A Mini Review on Microcystins and Bacterial Degradation



Abstract
:1. Introduction
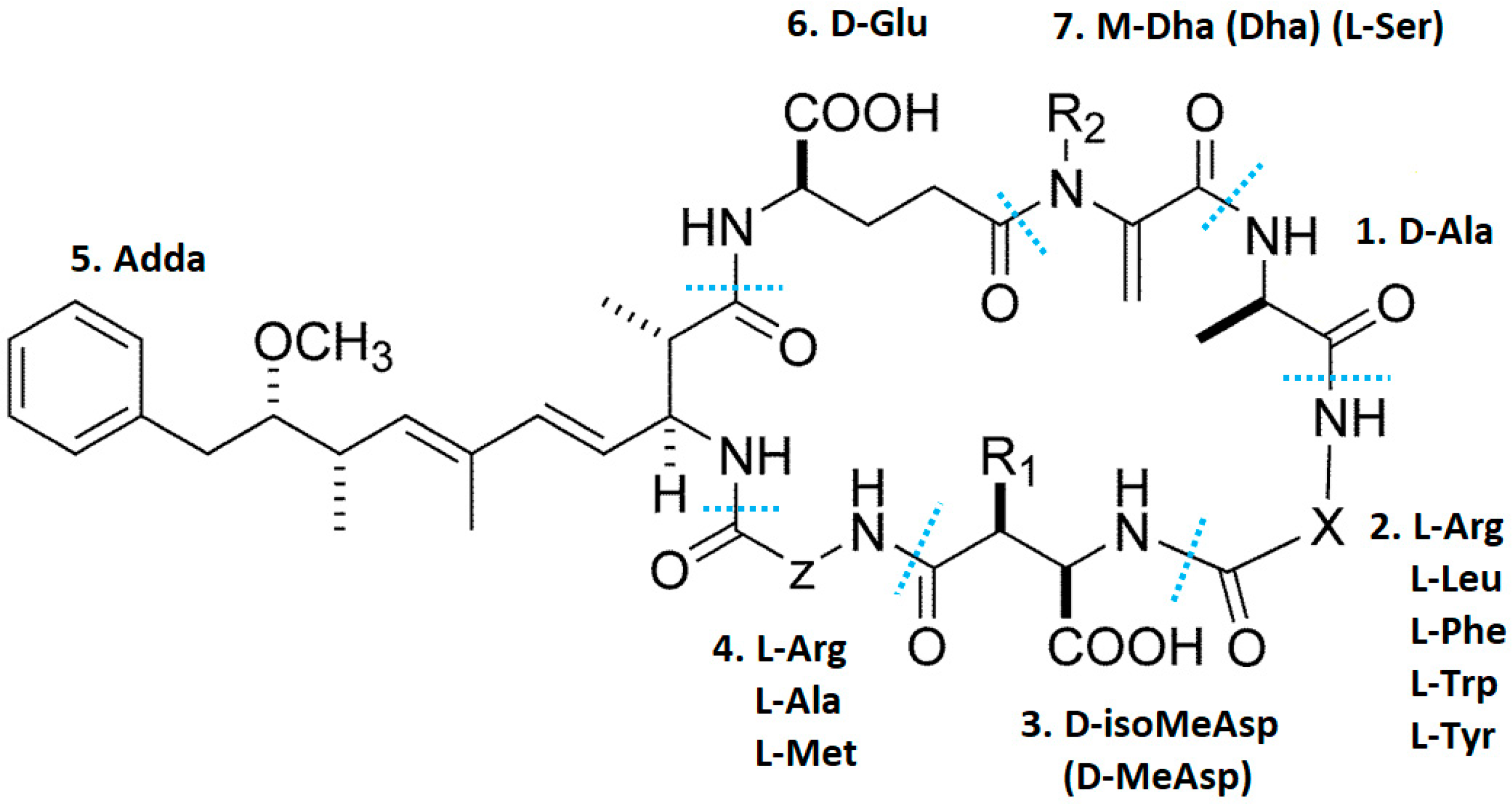
2. Microcystins
2.1. Microcystins Synthesis
2.2. Chemical Properties
2.3. Toxicity and Carcinogenicity
3. Treatment Approach
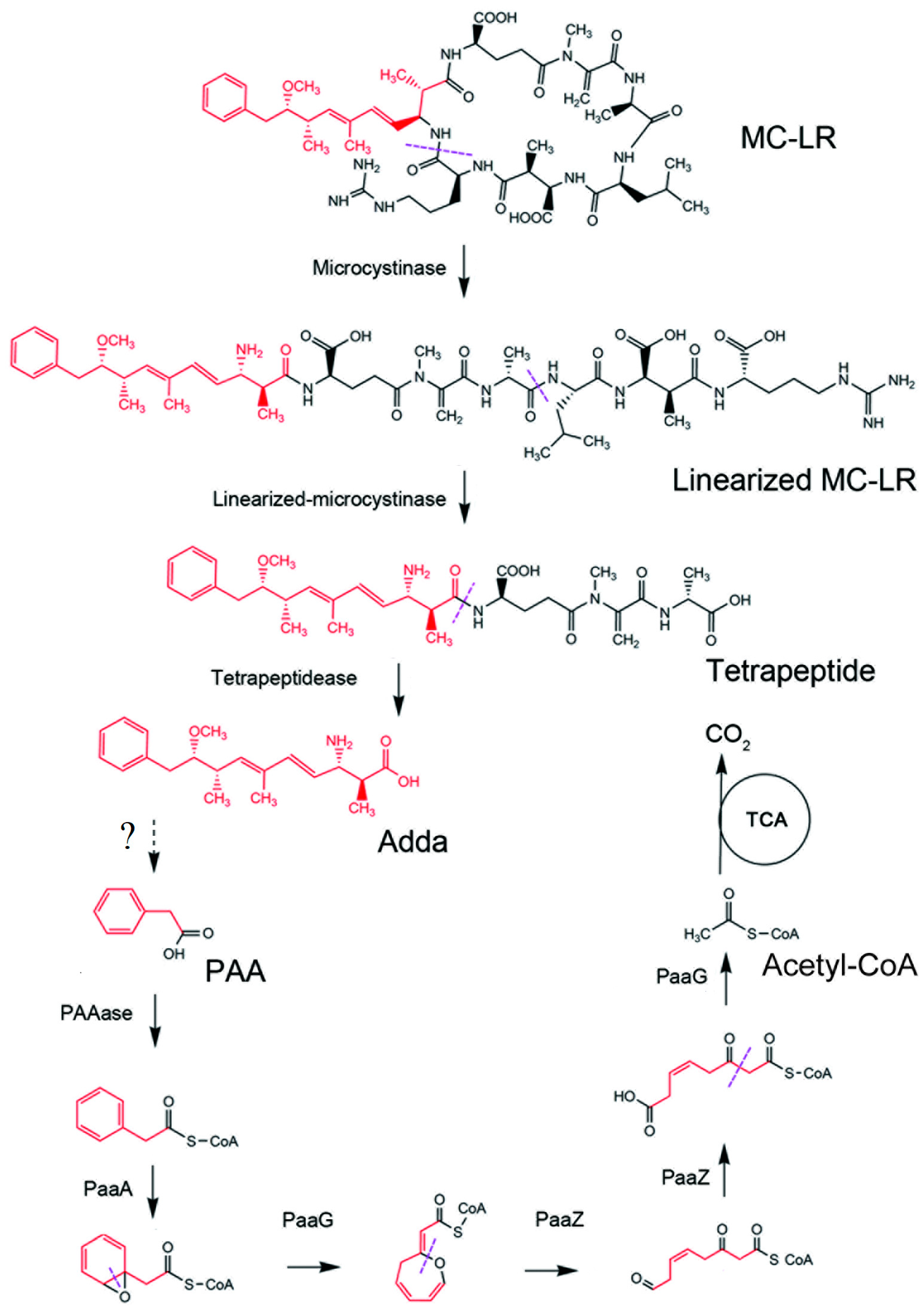
3.1. Biological Degradation by Bacteria
3.2. Enzymatic Mechanisms of Microcystins Biodegradation
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Technical Brief Technical Management of Cyanobacteria in Drinkingwater Supplies: Information for Regulators and Water Suppliers; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Ji, X.; Verspagen, J.M.H.; Stomp, M.; Huisman, J. Competition between cyanobacteria and green algae at low versus elevated CO2: Who will win, and why? J. Exp. Bot. 2017, 68, 3815–3828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.; Chen, W.; Zuo, Y.; Lin, L.; Song, L. Heavy metal migration and risk transference associated with cyanobacterial blooms in eutrophic freshwater. Sci. Total Environ. 2018, 613–614, 1324–1330. [Google Scholar] [CrossRef] [PubMed]
- Silveira, S.B.; Odebrecht, C. Effects of Salinity and temperature on the growth, toxin production, and akinete germination of the cyanobacterium Nodularia spumigena. Front. Mar. Sci. 2019, 6. [Google Scholar] [CrossRef]
- Wang, J.H.; Yang, C.; He, L.Q.; Dao, G.H.; Du, J.S.; Han, Y.P.; Wu, G.X.; Wu, Q.Y.; Hu, H.Y. Meteorological factors and water quality changes of Plateau Lake Dianchi in China (1990–2015) and their joint influences on cyanobacterial blooms. Sci. Total Environ. 2019, 665, 406–418. [Google Scholar] [CrossRef]
- Jankowiak, J.; Hattenrath-Lehmann, T.; Kramer, B.J.; Ladds, M.; Gobler, C.J. Deciphering the effects of nitrogen, phosphorus, and temperature on cyanobacterial bloom intensification, diversity, and toxicity in western Lake Erie. Limnol. Oceanogr. 2019, 64, 1347–1370. [Google Scholar] [CrossRef] [Green Version]
- Drobac, D.; Tokodi, N.; Simeunovic, J.; Baltic, V.; Stanic, D.; Svircev, Z. Human exposure to cyanotoxins and their effects on health. Arh. Hig. Rada Toksikol. 2013, 64, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Sivonen, K.; Jones, G. Cyanobacterial toxins. In Toxic Cyanobacteria in Water. A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; E and FN Spon: London, UK, 1999; pp. 41–111. [Google Scholar]
- Massey, I.Y.; Yang, F.; Ding, Z.; Yang, S.; Guo, J.; Tezi, C.; Al-Osman, M.; Kamegni, R.B.; Zeng, W. Exposure routes and health effects of microcystins on animals and humans: A mini-review. Toxicon 2018, 151, 156–162. [Google Scholar] [CrossRef]
- Zhang, C.; Massey, I.Y.; Liu, Y.; Huang, F.; Gao, R.; Ding, M.; Xiang, L.; He, C.; Wei, J.; Li, Y.; et al. Identification and characterization of a novel indigenous algicidal bacterium Chryseobacterium species against Microcystis aeruginosa. J. Toxicol. Environ. Health Part A 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- IARC. Ingested Nitrate and Nitrite, and Cyanobacterial Peptide Toxins; World Health Organization, International Agency for Research on Cancer: Lyon, France, 2010. [Google Scholar]
- Mantzouki, E.; Lürling, M.; Fastner, J.; de Senerpont Domis, L.; Wilk-Woźniak, E.; Koreivienė, J.; Seelen, L.; Teurlincx, S.; Verstijnen, Y.; Krztoń, W.; et al. Temperature Effects Explain Continental Scale Distribution of Cyanobacterial Toxins. Toxins 2018, 10, 156. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Lee, C. Ozonation of Microcystins: Kinetics and Toxicity Decrease. Environ. Sci. Technol. 2019, 53, 6427–6435. [Google Scholar] [CrossRef]
- Massey, I.Y.; Zhang, X.; Yang, F. Importance of bacterial biodegradation and detoxification processes of microcystins for environmental health. J. Toxicol. Environ. Health Part B 2018, 1–13. [Google Scholar] [CrossRef]
- Sun, J.L.; Bu, L.J.; Chen, S.Y.; Lu, X.L.; Wu, Y.T.; Shi, Z.; Zhou, S.Q. Oxidation of Microcystin-LR via the solar/chlorine process: Radical mechanism, pathways and toxicity assessment. Ecotoxicol. Environ. Saf. 2019, 183. [Google Scholar] [CrossRef]
- Idroos, E.S.; De Silva, B.; Manage, P.M. Biodegradation of microcystin analogues by Stenotrophomonas maltophilia isolated from Beira Lake Sri Lanka. J. Natl. Sci. Found. Sri Lanka 2017, 45, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Maghsoudi, E.; Fortin, N.; Greer, C.; Maynard, C.; Page, A.; Duy, S.V.; Sauve, S.; Prevost, M.; Dorner, S. Cyanotoxin degradation activity and mlr gene expression profiles of a Sphingopyxis sp. isolated from Lake Champlain, Canada. Environ. Sci. Process. Impacts 2016, 18, 1417–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Wei, J.; Yang, K.; Massey, I.Y.; Guo, J.; Zhang, C.; Yang, F. Isolation, molecular identification, and characterization of a unique toxic cyanobacterium Microcystis sp. found in Hunan Province, China. J. Toxicol. Environ. Health Part A 2018, 81, 1142–1149. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Xie, X.; Huang, F.; Xiang, L.; Wang, Y.; Han, T.; Massey, I.Y.; Liang, G.; Pu, Y.; Yang, F. Simultaneous Microcystis algicidal and microcystin synthesis inhibition by a red pigment prodigiosin. Environ. Pollut. 2019, 256, 113444. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Richer, R.; Cox, P.A.; Codd, G.A. Cyanotoxins in desert environments may present a risk to human health. Sci. Total Environ. 2012, 421–422, 118–123. [Google Scholar] [CrossRef]
