the nocturnal decline in blood pressure in individuals with and without high 24-h blood pressure: The Ohasama study. J. Hypertens. 2002, 20, 2183–2189. [Google Scholar] [CrossRef]
- Fedecostante, M.; Spannella, F.; Cola, G.; Espinosa, E.; Dessì-Fulgheri, P.; Sarzani, R. Chronic kidney disease is characterized by “double trouble” higher pulse pressure plus night-time systolic blood pressure and more severe cardiac damage. PLoS ONE 2014, 9, e86155. [Google Scholar] [CrossRef]
- Huang, J.T.; Cheng, H.M.; Yu, W.C.; Lin, Y.P.; Sung, S.H.; Chen, C.H. Increased Nighttime Pulse Pressure Variability but Not Ambulatory Blood Pressure Levels Predicts 14-Year All-Cause Mortality in Patients on Hemodialysis. Hypertension 2019, 74, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Tokonami, N.; Mordasini, D.; Pradervand, S.; Centeno, G.; Jouffe, C.; Maillard, M.; Bonny, O.; Gachon, F.; Gomez, R.A.; Sequeira-Lopez, M.L. Local renal circadian clocks control fluid-electrolyte homeostasis and BP. J. Am. Soc. Nephrol. 2014, 25, 1430–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivy, J.R.; Jones, N.K.; Costello, H.M.; Mansley, M.K.; Peltz, T.S.; Flatman, P.W.; Bailey, M.A. Glucocorticoid receptor activation stimulates the sodium-chloride cotransporter and influences the diurnal rhythm of its phosphorylation. Am. J. Physiol. Renal Physiol. 2019, 317, F1536–F1548. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Kawamoto, T.; Noshiro, M.; Ueno, T.; Doi, S.; Honda, K.; Masaki, T.; Higashi, Y.; Kato, Y. Dec1 and CLOCK Regulate Na. Hypertension 2018, 72, 746–754. [Google Scholar] [CrossRef]
- Valiño-Rivas, L.; Cuarental, L.; Agustin, M.; Husi, H.; Cannata-Ortiz, P.; Sanz, A.B.; Mischak, H.; Ortiz, A.; Sanchez-Niño, M.D. MAGE genes in the kidney: Identification of MAGED2 as upregulated during kidney injury and in stressed tubular cells. Nephrol. Dial. Transplant. 2019, 34, 1498–1507. [Google Scholar] [CrossRef]
- Gil, R.B.; Ortiz, A.; Sanchez-Niño, M.D.; Markoska, K.; Schepers, E.; Vanholder, R.; Glorieux, G.; Schmitt-Kopplin, P.; Heinzmann, S. Increased urinary osmolyte excretion indicates chronic kidney disease severity and progression rate. Nephrol. Dial. Transplant. 2018, 33, 2156–2164. [Google Scholar] [CrossRef]
- Myung, J.; Wu, M.Y.; Lee, C.Y.; Rahim, A.R.; Truong, V.H.; Wu, D.; Piggins, H.D.; Wu, M.S. The Kidney Clock Contributes to Timekeeping by the Master Circadian Clock. Int. J. Mol. Sci. 2019, 20, 2765. [Google Scholar] [CrossRef] [Green Version]
- Gansevoort, R.; Arici, M.; Benzing, T.; Birn, H.; Capasso, G.; Covic, A.; Devuyst, O.; Drechsler, C.; Eckardt, K.U.; Emma, F.; et al. Recommendations for the use of tolvaptan in autosomal dominant polycystic kidney disease: A position statement on behalf of the ERA-EDTA Working Groups on Inherited Kidney Disorders and European Renal Best Practice. Nephrol. Dial. Transplant. 2016, 31, 337–348. [Google Scholar] [CrossRef]
- Torres, V.E.; Chapman, A.B.; Devuyst, O.; Gansevoort, R.T.; Perrone, R.D.; Koch, G.; Ouyang, J.; McQuade, R.D.; Blais, J.D.; Czerwiec, F.S.; et al. Tolvaptan in Later-Stage Autosomal Dominant Polycystic Kidney Disease. N. Engl. J. Med. 2017, 377, 1930–1942. [Google Scholar] [CrossRef]
- Montero, D.; Diaz-Canestro, C.; Oberholzer, L.; Lundby, C. The role of blood volume in cardiac dysfunction and reduced exercise tolerance in patients with diabetes. Lancet Diabetes Endocrinol. 2019, 7, 807–816. [Google Scholar] [CrossRef]
