Use of Mass Spectrometry to Determine the Diversity of Toxins Produced by Gambierdiscus and Fukuyoa Species from Balearic Islands and Crete (Mediterranean Sea) and the Canary Islands (Northeast Atlantic)

 ,
,  ,
,  ,
,  , ,
, , 

Abstract
:1. Introduction
2. Results
2.1. LC-MS/MS Analysis
2.2. LC-HRMS/MS Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reference Toxins and Chemicals
5.2. Gambierdiscus and Fukuyoa Strains
Culturing, Harvesting, Toxin Extraction of Gambierdiscus and Fukuyoa, and N2a Assay
5.3. Sample Pretreatment
5.4. LC-MS Analysis
5.4.1. LC-MS/MS (API 4000 QTrap)
5.4.2. LC-HRMS and HRMS/MS (Q-Tof 6550 iFunnel)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yasumoto, T. The chemistry and biological function of natural marine toxins. Chem. Rec. 2001, 1, 228–242. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Murata, M. Marine toxins. Chem. Rev. 1993, 93, 1897–1909. [Google Scholar] [CrossRef]
- Oshiro, N.; Yogi, K.; Asato, S.; Sasaki, T.; Tamanaha, K.; Hirama, M.; Yasumoto, T.; Inafuku, Y. Ciguatera incidence and fish toxicity in Okinawa, Japan. Toxicon 2010, 56, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Diogène, J.; Reverté, L.; Rambla-Alegre, M.; Del Río, V.; De La Iglesia, P.; Campàs, M.; Palacios, O.; Flores, C.; Caixach, J.; Ralijaona, C.; et al. Identification of ciguatoxins in a shark involved in a fatal food poisoning in the Indian Ocean. Sci. Rep. 2017, 7, 8240. [Google Scholar] [CrossRef] [PubMed]
- Estevez, P.; Castro, D.; Valtierra, A.P.; Leão-Martins, J.M.; Vilariño, O.; Diogène, J.; Gago-Martínez, A. An Attempt to Characterize the Ciguatoxin Profile in Seriola fasciata Causing Ciguatera Fish Poisoning in Macaronesia. Toxins 2019, 11, 221. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.S.; Boundy, M.J.; Selwood, A.I.; Harwood, T. Development of an LC–MS/MS method to simultaneously monitor maitotoxins and selected ciguatoxins in algal cultures and P-CTX-1B in fish. Harmful Algae 2018, 80, 80–87. [Google Scholar] [CrossRef]
- Abraham, A.; Jester, E.L.; Granade, H.R.; Plakas, S.M.; Dickey, R.W. Caribbean ciguatoxin profile in raw and cooked fish implicated in ciguatera. Food Chem. 2012, 131, 192–198. [Google Scholar] [CrossRef]
- Murata, M.; Naoki, H.; Matsunaga, S.; Satake, M.; Yasumoto, T. Structure and Partial Stereochemical Assignments for Maitotoxin, the Most Toxic and Largest Natural Non-Biopolymer. J. Am. Chem. Soc. 1994, 116, 7098–7107. [Google Scholar] [CrossRef]