- Svircev, Z.; Lalic, D.; Bojadzija, S.G.; Tokodi, N.; Drobac, B.D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global geographical and historical overview of cyanotoxin distribution and cyanobacterial poisonings. Arch. Toxicol. 2019. [Google Scholar] [CrossRef]
- Zhang, X.J.; Chen, C.; Ding, J.Q.; Hou, A.; Li, Y.; Niu, Z.B.; Su, X.Y.; Xu, Y.J.; Laws, E.A. The 2007 water crisis in Wuxi, China: Analysis of the origin. J. Hazard. Mater. 2010, 182, 130–135. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, P.; Merkley, N.; Thomas, K.; Lewis, N.I.; Bekri, K.; Renaud, S.L.; Pick, F.R.; McCarron, P.; Miles, C.O.; Quilliam, M.A. Isolation and Characterization of DLeu1 microcystin-LY from Microcystis aeruginosa CPCC-464. Toxins 2020, 12, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, H.; Clara, T.; Huang, F.Y.; Wei, J.; Yang, F. Identification and characterization of the dominant Microcystis sp. cyanobacteria detected in Lake Dong Ting, China. J. Toxicol. Environ. Health Part A Curr. Issues 2019, 82, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Gkelis, S.; Tussy, P.F.; Zaoutsos, N. Isolation and preliminary characterization of cyanobacteria strains from freshwaters of Greece. Open Life Sci. 2015, 10, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Tillett, D.; Dittmann, E.; Erhard, M.; von Dohren, H.; Borner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide-polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Carmichael, W.W. The cyanotoxins. Adv. Bot. Res. 1997, 27, 211–256. [Google Scholar]
- Carmichael, W.W. Cyanobacteria secondary metabolites--the cyanotoxins. J. Appl. Bacteriol. 1992, 72, 445–459. [Google Scholar] [CrossRef]
- Zhang, M.; Pan, G.; Yan, H. Microbial biodegradation of microcystin-RR by bacterium Sphingopyxis sp. USTB-05. J. Environ. Sci. 2010, 22, 168–175. [Google Scholar] [CrossRef]
- Spoof, L.; Catherine, A. Appendix 3 Tables of Microcystins and Nodularins. Handbook of Cyanobacterial Mmonitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 526–537. [Google Scholar] [CrossRef]
- Spoof, L.; Vesterkvist, P.; Lindholm, T.; Meriluoto, J. Screening for cyanobacterial hepatotoxins, microcystins and nodularin in environmental water samples by reversed-phase liquid chromatography-electrospray ionisation mass spectrometry. J. Chromatogr. A 2003, 1020, 105–119. [Google Scholar] [CrossRef]
- Gurbuz, F.; Metcalf, J.S.; Karahan, A.G.; Codd, G.A. Analysis of dissolved microcystins in surface water samples from Kovada Lake, Turkey. Sci. Total Environ. 2009, 407, 4038–4046. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt-Oliveira, M.C.; Oliveira, M.C.; Pinto, E. Diversity of microcystin-producing genotypes in Brazilian strains of Microcystis (Cyanobacteria). Braz. J. Biol. 2011, 71, 209–216. [Google Scholar] [CrossRef]
- Cao, L.; Massey, I.Y.; Feng, H.; Yang, F. A Review of Cardiovascular Toxicity of Microcystins. Toxins 2019, 11, 507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, K.; Naito, S.; Kondo, F.; Ishikawa, N.; Watanabe, M.F.; Suzuki, M.; Harada, K. Stability of microcystins from cyanobacteria: Effect of light on decomposition and isomerization. Environ. Sci. Technol. 2004, 28, 173–177. [Google Scholar] [CrossRef] [PubMed]
- De Figueiredo, D.R.; Azeiteiro, U.M.; Esteves, S.M.; Goncalves, F.J.; Pereira, M.J. Microcystin-producing blooms-a serious global public health issue. Ecotoxicol. Environ. Saf. 2004, 59, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Tsuji, K.; Watanabe, M.F.; Kondo, F. Stability of microcystins from cyanobacteria—III. Effect of pH and temperature. Phycologia 1996, 35, 83–88. [Google Scholar] [CrossRef]
- Donati, C.; Drikas, M.; Hayes, R.; Newcombe, G. Microcystin-LR adsorption by powdered activated carbon. Water Res. 1994, 28, 1735–1742. [Google Scholar] [CrossRef]
- Vesterkvist, P.S.M.; Misiorek, J.O.; Spoof, L.E.M.; Toivola, D.M.; Meriluoto, J.A.O. Comparative Cellular Toxicity of Hydrophilic and Hydrophobic Microcystins on Caco-2 Cells. Toxins 2012, 4, 1008–1023. [Google Scholar] [CrossRef] [Green Version]
- Feitz, A.J.; Waite, T.D.; Jones, G.J.; Boyden, B.H.; Orr, P.T. Photocatalytic degradation of the blue green algal toxin microcystin-LR in a natural organic-aqueous matrix. Environ. Sci. Technol. 1999, 33, 243–249. [Google Scholar] [CrossRef]