- Parizadeh, S.M.; Ghandehari, M.; Parizadeh, M.R.; Ferns, G.A.; Ghayour-Mobarhan, M.; Avan, A.; Hassanian, S. The diagnostic and prognostic value of copeptin in cardiovascular disease, current status, and prospective. J. Cell. Biochem. 2018, 119, 7913–7923. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, E.E.; Johnson, E.C.; Yusifova, M.; Bruns, D.R. The renal molecular clock: Broken by aging and restored by exercise. Am. J. Physiol. Renal Physiol. 2019, 317, F1087–F1093. [Google Scholar] [CrossRef]
- Batinga, H.; Martinez-Nicolas, A.; Zornoza-Moreno, M.; Sánchez-Solis, M.; Larqué, E.; Mondéjar, M.T.; Moreno-Casbas, M.; García, F.J.; Campos, M.; Rol, M.A.; et al. Ontogeny and aging of the distal skin temperature rhythm in humans. Age 2015, 37, 29. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, N.; Ishigaki, S.; Isobe, S.; Matsuyama, T.; Sato, T.; Fujikura, T.; Tsuji, T.; Kato, A.; Yasuda, H. Salt Loading Aggravates the Relationship between Melatonin and Proteinuria in Patients with Chronic Kidney Disease. Intern. Med. 2019, 58, 1557–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speed, J.S.; Hyndman, K.A.; Roth, K.; Heimlich, J.B.; Kasztan, M.; Fox, B.M.; Johnston, J.G.; Becker, B.K.; Jin, C.; Gamble, K.L.; et al. High dietary sodium causes dyssynchrony of the renal molecular clock in rats. Am. J. Physiol. Renal Physiol. 2018, 314, F89–F98. [Google Scholar] [CrossRef] [PubMed]
- Daimiel-Ruiz, L.; Klett-Mingo, M.; Konstantinidou, V.; Micó, V.; Aranda, J.F.; García, B.; Martínez-Botas, J.; Dávalos, A.; Fernández-Hernando, C.; Ordovás, J.M. Dietary lipids modulate the expression of miR-107, a miRNA that regulates the circadian system. Mol. Nutr. Food Res. 2015, 59, 1865–1878. [Google Scholar] [CrossRef] [Green Version]
- Laermans, J.; Depoortere, I. Chronobesity: Role of the circadian system in the obesity epidemic. Obes. Rev. 2016, 17, 108–125. [Google Scholar] [CrossRef]
- Galindo Muñoz, J.S.; Gómez Gallego, M.; Díaz Soler, I.; Barberá Ortega, M.C.; Martínez Cáceres, C.M.; Hernández Morante, J.J. Effect of a chronotype-adjusted diet on weight loss effectiveness: A randomized clinical trial. Clin. Nutr. 2019. [Google Scholar] [CrossRef]
- Bandín, C.; Scheer, F.A.; Luque, A.J.; Ávila-Gandía, V.; Zamora, S.; Madrid, J.A. Meal timing affects glucose tolerance, substrate oxidation and circadian-related variables: A randomized, crossover trial. Int. J. Obes. 2015, 39, 828–833. [Google Scholar]
- Challet, E. The circadian regulation of food intake. Nat. Rev. Endocrinol. 2019, 15, 393–405. [Google Scholar] [CrossRef]
- Parkar, S.G.A.; Cheeseman, J.F. Potential Role for the Gut Microbiota in Modulating Host Circadian Rhythms and Metabolic Health. Microorganisms 2019, 7, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilera-Correa, J.-J.; Madrazo-Clemente, P.; Martínez-Cuesta, M.D.C.; Peláez, C.; Ortiz, A.; Sánchez-Niño, M.D.; Esteban, J.; Requena, T. Lyso-Gb3 modulates the gut microbiota and decreases butyrate production. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perna, A.F.; Glorieux, G.; Zacchia, M.; Trepiccione, F.; Capolongo, G.; Vigorito, C.; Anishchenko, E.; Ingrosso, D. The role of the intestinal microbiota in uremic solute accumulation: A focus on sulfur compounds. J. Nephrol. 2019, 32, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Joossens, M.; Faust, K.; Gryp, T.; Nguyen, A.T.L.; Wang, J.; Eloot, S.; Schepers, E.; Dhondt, A.; Pletinck, A.; Vieira-Silva, S.; et al. Gut microbiota dynamics and uraemic toxins: One size does not fit all. Gut 2019, 68, 2257–2260. [Google Scholar] [CrossRef]