- Murata, M.; Yasumoto, T. The structure elucidation and biological activities of high molecular weight algal toxins: Maitotoxin, prymnesins and zooxanthellatoxins. Nat. Prod. Rep. 2000, 17, 293–314. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J. Purification and characterisation of large and small maitotoxins from cultured Gambierdiscus toxicus. Nat. Toxins 1994, 2, 64–72. [Google Scholar] [CrossRef]
- Pisapia, F.; Sibat, M.; Herrenknecht, C.; Lhaute, K.; Gaiani, G.; Ferron, P.-J.; Fessard, V.; Fraga, S.; Nascimento, S.M.; Litaker, R.W.; et al. Maitotoxin-4, a Novel MTX Analog Produced by Gambierdiscus excentricus. Mar. Drugs 2017, 15, 220. [Google Scholar] [CrossRef] [PubMed]
- Mazzola, E.P.; Deeds, J.R.; Stutts, W.L.; Ridge, C.D.; Dickey, R.W.; White, K.D.; Williamson, R.T.; Martin, G.E. Elucidation and partial NMR assignment of monosulfated maitotoxins from the Caribbean. Toxicon 2019, 164, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Boente-Juncal, A.; Álvarez, M.; Antelo, Á.; Rodríguez, I.; Calabro, K.; Vale, C.; Thomas, O.P.; Botana, L.M. Structure Elucidation and Biological Evaluation of Maitotoxin-3, a Homologue of Gambierone, from Gambierdiscus belizeanus. Toxins 2019, 11, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, J.S.; Selwood, A.; Harwood, D.T.; Van Ginkel, R.; Puddick, J.; Rhodes, L.L.; Rise, F.; Wilkins, A.L. 44-Methylgambierone, a new gambierone analogue isolated from Gambierdiscus australes. Tetrahedron Lett. 2019, 60, 621–625. [Google Scholar] [CrossRef]
- Rodríguez, I.; Genta-Jouve, G.; Alfonso, C.; Calabro, K.; Alonso, E.; Sánchez, J.A.; Alfonso, A.; Thomas, O.P.; Botana, L.M. Gambierone, a Ladder-Shaped Polyether from the Dinoflagellate Gambierdiscus belizeanus. Org. Lett. 2015, 17, 2392–2395. [Google Scholar] [CrossRef]
- Satake, M.; Murata, M.; Yasumoto, T. Gambierol: A new toxic polyether compound isolated from the marine dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1993, 115, 361–362. [Google Scholar] [CrossRef]
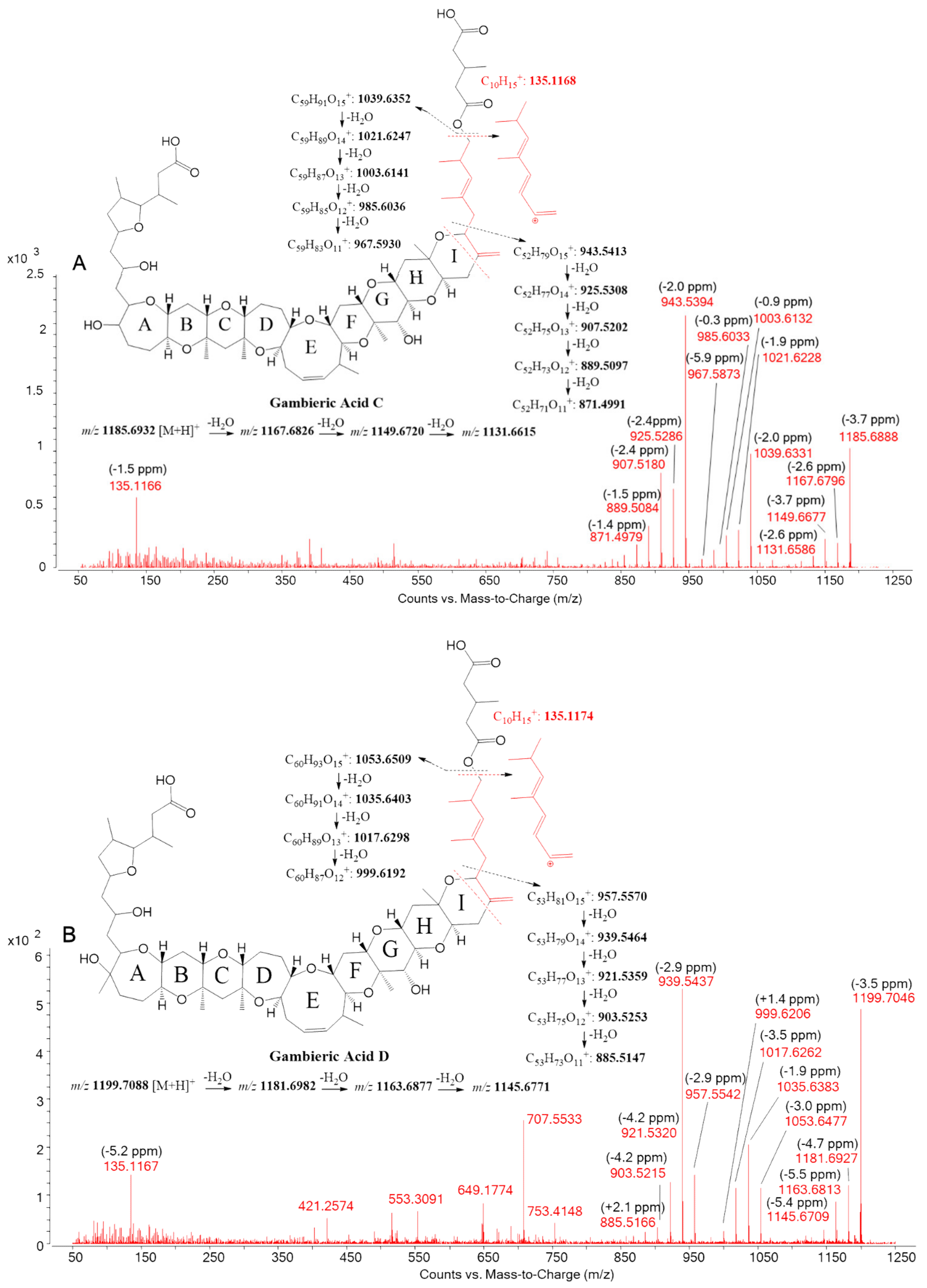
- Nagai, H.; Torigoe, K.; Satake, M.; Murata, M.; Yasumoto, T.; Hirota, H. Gambieric acids: Unprecedented potent antifungal substances isolated from cultures of a marine dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1992, 114, 1102–1103. [Google Scholar] [CrossRef]