- Cao, L.; Huang, F.; Massey, I.Y.; Wen, C.; Zheng, S.; Xu, S.; Yang, F. Effects of Microcystin-LR on the Microstructure and Inflammation-Related Factors of Jejunum in Mice. Toxins 2019, 11, 482. [Google Scholar] [CrossRef] [Green Version]
- Alosman, M.; Cao, L.H.; Massey, I.Y.; Yang, F. The lethal effects and determinants of microcystin-LR on heart: A mini review. Toxin Rev. 2020. [Google Scholar] [CrossRef]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Dawson, R.M. The toxicology of microcystins. Toxicon 1998, 36, 953–962. [Google Scholar] [CrossRef]
- Zhou, M.; Tu, W.; Xu, J. Mechanisms of microcystin-LR-induced cytoskeletal disruption in animal cells. Toxicon 2015, 101, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sun, Y. The role of PP2A-associated proteins and signal pathways in microcystin-LR toxicity. Toxicol. Lett. 2015, 236, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mrdjen, I.; Morse, M.A.; Ruch, R.J.; Knobloch, T.J.; Choudhary, S.; Weghorst, C.M.; Lee, J. Impact of Microcystin-LR on Liver Function Varies by Dose and Sex in Mice. Toxins 2018, 10, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calado, S.L.D.; Vicentini, M.; Santos, G.S.; Pelanda, A.; Santos, H.; Coral, L.A.; Magalhaes, V.D.; Mela, M.; Cestari, M.M.; de Assis, H.C.S. Sublethal effects of microcystin-LR in the exposure and depuration time in a neotropical fish: Multibiomarker approach. Ecotoxicol. Environ. Saf. 2019, 183. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yan, Y.; Xie, L.; Wang, L.; He, Y.; Wan, X.; Xue, Q. Long-term environmental exposure to microcystins increases the risk of nonalcoholic fatty liver disease in humans: A combined fisher-based investigation and murine model study. Environ. Int. 2020, 138, 105648. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xie, P. Mechanisms of microcystin-induced cytotoxicity and apoptosis. Mini Rev. Med. Chem. 2016, 16, 1018–1031. [Google Scholar] [CrossRef]
- Wang, C.; Gu, S.; Yin, X.; Yuan, M.; Xiang, Z.; Li, Z.; Cao, H.; Meng, X.; Hu, K.; Han, X. The toxic effects of microcystin-LR on mouse lungs and alveolar type II epithelial cells. Toxicon 2016, 115, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.F.; Xu, L.Z.; Li, X.X.; Chen, J.W.; Zhou, W.; Sun, J.P.; Wang, Y.P. The differential effects of microcystin-LR on mitochondrial DNA in the hippocampus and cerebral cortex. Environ. Pollut. 2018, 240, 68–76. [Google Scholar] [CrossRef]
- Yi, X.P.; Xu, S.S.; Huang, F.Y.; Wen, C.; Zheng, S.L.; Feng, H.; Guo, J.; Chen, J.H.; Feng, X.L.; Yang, F. Effects of chronic exposure to microcystin-LR on kidney in mice. Int. J. Environ. Res. Public Health 2019, 16, 5030. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Y.; Zhuang, H.; Yang, H.; Xue, W.; Wang, L.F.; Wei, W.Z. Microcystin-LR disturbs testicular development of giant freshwater prawn Macrobrachium rosenbergii. Chemosphere 2019, 222, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.L.; Liu, Y.L.; Guo, J.S.; Lin, S.; Wang, Y.Q.; Yin, T.Y.; Gregersen, H.; Hu, T.Z.; Wang, G.X. Microcystin-LR induces angiodysplasia and vascular dysfunction through promoting cell apoptosis by the mitochondrial signaling pathway. Chemosphere 2019, 218, 438–448. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sinha, R.P.; Incharoensakdi, A. The cyanotoxin-microcystins: Current overview. Rev. Environ. Sci. Bio Technol. 2014, 13, 215–249. [Google Scholar] [CrossRef]
- Jochimsen, E.M.; Carmichael, W.W.; An, J.S.; Cardo, D.M.; Cookson, S.T.; Holmes, C.E.; Antunes, M.B.; de Melo Filho, D.A.; Lyra, T.M.; Barreto, V.S.; et al. Liver failure and death after exposure to microcystins at a hemodialysis center in Brazil. N. Engl. J. Med. 1998, 338, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, Z.; Ahmed, Z.; Bakr, A.; Hashem, M.; Alamri, S. Detection of free and bound microcystins in tilapia fish from Egyptian fishpond farms and its related public health risk assessment. J. Consum. Prot. Food Saf. 2020, 15, 37–47. [Google Scholar] [CrossRef]
- Greer, B.; Maul, R.; Campbell, K.; Elliott, C. Detection of freshwater cyanotoxins and measurement of masked microcystins in tilapia from Southeast Asian aquaculture farms. Anal. Bioanal. Chem. 2017, 409, 4057–4069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, C.; Wang, W. Response and recovery of rice (Oryza sativa) seedlings to irrigation with microcystin-contaminated water. Environ. Earth Sci. 2015, 73, 4573–4580. [Google Scholar] [CrossRef]
- Drobac, D.; Tokodi, N.; Kiprovski, B.; Malencic, D.; Vazic, T.; Nybom, S.; Meriluoto, J.; Svircev, Z. Microcystin accumulation and potential effects on antioxidant capacity of leaves and fruits of Capsicum annuum. J. Toxicol. Environ. Health Part A Curr. Issues 2017, 80, 145–154. [Google Scholar] [CrossRef]
- Levizou, E.; Statiris, G.; Papadimitriou, T.; Laspidou, C.S.; Kormas, K.A. Lettuce facing microcystins-rich irrigation water at different developmental stages: Effects on plant performance and microcystins bioaccumulation. Ecotoxicol. Environ. Saf. 2017, 143, 193–200. [Google Scholar] [CrossRef]