- Erren, T.C.; Morfeld, P.; Groß, J.V.; Wild, U.; Lewis, P. IARC 2019: “Night shift work” is probably carcinogenic: What about disturbed chronobiology in all walks of life? J. Occup. Med. Toxicol. 2019, 14, 29. [Google Scholar] [CrossRef]
- Uhm, J.Y.; Kim, H.R.; Kang, G.H.; Choi, Y.G.; Park, T.H.; Kim, S.Y.; Chang, S.S.; Choo, W.O. The association between shift work and chronic kidney disease in manual labor workers using data from the Korea National Health and Nutrition Examination Survey (KNHANES 2011–2014). Ann. Occup. Environ. Med. 2018, 30, 69. [Google Scholar] [CrossRef]
- Sanz, A.B.; Ruiz-Andres, O.; Sanchez-Niño, M.D.; Ruiz-Ortega, M.; Ramos, A.M.; Ortiz, A. Out of the TWEAKlight: Elucidating the Role of Fn14 and TWEAK in Acute Kidney Injury. Semin. Nephrol. 2016, 36, 189–198. [Google Scholar] [CrossRef]
- Sanz, A.B.; Izquierdo, M.C.; Sanchez-Niño, M.D.; Ucero, A.C.; Egido, J.; Ruiz-Ortega, M.; Ramos, A.M.; Putterman, C.; Ortiz, A. TWEAK and the progression of renal disease: Clinical translation. Nephrol. Dial. Transplant. 2014, 29 (Suppl. 1), i54–i62. [Google Scholar] [CrossRef] [Green Version]
- Valiño-Rivas, L.; Vaquero, J.J.; Sucunza, D.; Gutierrez, S.; Sanz, A.B.; Fresno, M.; Ortiz, A.; Sanchez-Niño, M.D. NIK as a Druggable Mediator of Tissue Injury. Trends Mol. Med. 2019, 25, 341–360. [Google Scholar] [CrossRef]
- Valiño-Rivas, L.; Gonzalez-Lafuente, L.; Sanz, A.B.; Ruiz-Ortega, M.; Ortiz, A.; Sanchez-Niño, M.D. Non-canonical NFκB activation promotes chemokine expression in podocytes. Sci. Rep. 2016, 6, 28857. [Google Scholar] [CrossRef] [Green Version]
- Sanz, A.B.; Sanchez-Niño, M.D.; Izquierdo, M.C.; Jakubowski, A.; Justo, P.; Blanco-Colio, L.M.; Blanco-Colio, L.M.; Ruiz-Ortega, M.; Selgas, R.; Egido, J.; et al. TWEAK activates the non-canonical NFkappaB pathway in murine renal tubular cells: Modulation of CCL21. PLoS ONE 2010, 5, e8955. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, A.; Husi, H.; Gonzalez-Lafuente, L.; Valiño-Rivas, L.; Fresno, M.; Sanz, A.B.; Mullen, W.; Albalat, A.; Mezzano, S.; Vlahou, T.; et al. Mitogen-Activated Protein Kinase 14 Promotes AKI. J. Am. Soc. Nephrol. 2017, 28, 823–836. [Google Scholar] [CrossRef] [Green Version]
- Cuarental, L.; Sucunza-Sáenz, D.; Valiño-Rivas, L.; Fernandez-Fernandez, B.; Sanz, A.B.; Ortiz, A.; Vaquero, J.J.; Sanchez-Niño, M.D. MAP3K kinases and kidney injury. Nefrologia 2019, 39, 568–580. [Google Scholar] [CrossRef] [PubMed]
- Sanz, A.B.; Sanchez-Niño, M.D.; Ramos, A.M.; Moreno, J.A.; Santamaria, B.; Ruiz-Ortega, M.; Mullen, W.; Albalat, A.; Mezzano, S.; Vlahou, T.; et al. NF-kappaB in renal inflammation. J. Am. Soc. Nephrol. 2010, 21, 1254–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poveda, J.; Tabara, L.C.; Fernandez-Fernandez, B.; Martin-Cleary, C.; Sanz, A.B.; Selgas, R.; Ortiz, A.; Sanchez-Niño, M.D. TWEAK/Fn14 and Non-Canonical NF-kappaB Signaling in Kidney Disease. Front. Immunol. 2013, 4, 447. [Google Scholar] [CrossRef] [Green Version]