- Watanabe, R.; Uchida, H.; Suzuki, T.; Matsushima, R.; Nagae, M.; Toyohara, Y.; Satake, M.; Oshima, Y.; Inoue, A.; Yasumoto, T. Gambieroxide, a novel epoxy polyether compound from the dinoflagellate Gambierdiscus toxicus GTP2 strain. Tetrahedron 2013, 69, 10299–10303. [Google Scholar] [CrossRef]
- Litaker, R.W.; Vandersea, M.; Faust, M.A.; Kibler, S.; Nau, A.W.; Holland, W.C.; Chinain, M.; Holmes, M.J.; Tester, P.A. Global distribution of ciguatera causing dinoflagellates in the genus Gambierdiscus. Toxicon 2010, 56, 711–730. [Google Scholar] [CrossRef]
- Pisapia, F.; Holland, W.C.; Hardison, D.R.; Litaker, R.W.; Fraga, S.; Nishimura, T.; Adachi, M.; Nguyen-Ngoc, L.; Séchet, V.; Amzil, Z.; et al. Toxicity screening of 13 Gambierdiscus strains using neuro-2a and erythrocyte lysis bioassays. Harmful Algae 2017, 63, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Fraga, S.; Rodríguez, F. Genus Gambierdiscus in the Canary Islands (NE Atlantic Ocean) with Description of Gambierdiscus silvae sp. nov., a New Potentially Toxic Epiphytic Benthic Dinoflagellate. Protist 2014, 165, 839–853. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, F.; Fraga, S.; Ramilo, I.; Rial, P.; Figueroa, R.I.; Riobó, P.; Bravo, I. Canary Islands (NE Atlantic) as a biodiversity ‘hotspot’ of Gambierdiscus: Implications for future trends of ciguatera in the area. Harmful Algae 2017, 67, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Reverté, L.; Toldrà, A.; Andree, K.B.; Fraga, S.; De Falco, G.; Campàs, M.; Diogène, J. Assessment of cytotoxicity in ten strains of Gambierdiscus australes from Macaronesian Islands by neuro-2a cell-based assays. Environ. Boil. Fishes 2018, 30, 2447–2461. [Google Scholar] [CrossRef]
- Aligizaki, K.; Nikolaidis, G. Morphological identification of two tropical dinoflagellates of the genera Gambierdiscus and Sinophysis in the Mediterranean Sea. J. Biol. Res. Thessaloniki 2008, 9, 75–82. [Google Scholar]
- Laza-Martínez, A.; David, H.; Riobó, P.; Miguel, I.; Orive, E. Characterization of a Strain of Fukuyoa paulensis (Dinophyceae) from the Western Mediterranean Sea. J. Eukaryot. Microbiol. 2016, 63, 481–497. [Google Scholar] [CrossRef] [Green Version]
- AECOSAN Website. Available online: http://www.aecosan.msssi.gob.es/AECOSAN/web/ciguatera/home/aecosan_home_ciguatera.htm (accessed on 25 March 2020).