- Zhu, J.Z.; Ren, X.Q.; Liu, H.Y.; Liang, C.J. Effect of irrigation with microcystins-contaminated water on growth and fruit quality of Cucumis sativus L. and the health risk. Agric. Water Manag. 2018, 204, 91–99. [Google Scholar] [CrossRef]
- Xiang, L.; Li, Y.W.; Liu, B.L.; Zhao, H.M.; Li, H.; Cai, Q.Y.; Mo, C.H.; Wong, M.H.; Li, Q.X. High ecological and human health risks from microcystins in vegetable fields in southern China. Environ. Int. 2019, 133. [Google Scholar] [CrossRef] [PubMed]
- WHO. Cyanobacterial Toxins: Microcystin-LR. Guidelines for Drinking Water Quality; World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- WHO. Coastal and Fresh Waters, Guidelines for Safe Recreational Water Environments; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- He, X.; Liu, Y.L.; Conklin, A.; Westrick, J.; Weavers, L.K.; Dionysiou, D.D.; Lenhart, J.J.; Mouser, P.J.; Szlag, D.; Walker, H.W. Toxic cyanobacteria and drinking water: Impacts, detection, and treatment. Harmful Algae 2016, 54, 174–193. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.C.B.; Lee, A.K.; Yates, R.S.; Liang, S.; Rochelle, P.A. Analysis of Microcystins in Drinking Water by ELISA and LC/MS/MS. J. Am. Water Works Assoc. 2017, 109, 13–25. [Google Scholar] [CrossRef]
- Jasa, L.; Sadilek, J.; Kohoutek, J.; Strakova, L.; Marsalek, B.; Babica, P. Application of passive sampling for sensitive time-integrative monitoring of cyanobacterial toxins microcystins in drinking water treatment plants. Water Res. 2019, 153, 108–120. [Google Scholar] [CrossRef]
- Szlag, D.C.; Sinclair, J.L.; Southwell, B.; Westrick, J.A. Cyanobacteria and Cyanotoxins Occurrence and Removal from Five High-Risk Conventional Treatment Drinking Water Plants. Toxins 2015, 7, 2198–2220. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.Y.; Feng, H.; Li, X.Y.; Yi, X.P.; Guo, J.; Clara, T.; Yang, F. Anaerobic degradation of microcystin-LR by an indigenous bacterial Enterobacter sp. YF3. J. Toxicol. Environ. Health Part A Curr. Issues 2019. [Google Scholar] [CrossRef]
- Zhu, X.Y.; Shen, Y.T.; Chen, X.G.; Hu, Y.O.O.; Xiang, H.Y.; Tao, J.; Ling, Y. Biodegradation mechanism of microcystin-LR by a novel isolate of Rhizobium sp. TH and the evolutionary origin of the mlrA gene. Int. Biodeterior. Biodegrad. 2016, 115, 17–25. [Google Scholar] [CrossRef]
- Krishnan, A.; Zhang, Y.Q.; Mou, X.Z. Isolation and Characterization of Microcystin-Degrading Bacteria from Lake Erie. Bull. Environ. Contam. Toxicol. 2018, 101, 617–623. [Google Scholar] [CrossRef]
- Lemes, G.A.; Kist, L.W.; Bogo, M.R.; Yunes, J.S. Biodegradation of [D-Leu(1)] microcystin-LR by a bacterium isolated from sediment of Patos Lagoon estuary, Brazil. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Lu, Q.; Ding, Q.; Yin, L.; Pu, Y. A Novel and Native Microcystin-Degrading Bacterium of Sphingopyxis sp. Isolated from Lake Taihu. Int. J. Environ. Res. Public Health 2017, 14, 1187. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.J.; Bourne, D.G.; Blakeley, R.L.; Doelle, H. Degradation of the cyanobacterial hepatotoxin microcystin by aquatic bacteria. Nat. Toxins 1994, 2, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ai, H.; Kang, L.; Sun, X.; He, Q. Simultaneous Microcystis Algicidal and Microcystin Degrading Capability by a Single Acinetobacter Bacterial Strain. Environ. Sci. Technol. 2016, 50, 11903–11911. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Pan, G. Enhanced and continued degradation of microcystins using microorganisms obtained through natural media. J. Microbiol. Methods 2014, 96, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Mankiewicz-Boczek, J.; Gagala, I.; Jurczak, T.; Jaskulska, A.; Pawelczyk, J.; Dziadek, J. Bacteria homologus to Aeromonas capable of microcystin degradation. Open Life Sci. 2015, 10, 119–129. [Google Scholar] [CrossRef]
- Lawton, L.A.; Welgamage, A.; Manage, P.M.; Edwards, C. Novel bacterial strains for the removal of microcystins from drinking water. Water Sci. Technol. 2011, 63, 1137–1142. [Google Scholar] [CrossRef]
- Manage, P.M.; Edwards, C.; Singh, B.K.; Lawton, L.A. Isolation and identification of novel microcystin-degrading bacteria. Appl. Environ. Microbiol. 2009, 75, 6924–6928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alamri, S.A. Biodegradation of microcystin by a new Bacillus sp. isolated from a Saudi freshwater lake. Afr. J. Biotechnol. 2010, 9, 6552–6559. [Google Scholar]
- Hu, L.; Zhang, F.; Liu, C.; Wang, M. Biodegradation of Microcystins by Bacillus sp. strain EMB. Energy Procedia 2012, 16, 2054–2059. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Shi, H.; Liu, A.; Cao, Z.; Hao, J.; Gong, R. Identification of a new microcystin-degrading bacterium isolated from Lake Chaohu, China. Bull. Environ. Contam. Toxicol. 2015, 94, 661–666. [Google Scholar] [CrossRef]