- Bellet, M.M.; Zocchi, L.; Sassone-Corsi, P. The RelB subunit of NFκB acts as a negative regulator of circadian gene expression. Cell Cycle 2012, 11, 3304–3311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poveda, J.; Sanz, A.B.; Carrasco, S.; Ruiz-Ortega, M.; Cannata-Ortiz, P.; Sanchez-Niño, M.D.; Ortiz, A. Bcl3: A regulator of NF-κB inducible by TWEAK in acute kidney injury with anti-inflammatory and antiapoptotic properties in tubular cells. Exp. Mol. Med. 2017, 49, e352. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.A.; Izquierdo, M.C.; Sanchez-Niño, M.D.; Suárez-Alvarez, B.; Lopez-Larrea, C.; Jakubowski, A.; Blanco, J.; Ramirez, R.; Selgas, R.; Ruiz-Ortega, M.; et al. The inflammatory cytokines TWEAK and TNFα reduce renal klotho expression through NFκB. J. Am. Soc. Nephrol. 2011, 22, 1315–1325. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Andres, O.; Sanchez-Niño, M.D.; Moreno, J.A.; Ruiz-Ortega, M.; Ramos, A.M.; Sanz, A.B.; Ortiz, A. Downregulation of kidney protective factors by inflammation: Role of transcription factors and epigenetic mechanisms. Am. J. Physiol. Renal Physiol. 2016, 311, F1329–F1340. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Andres, O.; Suarez-Alvarez, B.; Sánchez-Ramos, C.; Monsalve, M.; Sanchez-Niño, M.D.; Ruiz-Ortega, M.; Egido, J.; Ortiz, A.; Sanz, A.B. The inflammatory cytokine TWEAK decreases PGC-1α expression and mitochondrial function in acute kidney injury. Kidney Int. 2016, 89, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Millet, P.; McCall, C.; Yoza, B. RelB: An outlier in leukocyte biology. J. Leukoc. Biol. 2013, 94, 941–951. [Google Scholar] [CrossRef]
- Sato, F.; Otsuka, T.; Kohsaka, A.; Le, H.T.; Bhawal, U.K.; Muragaki, Y. Smad3 Suppresses Epithelial Cell Migration and Proliferation via the Clock Gene Dec1, Which Negatively Regulates the Expression of Clock Genes Dec2 and Per1. Am. J. Pathol. 2019, 189, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Sallée, M.; Dou, L.; Cerini, C.; Poitevin, S.; Brunet, P.; Burtey, S. The aryl hydrocarbon receptor-activating effect of uremic toxins from tryptophan metabolism: A new concept to understand cardiovascular complications of chronic kidney disease. Toxins 2014, 6, 934–949. [Google Scholar] [CrossRef]
- Castillo-Rodriguez, E.; Fernandez-Prado, R.; Esteras, R.; Perez-Gomez, M.V.; Gracia-Iguacel, C.; Fernandez-Fernandez, B.; Kanbay, M.; Tejedor, A.; Lazaro, A.; Ruiz-Ortega, M.; et al. Impact of Altered Intestinal Microbiota on Chronic Kidney Disease Progression. Toxins 2018, 10, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tischkau, S.A. Mechanisms of circadian clock interactions with aryl hydrocarbon receptor signalling. Eur. J. Neurosci. 2019, 51, 379–395. [Google Scholar] [CrossRef] [Green Version]
- Jaeger, C.; Khazaal, A.Q.; Xu, C.; Sun, M.; Krager, S.L.; Tischkau, S.A. Aryl Hydrocarbon Receptor Deficiency Alters Circadian and Metabolic Rhythmicity. J. Biol. Rhythms 2017, 32, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J.; Jansen, K.; Neven, E.; Poesen, R.; Othman, A.; van Mil, A.; Sluijter, J.; Sastre Torano, J.; Zaal, E.A.; Berkers, C.R.; et al. Remote sensing and signaling in kidney proximal tubules stimulates gut microbiome-derived organic anion secretion. Proc. Natl. Acad. Sci. USA 2019, 116, 16105–16110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, J.S.; Borges, N.A.; Anjos, J.S.D.; Nakao, L.S.; Stockler-Pinto, M.B.; Paiva, B.R.; Cardoso-Weide, L.C.; Cardozo, L.F.M.F.; Mafra, D. Aryl Hydrocarbon Receptor and Uremic Toxins from the Gut Microbiota in Chronic Kidney Disease Patients: Is There a Relationship between Them? Biochemistry 2019, 58, 2054–2060. [Google Scholar] [CrossRef]
- Addi, T.; Poitevin, S.; McKay, N.; El Mecherfi, K.E.; Kheroua, O.; Jourde-Chiche, N.; de Macedo, A.; Gondouin, B.; Cerini, C.; Brunet, P.; et al. Mechanisms of tissue factor induction by the uremic toxin indole-3 acetic acid through aryl hydrocarbon receptor/nuclear factor-kappa B signaling pathway in human endothelial cells. Arch. Toxicol. 2019, 93, 121–136. [Google Scholar] [CrossRef]