- Lewis, R.J.; Holmes, M.J.; Alewood, P.F.; Jones, A. Lonspray mass spectrometry of ciguatoxin-1, maitotoxin-2 and -3, and related marine polyether toxins. Nat. Toxins 1994, 2, 56–63. [Google Scholar] [CrossRef]
- Rhodes, L.L.; Smith, K.F.; Murray, J.S.; Harwood, T.; Trnski, T.; Munday, R. The Epiphytic Genus Gambierdiscus (Dinophyceae) in the Kermadec Islands and Zealandia Regions of the Southwestern Pacific and the Associated Risk of Ciguatera Fish Poisoning. Mar. Drugs 2017, 15, 219. [Google Scholar] [CrossRef] [Green Version]
- Kohli, G.S.; Papiol, G.G.; Rhodes, L.L.; Harwood, T.; Selwood, A.; Jerrett, A.R.; Murray, S.A.; Neilan, B.A. A feeding study to probe the uptake of Maitotoxin by snapper (Pagrus auratus). Harmful Algae 2014, 37, 125–132. [Google Scholar] [CrossRef]
- Nagai, H.; Murata, M.; Torigoe, K.; Satake, M.; Yasumoto, T. Gambieric acids, new potent antifungal substances with unprecedented polyether structures from a marine dinoflagellate Gambierdiscus toxicus. J. Org. Chem. 1992, 57, 5448–5453. [Google Scholar] [CrossRef]