- Alamri, S.A. Biodegradation of microcystin-RR by Bacillus flexus isolated from a Saudi freshwater lake. Saudi J. Biol. Sci. 2012, 19, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Kansole, M.M.R.; Lin, T.F. Microcystin-LR Biodegradation by Bacillus sp.: Reaction Rates and Possible Genes Involved in the Degradation. Water 2016, 8, 508. [Google Scholar] [CrossRef] [Green Version]
- Nybom, S.M.K.; Salminen, S.J.; Meriluoto, J.A.O. Removal of microcystin-LR by strains of metabolically active probiotic bacteria. FEMS Microbiol. Lett. 2007, 270, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Zhou, Y.; Sun, R.; Wei, H.; Li, Y.; Yin, L.; Pu, Y. Biodegradation of microcystin-LR and-RR by a novel microcystin-degrading bacterium isolated from Lake Taihu. Biodegradation 2014, 25, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Lemes, G.A.; Kersanach, R.; Pinto, L.D.S.; Dellagostin, O.A.; Yunes, J.S.; Matthiensen, A. Biodegradation of microcystins by aquatic Burkholderia sp. from a South Brazilian coastal lagoon. Ecotoxicol. Environ. Saf. 2008, 69, 358–365. [Google Scholar] [CrossRef]
- Shen, R.Y.; Chen, Z.H.; Dong, X.N.; Shen, H.C.; Su, P.; Mao, L.Q.; Zhang, W.Y. Biodegradation kinetics of microcystins-LR crude extract by Lysinibacillus boronitolerans strain CQ5. Ann. Microbiol. 2019, 69, 1259–1266. [Google Scholar] [CrossRef]
- Hu, L.B.; Yang, J.D.; Zhou, W.; Yin, Y.F.; Chen, J.; Shi, Z.Q. Isolation of a Methylobacillus sp. that degrades microcystin toxins associated with cyanobacteria. New Biotechnol. 2009, 26, 205–211. [Google Scholar] [CrossRef]
- Eleuterio, L.; Batista, J.R. Biodegradation studies and sequencing of microcystin-LR degrading bacteria isolated from a drinking water biofilter and a fresh water lake. Toxicon 2010, 55, 1434–1442. [Google Scholar] [CrossRef]
- Phujomjai, Y.; Somdee, T. Isolation and characterization of microcystin-degrading bacterium. J. Life Sci. Technol. 2013, 1, 220–222. [Google Scholar] [CrossRef]
- Somdee, T.; Wibuloutai, J.; Somdee, T.; Somdee, A. Biodegradation of the cyanobacterial hepatotoxin Dha(7) microcystin-LR within a biologically active sand filter. Water Sci. Technol. Water Supply 2014, 14, 672–680. [Google Scholar] [CrossRef]
- Phujomjai, Y.; Somdee, A.; Somdee, T. Biodegradation of microcystin Dha(7) MC-LR by a novel microcystin-degrading bacterium in an internal airlift loop bioreactor. Water Sci. Technol. 2016, 73, 267–274. [Google Scholar] [CrossRef]
- Jiang, Y.; Shao, J.; Wu, X.; Xu, Y.; Li, R. Active and silent members in the mlr gene cluster of a microcystin-degrading bacterium isolated from Lake Taihu, China. FEMS Microbiol. Lett. 2011, 322, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Jing, W.; Sui, G.; Liu, S. Characteristics of a Microcystin-LR Biodegrading Bacterial Isolate: Ochrobactrum sp. FDT5. Bull. Environ. Contam. Toxicol. 2014, 92, 119–122. [Google Scholar] [CrossRef] [PubMed]
- You, D.J.; Chen, X.G.; Xiang, H.Y.; Ouyang, L.; Yang, B. Isolation, identification and characterization of a microcystin-degrading bacterium Paucibacter sp. strain CH. Huan Jing Ke Xue 2014, 35, 313–318. [Google Scholar] [PubMed]
- Lezcano, M.A.; Moron-Lopez, J.; Agha, R.; Lopez-Heras, I.; Nozal, L.; Quesada, A.; El-Shehawy, R. Presence or Absence of mlr Genes and Nutrient Concentrations Co-Determine the Microcystin Biodegradation Efficiency of a Natural Bacterial Community. Toxins 2016, 8, 318. [Google Scholar] [CrossRef] [PubMed]
- Rapala, J.; Berg, K.A.; Lyra, C.; Niemi, R.M.; Manz, W.; Suomalainen, S.; Paulin, L.; Lahti, K. Paucibacter toxinivorans gen. nov., sp. nov., a bacterium that degrades cyclic cyanobacterial hepatotoxins microcystins and nodularin. Int. J. Syst. Evol. Microbiol. 2005, 55, 1563–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenaka, S.; Watanabe, M.F. Microcystin LR degradation by Pseudomonas aeruginosa alkaline protease. Chemosphere 1997, 34, 749–757. [Google Scholar] [CrossRef]
- Zhang, M.L.; Yan, H.; Pan, G. Microbial degradation of microcystin-LR by Ralstonia solanacearum. Environ. Technol. 2011, 32, 1779–1787. [Google Scholar] [CrossRef]
- Saito, T.; Okano, K.; Park, H.D.; Itayama, T.; Inamori, Y.; Neilan, B.A.; Burns, B.P.; Sugiura, N. Detection and sequencing of the microcystin LR-degrading gene, mlrA, from new bacteria isolated from Japanese lakes. FEMS Microbiol. Lett. 2003, 229, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Bourne, D.G.; Jones, G.J.; Blakeley, R.L.; Jones, A.; Negri, A.P.; Riddles, P. Enzymatic pathway for the bacterial degradation of the cyanobacterial cyclic peptide toxin microcystin LR. Appl. Environ. Microbiol. 1996, 62, 4086–4094. [Google Scholar] [CrossRef] [Green Version]
- Imanishi, S.; Kato, H.; Mizuno, M.; Tsuji, K.; Harada, K. Bacterial degradation of microcystins and nodularin. Chem. Res. Toxicol. 2005, 18, 591–598. [Google Scholar] [CrossRef]