- Dimova, E.Y.; Jakupovic, M.; Kubaichuk, K.; Mennerich, D.; Chi, T.F.; Tamanini, F.; Oklejewicz, M.; Hänig, J.; Byts, N.; Mäkelä, K.A.; et al. The Circadian Clock Protein CRY1 Is a Negative Regulator of HIF-1α. iScience 2019, 13, 284–304. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Morinibu, A.; Koyasu, S.; Goto, Y.; Hiraoka, M.; Harada, H. A circadian clock gene, PER2, activates HIF-1 as an effector molecule for recruitment of HIF-1α to promoter regions of its downstream genes. FEBS J. 2017, 284, 3804–3816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckle, T.; Hartmann, K.; Bonney, S.; Reithel, S.; Mittelbronn, M.; Walker, L.A.; Lowes, B.D.; Han, J.; Borchers, C.H.; Buttrick, P.M.; et al. Adora2b-elicited Per2 stabilization promotes a HIF-dependent metabolic switch crucial for myocardial adaptation to ischemia. Nat. Med. 2012, 18, 774–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peek, C.B.; Levine, D.C.; Cedernaes, J.; Taguchi, A.; Kobayashi, Y.; Tsai, S.J.; Bonar, N.A.; Mc, M.R.; Ramsey, K.M.; Bass, J. Circadian Clock Interaction with HIF1α Mediates Oxygenic Metabolism and Anaerobic Glycolysis in Skeletal Muscle. Cell Metab. 2017, 25, 86–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egg, M.; Köblitz, L.; Hirayama, J.; Schwerte, T.; Folterbauer, C.; Kurz, A.; Fiechtner, B.; Möst, M.; Salvenmoser, W.; Sassone, P.; et al. Linking oxygen to time: The bidirectional interaction between the hypoxic signaling pathway and the circadian clock. Chronobiol. Int. 2013, 30, 510–529. [Google Scholar] [CrossRef]
- Chen, N.; Hao, C.; Liu, B.C.; Lin, H.; Wang, C.; Xing, C.; Liang, X.; Jiang, G.; Liu, Z.; Li, X.; et al. Roxadustat Treatment for Anemia in Patients Undergoing Long-Term Dialysis. N. Engl. J. Med. 2019, 381, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, C.; Xu, C.; Sun, M.; Krager, S.; Tischkau, S.A. Aryl hydrocarbon receptor-deficient mice are protected from high fat diet-induced changes in metabolic rhythms. Chronobiol. Int. 2017, 34, 318–336. [Google Scholar] [CrossRef]
- Martinez-Nicolas, A.; Ortiz-Tudela, E.; Rol, M.A.; Madrid, J.A. Uncovering different masking factors on wrist skin temperature rhythm in free-living subjects. PLoS ONE 2013, 8, e61142. [Google Scholar] [CrossRef]
- Corbalán-Tutau, M.D.; Gómez-Abellán, P.; Madrid, J.A.; Canteras, M.; Ordovás, J.M.M. Toward a chronobiological characterization of obesity and metabolic syndrome in clinical practice. Clin. Nutr. 2015, 34, 477–483. [Google Scholar] [CrossRef]
- Wei, N.; Gumz, M.L.; Layton, A.T. Predicted effect of circadian clock modulation of NHE3 of a proximal tubule cell on sodium transport. Am. J. Physiol. Renal Physiol. 2018, 315, F665–F676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMullan, C.J.; Curhan, G.C.; Forman, J.P. Association of short sleep duration and rapid decline in renal function. Kidney Int. 2016, 89, 1324–1330. [Google Scholar] [CrossRef] [Green Version]
- De Lavallaz, L.; Musso, C.G. Chronobiology in nephrology: The influence of circadian rhythms on renal handling of drugs and renal disease treatment. Int. Urol. Nephrol. 2018, 50, 2221–2228. [Google Scholar] [CrossRef] [PubMed]