- Munday, R.; Murray, J.S.; Rhodes, L.L.; Larsson, M.E.; Harwood, T. Ciguatoxins and Maitotoxins in Extracts of Sixteen Gambierdiscus Isolates and One Fukuyoa Isolate from the South Pacific and Their Toxicity to Mice by Intraperitoneal and Oral Administration. Mar. Drugs 2017, 15, 208. [Google Scholar] [CrossRef] [Green Version]
- Larsson, M.E.; Harwood, T.D.; Lewis, R.J.; Himaya, S.W.A.; Doblin, M.A. Toxicological characterization of Fukuyoa paulensis (Dinophyceae) from temperate Australia. Phycol. Res. 2018, 67, 65–71. [Google Scholar] [CrossRef]
- Tudó, À.; Toldrà, A.; Rey, M.; Todolí, I.; Andree, K.B.; Fernández-Tejedor, M.; Campàs, M.; Sureda, F.X.; Diogène, J. Gambierdiscus and Fukuyoa as potential indicators of ciguatera risk in the Balearic Islands. Harmful Algae 2020. submitted. [Google Scholar]
- Vial, J.; Jardy, A. Experimental Comparison of the Different Approaches To Estimate LOD and LOQ of an HPLC Method. Anal. Chem. 1999, 71, 2672–2677. [Google Scholar] [CrossRef]