- Harada, K.; Imanishi, S.; Kato, H.; Mizuno, M.; Ito, E.; Tsuji, K. Isolation of Adda from microcystin-LR by microbial degradation. Toxicon 2004, 44, 107–109. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Imanishi, S.Y.; Tsuji, K.; Harada, K. Microbial degradation of cyanobacterial cyclic peptides. Water Res. 2007, 41, 1754–1762. [Google Scholar] [CrossRef] [PubMed]
- Valeria, A.M.; Ricardo, E.J.; Stephan, P.; Alberto, W.D. Degradation of Microcystin-RR by Sphingomonas sp. CBA4 isolated from San Roque reservoir (Cordoba—Argentina). Biodegradation 2006, 17, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, T.; Park, H.D.; Ozawa, K.; Tanaka, Y.; Sumino, T.; Hamana, K.; Hiraishi, A.; Kato, K. Sphingosinicella microcystinivorans gen. nov., sp nov., a microcystin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2006, 56, 85–89. [Google Scholar] [CrossRef]
- Park, H.D.; Sasaki, Y.; Maruyama, T.; Yanagisawa, E.; Hiraishi, A.; Kato, K. Degradation of the cyanobacterial hepatotoxin microcystin by a new bacterium isolated from a hypertrophic lake. Environ. Toxicol. 2001, 16, 337–343. [Google Scholar] [CrossRef]
- Ishii, H.; Nishijima, M.; Abe, T. Characterization of degradation process of cyanobacterial hepatotoxins by a gram-negative aerobic bacterium. Water Res. 2004, 38, 2667–2676. [Google Scholar] [CrossRef]
- Okano, K.; Shimizu, K.; Kawauchi, Y.; Maseda, H.; Utsumi, M.; Zhang, Z.; Neilan, B.A.; Sugiura, N. Characteristics of a Microcystin-Degrading Bacterium under Alkaline Environmental Conditions. J. Toxicol. 2009, 2009, 954291. [Google Scholar] [CrossRef]
- Shimizu, K.; Maseda, H.; Okano, K.; Kurashima, T.; Kawauchi, Y.; Xue, Q.; Utsumi, M.; Zhang, Z.; Sugiura, N. Enzymatic pathway for biodegrading microcystin LR in Sphingopyxis sp. C-1. J. Biosci. Bioeng. 2012, 114, 630–634. [Google Scholar] [CrossRef]
- Ho, L.; Hoefel, D.; Saint, C.P.; Newcombe, G. Isolation and identification of a novel microcystin-degrading bacterium from a biological sand filter. Water Res. 2007, 41, 4685–4695. [Google Scholar] [CrossRef]
- Ding, Q.; Liu, K.Y.; Xu, K.; Sun, R.L.; Zhang, J.; Yin, L.H.; Pu, Y.P. Further Understanding of Degradation Pathways of Microcystin-LR by an Indigenous Sphingopyxis sp. in Environmentally Relevant Pollution Concentrations. Toxins 2018, 10, 536. [Google Scholar] [CrossRef] [Green Version]
- Ho, L.; Tang, T.; Monis, P.T.; Hoefel, D. Biodegradation of multiple cyanobacterial metabolites in drinking water supplies. Chemosphere 2012, 87, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, P.; Chen, J.; Yan, H. Biodegradation of Microcystin-RR by a New Isolated Sphingopyxis sp. USTB-05. Chin. J. Chem. Eng. 2010, 18, 108–112. [Google Scholar] [CrossRef]
- Xiao, W.; Yan, W.; Jian, O.; Wei, N.; Pan, G. Microcystin-LR biodegradation by Sphingopyxis sp. USTB-05. Front. Environ. Sci. Eng. China 2011, 5, 526–532. [Google Scholar] [CrossRef]
- Xu, H.; Wang, H.; Xu, Q.; Lv, L.; Yin, C.; Liu, X.; Du, H.; Yan, H. Pathway for Biodegrading Microcystin-YR by Sphingopyxis sp. USTB-05. PLoS ONE 2015, 10, e0124425. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Wang, H.; Wang, J.; Yin, C.; Ma, S.; Liu, X.; Yin, X. Cloning and expression of the first gene for biodegrading microcystin LR by Sphingopyxis sp. USTB-05. J. Environ. Sci. 2012, 24, 1816–1822. [Google Scholar] [CrossRef]
- Qin, L.; Zhang, X.X.; Chen, X.G.; Wang, K.; Shen, Y.T.; Li, D. Isolation of a Novel Microcystin-Degrading Bacterium and the Evolutionary Origin of mlr Gene Cluster. Toxins 2019, 11, 269. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Huang, F.; Feng, H.; Wei, J.; Massey, I.Y.; Liang, G.; Zhang, F.; Yin, L.; Kacew, S.; Zhang, X.; et al. A complete route for biodegradation of potentially carcinogenic cyanotoxin microcystin-LR in a novel indigenous bacterium. Water Res. 2020, 174, 115638. [Google Scholar] [CrossRef]
- Chen, J.; Hu, L.B.; Zhou, W.; Yan, S.H.; Yang, J.D.; Xue, Y.F.; Shi, Z.Q. Degradation of microcystin-LR and RR by a Stenotrophomonas sp. strain EMS isolated from Lake Taihu, China. Int. J. Mol. Sci. 2010, 11, 896–911. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Zhou, Y.; Yin, L.; Zhu, G.; Liang, G.; Pu, Y. Microcystin-degrading activity of an indigenous bacterial strain Stenotrophomonas acidaminiphila MC-LTH2 isolated from Lake Taihu. PLoS ONE 2014, 9, e86216. [Google Scholar] [CrossRef] [Green Version]