- Hermida, R.C.; Ayala, D.E.; Mojón, A.; Fernández, J.R. Risk of incident chronic kidney disease is better reduced by bedtime than upon-awakening ingestion of hypertension medications. Hypertens. Res. 2018, 41, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Niño, M.D.; Sanz, A.B.; Ramos, A.M.; Fernandez-Fernandez, B.; Ortiz, A. Clinical proteomics in kidney disease as an exponential technology: Heading towards the disruptive phase. Clin. Kidney J. 2017, 10, 188–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, N.; Hao, C.; Peng, X.; Lin, H.; Yin, A.; Hao, L.; Tao, Y.; Liang, X.; Liu, Z.; Xing, C. Roxadustat for Anemia in Patients with Kidney Disease Not Receiving Dialysis. N. Engl. J. Med. 2019, 381, 1001–1010. [Google Scholar] [CrossRef]
- Asfar, B.; Asfar, R.; Sag, A.; Kanbay, A.; Korkmaz, H.; Cipolla-Neto, J.; Covic, A.; Ortiz, A.; Kanbay, M. Sweet Dreams: Therapeutic Insights, Targeting Imaging and Physiologic Evidence Linking Sleep, Melatonin, and Diabetic Nephropathy. Clin. Kidney J. 2019. accepted. [Google Scholar]



{kind=link}
{kind=link}
| Glomeruli | Circulation and Interstitial | Tubular |
|---|---|---|
| Glomerular filtration rate | Renal plasma flow | Water and electrolyte (sodium, potassium, calcium, magnesium, phosphate) excretion and corticomedullary interstitial osmolarity gradient |
| Kidney oxygenation and erythropoietin production | H+ excretion |
| Diet | Other Lifestyle Factors | Endogenous Factors |
|---|---|---|
| Dietary components, e.g., sodium | Night shift work | Gut microbiota and microbiota-associated uremic toxins |
| Mistimed eating | Kidney inflammation, non-canonical NFκB activation and RelB | |
| Mediators of kidney fibrosis such as Smad3 |
| When Does Chronodisruption Start in CKD Natural History? | What Are the Key Chronodisruptors in CKD and What Are Their Targets? Can Chronodisruptors Be Targeted Therapeutically? | Other Questions |
|---|---|---|
| Before or after the current GFR threshold to define CKD? | Can chronodisruptors be modified by altering the diet or timing of meals? | Is basic research in CKD tainted by chronodisruption resulting from performing mouse and rat experiments during daytime, which should be their inactive period? |
| Is a decreased GFR needed to trigger CKD-associated chronodisruption? | Or by altering the microbiota? | To what extent the age-associated loss of renal function contributes to age-associated circadian rhythm abnormalities? |
| Or is pathological albuminuria sufficient to trigger chronodisruption? | Or by drugs modulating their signaling pathways? | |
| Does therapeutic targeting of CKD-related chronodisruptors improve outcomes? | ||
| Has melatonin any role in managing CKD? | ||
| Has chronopharmacology a role in CKD? |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carriazo, S.; Ramos, A.M.; Sanz, A.B.; Sanchez-Niño, M.D.; Kanbay, M.; Ortiz, A. Chronodisruption: A Poorly Recognized Feature of CKD. Toxins 2020, 12, 151. https://doi.org/10.3390/toxins12030151
Carriazo S, Ramos AM, Sanz AB, Sanchez-Niño MD, Kanbay M, Ortiz A. Chronodisruption: A Poorly Recognized Feature of CKD. Toxins. 2020; 12(3):151. https://doi.org/10.3390/toxins12030151
Chicago/Turabian StyleCarriazo, Sol, Adrián M Ramos, Ana B Sanz, Maria Dolores Sanchez-Niño, Mehmet Kanbay, and Alberto Ortiz. 2020. "Chronodisruption: A Poorly Recognized Feature of CKD" Toxins 12, no. 3: 151. https://doi.org/10.3390/toxins12030151
APA StyleCarriazo, S., Ramos, A. M., Sanz, A. B., Sanchez-Niño, M. D., Kanbay, M., & Ortiz, A. (2020). Chronodisruption: A Poorly Recognized Feature of CKD. Toxins, 12(3), 151. https://doi.org/10.3390/toxins12030151