- Sanagi, M.M.; Ling, S.L.; Nasir, Z.; Hermawan, D.; Ibrahim, W.A.W.; Abu Naim, A. Comparison of Signal-to-Noise, Blank Determination, and Linear Regression Methods for the Estimation of Detection and Quantification Limits for Volatile Organic Compounds by Gas Chromatography. J. AOAC Int. 2009, 92, 1833–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
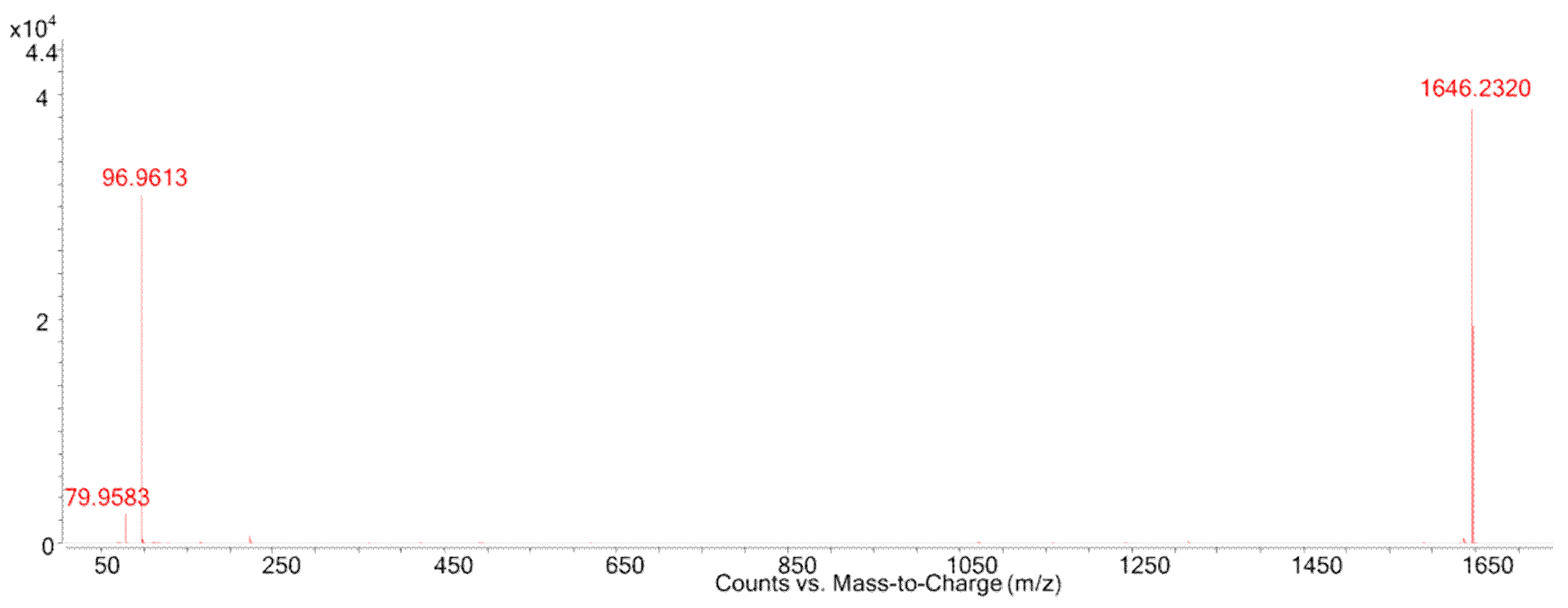
| MTX Congener | Chemical Formula | Monoisotopic Mass (Da) | Reference |
|---|---|---|---|
| MTX1 | C164H258O68S2 | 3379.6171 for the free acid form | [9] |
| MTX2 | Unknown | 3298 for the mono-sodium salt | [10] |
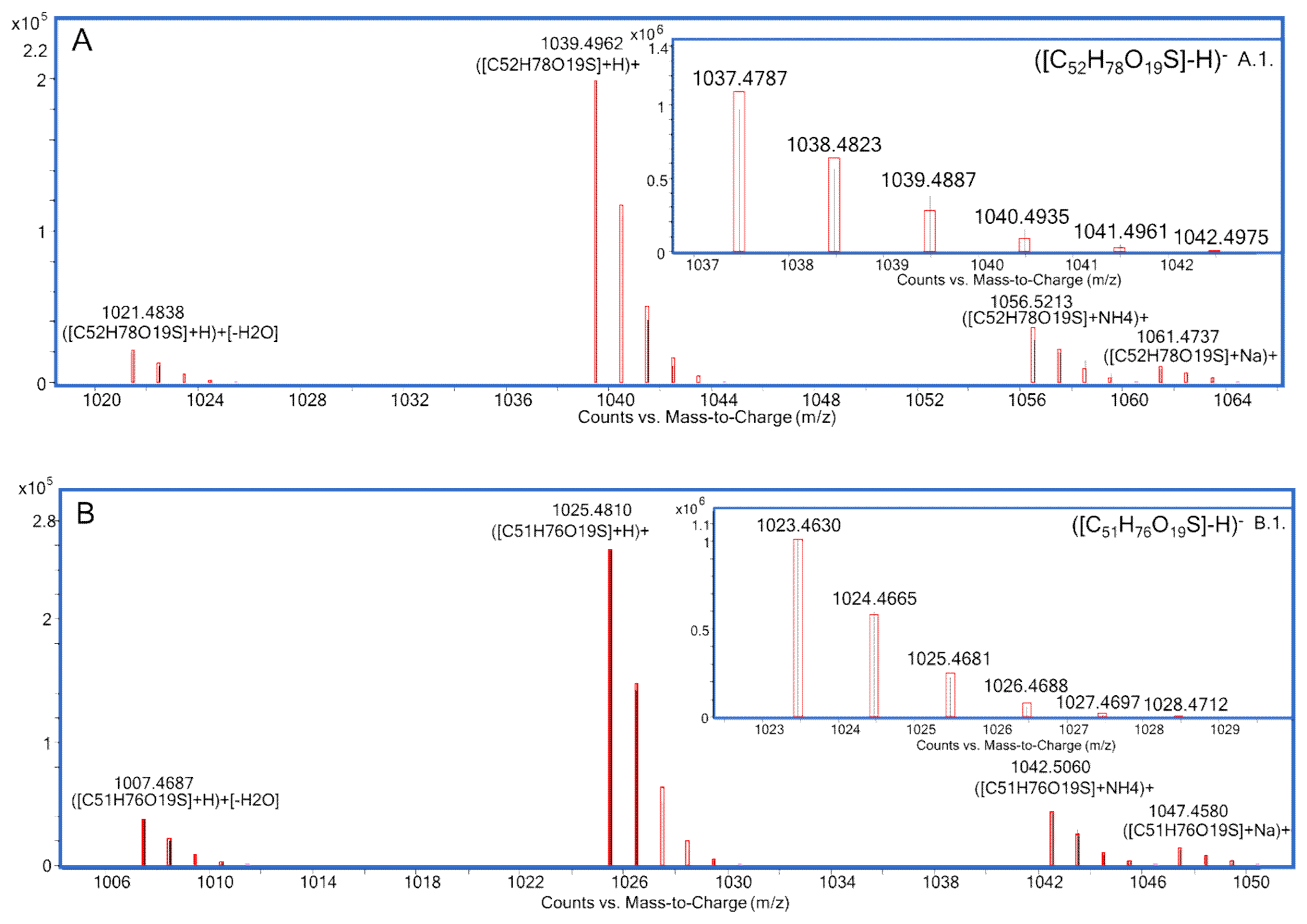
| MTX3 | C52H78O19S | 1038.4858 for the free acid form | [13,14] |
| MTX4 | Unknown | 3292.486 for the free acid form | [11] |
| desulfo-MTX1 | C164H258O65S | 3299.6603 for the free acid form | [12] |
| didehydro-demethyl-desulfo-MTX1 | C163H254O65S | 3283.6290 for the free acid form | [12] |
| Species | Strain Code | Location | pg MTX1 eq./Cell (% of TOTAL) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MTX1 | MTX2 | MTX3 | MTX4 | desulfo-MTX1 | didehydro-demethyl-desulfo-MTX1 | Gambierone | P-Gambierone Analogue | |||||||||||
| MSF (%) | DSF (%) | Σ | MSF (%) | DSF (%) | Σ | MSF (%) | DSF (%) | Σ | MSF (%) | DSF (%) | Σ | |||||||