- Manheim, D.; Cheung, Y.M.; Jiang, S. The Effect of Organic Carbon Addition on the Community Structure and Kinetics of Microcystin-Degrading Bacterial Consortia. Water 2018, 10, 1523. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Hegde, K.; Brar, S.K.; Cledon, M.; Kermanshahi-Pour, A.; Roy-Lachapelle, A.; Galvez-Cloutier, R. Biodegradation of microcystin-LR using acclimatized bacteria isolated from different units of the drinking water treatment plant. Environ. Pollut. 2018, 242, 407–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Guo, J.; Huang, F.; Massey, I.Y.; Huang, R.; Li, Y.; Wen, C.; Ding, P.; Zeng, W.; Liang, G. Removal of Microcystin-LR by a Novel Native Effective Bacterial Community Designated as YFMCD4 Isolated from Lake Taihu. Toxins 2018, 10, 363. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Massey, I.Y.; Guo, J.; Yang, S.; Pu, Y.; Zeng, W.; Tan, H. Microcystin-LR degradation utilizing a novel effective indigenous bacterial community YFMCD1 from Lake Taihu. J. Toxicol. Environ. Health A 2018, 1–10. [Google Scholar] [CrossRef]
- Tsao, S.; Wei, D.J.; Chang, Y.T.; Lee, J.F. Aerobic biodegradation biodegradation of microcystin-LR by an indigenous bacterial mixed culture isolated in Taiwan. Int. Biodeterior. Biodegrad. 2017, 124, 101–108. [Google Scholar] [CrossRef]
- Briand, E.; Humbert, J.F.; Tambosco, K.; Bormans, M.; Gerwick, W.H. Role of bacteria in the production and degradation of Microcystis cyanopeptides. Microbiologyopen 2016, 5, 469–478. [Google Scholar] [CrossRef]
- Mou, X.; Lu, X.; Jacob, J.; Sun, S.; Heath, R. Metagenomic Identification of Bacterioplankton Taxa and Pathways Involved in Microcystin Degradation in Lake Erie. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramani, A.; Rein, K.; Shetty, K.G.; Jayachandran, K. Microbial degradation of microcystin in Florida’s freshwaters. Biodegradation 2012, 23, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Christoffersen, K.; Lyck, S.; Winding, A. Microbial activity and bacterial community structure during degradation of microcystins. Aquat. Microb. Ecol. 2002, 27, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Bourne, D.G.; Riddles, P.; Jones, G.J.; Smith, W.; Blakeley, R.L. Characterisation of a gene cluster involved in bacterial degradation of the cyanobacterial toxin microcystin LR. Environ. Toxicol. 2001, 16, 523–534. [Google Scholar] [CrossRef]
- Dziga, D.; Wasylewski, M.; Szetela, A.; Bocheńska, O.; Wladyka, B. Verification of the Role of MlrC in Microcystin Biodegradation by Studies Using a Heterologously Expressed figureEnzyme. Chem. Res. Toxicol. 2012, 25, 1192–1194. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Strain | mlr Gene | Degradable MC Variant | Class Affiliated | Reference |
|---|---|---|---|---|---|
| Acidovorax facilis | LEw-2 | MC-LR | β-proteobacteria | [74] | |
| Acinetobacter sp. | CMDB-2 | MC-LR | γ-proteobacteria | [78] | |
| WC-5 | MC-LR, MC-RR | γ-proteobacteria | [79] | ||
| Aeromonas sp. | MC-LR | γ-proteobacteria | [80] | ||
| Arthrobacter sp. | C6 | MC-LR, MC-LW, MC-RR, MC-LF, MC-LY | Actinobacteria | [81,82] | |
| F7 | MC-LR, MC-LW, MC-RR, MC-LF, MC-LY | Actinobacteria | [81,82] | ||
| F10 | MC-LR | Actinobacteria | [82] | ||
| R1 | MC-LR | Actinobacteria | [82] | ||
| R4 | MC-LR, MC-LW, MC-RR, MC-LF, MC-LY | Actinobacteria | [81,82] | ||
| R6 | MC-LR | Actinobacteria | [82] | ||
| R9 | MC-LR | Actinobacteria | [82] | ||
| Bacillus sp. | AMRI-03 | mlrA | MC-RR | Bacilli | [83] |
| EMB | mlrA | MC-LR, MC-RR | Bacilli | [84] | |
| JZ-2013 | MC-LR | Bacilli | [85] | ||
| LEw-2010 | MC-LR | Bacilli | [74] | ||
| SSZ01 | mlrA | MC-RR | Bacilli | [86] | |
| MC-LR | Bacilli | [87] | |||
| Bifidobacterium longum | Bb12 | MC-LR | Actinobacteria | [88] | |
| 46 | MC-LR | Actinobacteria | [88] | ||
| 420 | MC-LR | Actinobacteria | [88] | ||
| Bordetella sp. | MC-LTH1 | mlrA | MC-LR, MC-RR | β-proteobacteria | [89] |
| Brevibacillus brevis | LEw-1238 | MC-LR | Bacilli | [74] | |
| Brevibacterium sp. | F3 | MC-LR, MC-LW, MC-RR, MC-LF, MC-LY | Actinobacteria | [81] | |
| Burkholderia sp. | MC-LR, [D-Leu1]MC–LR | β-proteobacteria | [90] | ||
| Enterobacter sp. | YF3 | MC-LR | γ-proteobacteria | [72] | |
| Lactobacillus rhamnosus | GG | MC-LR | Bacilli | [88] | |
| LC-705 | MC-LR | Bacilli | [88] | ||
| Lysinibacillus boronitolerans | CQ5 | MC-LR | Bacilli | [91] | |
| Methylobacillus sp. | J10 | MC-LR, MC-RR | β-proteobacteria | [92] | |
| Morganella morganii | C25216 | MC-LR | Actinobacteria | [93] | |
| C25217 | MC-LR | Actinobacteria | [93] | ||
| C25220 | MC-LR | Actinobacteria | [93] | ||
| Novosphingobium sp. | KKU-12 | [Dha7] MC-LR | α-proteobacteria | [94] | |
| KKU15 | [Dha7] MC-LR | α-proteobacteria | [95] | ||
| KKU-25s | mlrABCD | [Dha7] MC-LR | α-proteobacteria | [96] | |
| THN-1 | mlrABCD | MC-LR | α-proteobacteria | [97] | |
| Ochrobactrum sp. | FDT5 | MC-LR | α-proteobacteria | [98] | |
| Paucibacter sp. | CH | MC-LR | β-proteobacteria | [99] | |
| IM-4 | MC-LR, MC-RR, MC-YR | β-proteobacteria | [100] | ||
| 2C20T | MC-LR, MC-RR, MC-YR | β-proteobacteria | [101] | ||
| Pseudomonas sp. | DMXS | [D-Leu1] MC-LR | γ-proteobacteria | [75] | |
| LEw-1033 | MC-LR | γ-proteobacteria | [74] | ||
| LEw-2166 | MC-LR | γ-proteobacteria | [74] | ||
| Pseudomonas aeruginosa | MC-LR | γ-proteobacteria | [102] | ||
| WC-4 | MC-LR, MC-RR | γ-proteobacteria | [79] | ||