| G. australes | IRTA-SMM-17-189 | Torret, Menorca, Balearic Islands, Spain | n.d. | n.d. | 241 (70.2) | 102 (29.8) | 344.0 | n.d. | n.d. | n.d. | n.d. | 148 (99.7) | 0.4 (0.3) | 149 | ||||
| G. australes | IRTA-SMM-17-162 | St. Adeodat, Menorca, Balearic Islands, Spain | n.d. | n.d. | 575 (79.9) | 150 (20.1) | 720 | n.d. | n.d. | n.d. | n.d. | 275 (99.6) | 1.0 (0.4) | 276 | ||||
| G. australes | IRTA-SMM-17-164 | St. Adeodat, Menorca, Balearic Islands, Spain | n.d. | n.d. | 1108 (66.7) | 553 (33.3) | 1661 | n.d. | n.d. | n.d. | n.d. | 522 (99.8) | 1.2 (0.2) | 523 | ||||
| G. australes | IRTA-SMM-17-271 | Macarella, Menorca, Balearic Islands, Spain | n.d. | n.d. | 1107 (83.7) | 215 (16.3) | 1322 | n.d. | n.d. | n.d. | n.d. | 408 (99.9) | 0.4 (0.1) | 409 | ||||
| F. paulensis | IRTA-SMM-17-209 | Sacaleta, Menorca, Balearic Islands, Spain | n.d. | n.d. | 1.8 (17.7) | 8.6 (82.3) | 10.5 | n.d. | n.d. | n.d. | n.d. | n.d. | ||||||
| G. australes | IRTA-SMM-17-253 | Anguila, Mallorca, Balearic Islands, Spain | n.d. | n.d. | 781 (72.3) | 300 (27.7) | 1081 | n.d. | n.d. | n.d. | n.d. | 229 (99.8) | 0.5 (0.2) | 229 | ||||