| Ralstonia solanacearum | MC-LR, MC-RR | β-proteobacteria | [103] | ||
| Rhizobium sp. | TH | mlrABCD | MC-LR | α-proteobacteria | [73] |
| Rhodococcus sp. | C1 | MC-LR, MC-LW, MC-RR, MC-LF, MC-LY | Actinobacteria | [81] | |
| C3 | MC-LR | Actinobacteria | [81,82] | ||
| Sphingomonas sp. | ACM-3962/MJ-PV | mlrABCD | MC-LR, MC-RR | α-proteobacteria | [77,104,105] |
| B9 | mlrA | MC-LR, MC-RR, 3-DMMCLR, DHMCLR, MCLR-Cys | α-proteobacteria | [106,107,108] | |
| CBA4 | MC-RR | α-proteobacteria | [109] | ||
| MD-1 | mlrABCD | MC-LR, MC-RR, MC-YR | α-proteobacteria | [104] | |
| MDB2 | α-proteobacteria | [110] | |||
| MDB3 | α-proteobacteria | [110] | |||
| Y2 | mlrA | MC-LR, MC-RR, MC-YR | α-proteobacteria | [111] | |
| 7CY | MC-LR, MC-LW, MC-RR, MC-LF, MC-YR, MC-LY | α-proteobacteria | [112] | ||
| Sphingopyxis sp. | a7 | mlrACD | MC-LR | α-proteobacteria | [76] |
| C1 | mlrABC | MC-LR | α-proteobacteria | [113,114] | |
| IM-1 | mlrABCD | MC-LR, MC-RR, MC-YR | α-proteobacteria | [100] | |
| IM-2 | mlrABCD | MC-LR, MC-RR, MC-YR | α-proteobacteria | [100] | |
| IM-3 | mlrABCD | MC-LR, MC-RR, MC-YR | α-proteobacteria | [100] | |
| LH21 | mlrABCD | MC-LR, MC-LA | α-proteobacteria | [115] | |
| m6 | mlrABCD | MC-LR | α-proteobacteria | [116] | |
| MB-E | mlrABCD | MC-LR, MC-LW, MC-YR, MC-LY, MC-LF | α-proteobacteria | [17] | |
| TT25 | mlrA | MC-LR, MC-RR, MC-YR | α-proteobacteria | [117] | |
| USTB05 | mlrABCD | MC-LR, MC-RR, MC-YR | α-proteobacteria | [30,118,119,120,121] | |
| X20 | mlrABCD | MC-LR | α-proteobacteria | [122] | |
| YF1 | mlrABCD | MC-LR | α-proteobacteria | [123] | |
| Stenotrophomonas sp. | EMS | mlrA | MC-LR, MC-RR | γ-proteobacteria | [124] |
| LEw-1278 | MC-LR | γ-proteobacteria | [74] | ||
| MC-LTH2 | MC-LR, MC-RR | γ-proteobacteria | [125] | ||
| 4B4 | mlrABCD | MC-LR, MC-RR, MC-LW, MC-LF | γ-proteobacteria | [16] | |
| Acinetobacter sp. | Bacterial | MC-LR | γ-proteobacteria | [126] | |
| Aeromonas sp. | consortia | γ-proteobacteria | |||
| Novosphingobium sp. | α-proteobacteria | ||||
| Ochrobactrum sp. | α-proteobacteria | ||||
| Pseudomonas sp. | γ-proteobacteria | ||||
| Rhodococcus sp. | Actinobacteria | ||||
| Sphingomonas sp. | α-proteobacteria | ||||
| Sphingopyxis sp. | α-proteobacteria | ||||
| Stenotrophomonas sp. | γ-proteobacteria | ||||
| Steroidobacter sp. | γ-proteobacteria | ||||
| Chryseobacterium sp. | TSFU | mlrABC | MC-LR | Flavobacteriia | [127] |
| Pseudomonas fragi | γ-proteobacteria | ||||
| Alcaligenes faecalis | YFMCD4 | MC-LR | β-proteobacteria | [128] | |
| Stenotrophomonas acidaminiohila | γ-proteobacteria | ||||
| Klebsiella sp. | YFMCD1 | MC-LR | γ-proteobacteria | [129] | |
| Stenotrophomonas sp. | γ-proteobacteria | ||||
| Acinetobacter sp. | Indigenous | mlrAD | MC-LR | γ-proteobacteria | [130] |
| Hyphomicrobium aestuarii | bacterial | α-proteobacteria | |||
| Pseudoxanthomonas sp. | mixed | γ-proteobacteria | |||
| Rhizobium sp. | culture | α-proteobacteria | |||
| Sphingobium sp. | α-proteobacteria | ||||
| Sphingomonas sp. | α-proteobacteria | ||||
| Steroidobacter sp. | γ-proteobacteria | ||||
| Agrobacterium sp. | Natural | MC-LR, Des-MCLR | α-proteobacteria | [131] | |
| Bosea sp. | bacterial | α-proteobacteria | |||
| Brevundimonas sp. | community | α-proteobacteria | |||
| Hyphomicrobium sp. | α-proteobacteria | ||||
| Rasbo sp. | α-proteobacteria | ||||
| Rhizobium sp. | α-proteobacteria | ||||
| Rhodococcus sp. | Actinobacteria | ||||
| Roseomonas sp. | α-proteobacteria | ||||
| Mesorhizobium sp. | α-proteobacteria | ||||
| Nitrosococcus sp. | γ-proteobacteria | ||||
| Sandaracinobacter sp. | α-proteobacteria | ||||
| Sphingomonas sp. | α-proteobacteria | ||||
| Bordetella sp. | Bacterial | mlrABCD | MC-LR | β-proteobacteria | [132] |
| Burkholderia sp. | community | β-proteobacteria | |||
| Cupriavidus sp. | β-proteobacteria | ||||
| Methylotenera sp. | β-proteobacteria | ||||
| Polaromonas sp. | β-proteobacteria | ||||
| Polynucleobacter sp. | β-proteobacteria | ||||
| Ralstonia sp. | γ-proteobacteria | ||||
| Variovorax sp. | β-proteobacteria | ||||
| Microbacterium sp. | Bacterial | MC-LR | Actinobacteria | [133] | |
| Rhizobium sp. | consortium | α-proteobacteria |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Massey, I.Y.; Yang, F. A Mini Review on Microcystins and Bacterial Degradation. Toxins 2020, 12, 268. https://doi.org/10.3390/toxins12040268
Massey IY, Yang F. A Mini Review on Microcystins and Bacterial Degradation. Toxins. 2020; 12(4):268. https://doi.org/10.3390/toxins12040268
Chicago/Turabian StyleMassey, Isaac Yaw, and Fei Yang. 2020. "A Mini Review on Microcystins and Bacterial Degradation" Toxins 12, no. 4: 268. https://doi.org/10.3390/toxins12040268
APA StyleMassey, I. Y., & Yang, F. (2020). A Mini Review on Microcystins and Bacterial Degradation. Toxins, 12(4), 268. https://doi.org/10.3390/toxins12040268