| G. australes | IRTA-SMM-17-244 | Camp de Mar, Mallorca, Balearic Islands, Spain | n.d. | n.d. | 403 (84.1) | 75.9 (15.9) | 479 | n.d. | n.d. | n.d. | n.d. | 173 (99.6) | 0.7 (0.4) | 174 | ||||
| Gambierdiscus sp.2 | 0010G-CR-CCAUTH | Kolimpari, Crete, Greece | n.d. | n.d. | 3.0 (70.1) | 1.3 (29.9) | 4.3 | n.d. | n.d. | n.d. | 716 (92.2) | 60.3 (7.8) | 775.9 | n.d. | ||||
| G. excentricus | IRTA-SMM-17-407 | Playa de vueltas, La Gomera, Canary Islands, Spain | n.d. | n.d. | n.d. | 36.8 (99.3) | 0.3 (0.7) | 37.1 | n.d. | n.d. | n.d. | n.d. | ||||||
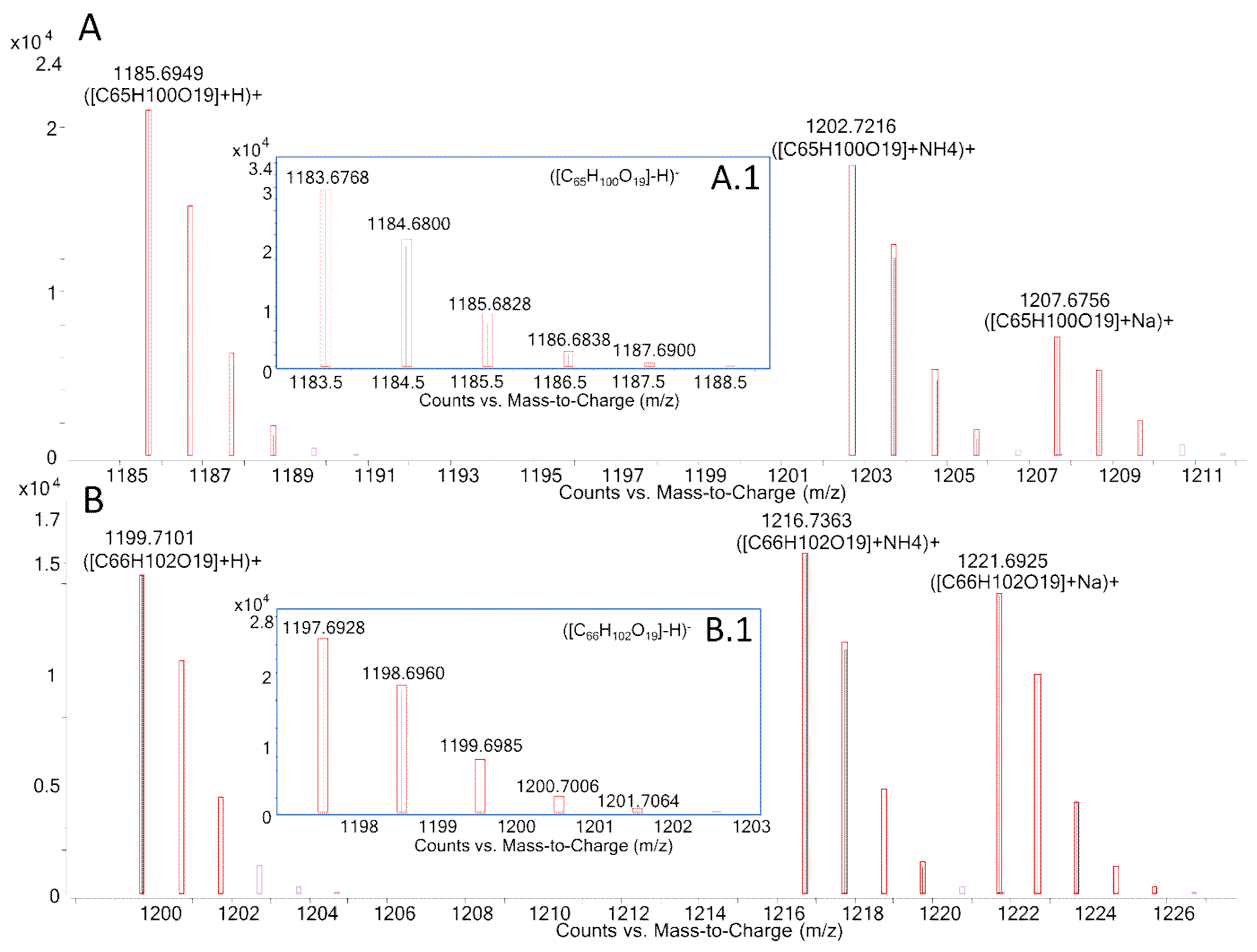
| Ion | MTX3 | Gambierone | ||||||
|---|---|---|---|---|---|---|---|---|
| Molecular Formula | m/z | Δ ppm | Molecular Formula | m/z | Δ ppm | |||
| Measured | Theoretical | Measured | Theoretical | |||||
| ESI− | ||||||||
| [M-H]− | C52H77O19S− | 1037.4774 | 1037.4785 | −1.1 | C51H75O19S− | 1023.4612 | 1023.4629 | −1.6 |
| [M-H-H2O]− | C52H75O18S− | 1019.4625 | 1019.4680 | −5.3 | C51H73O18S− | 1005.4463 | 1005.4523 | −6.0 |
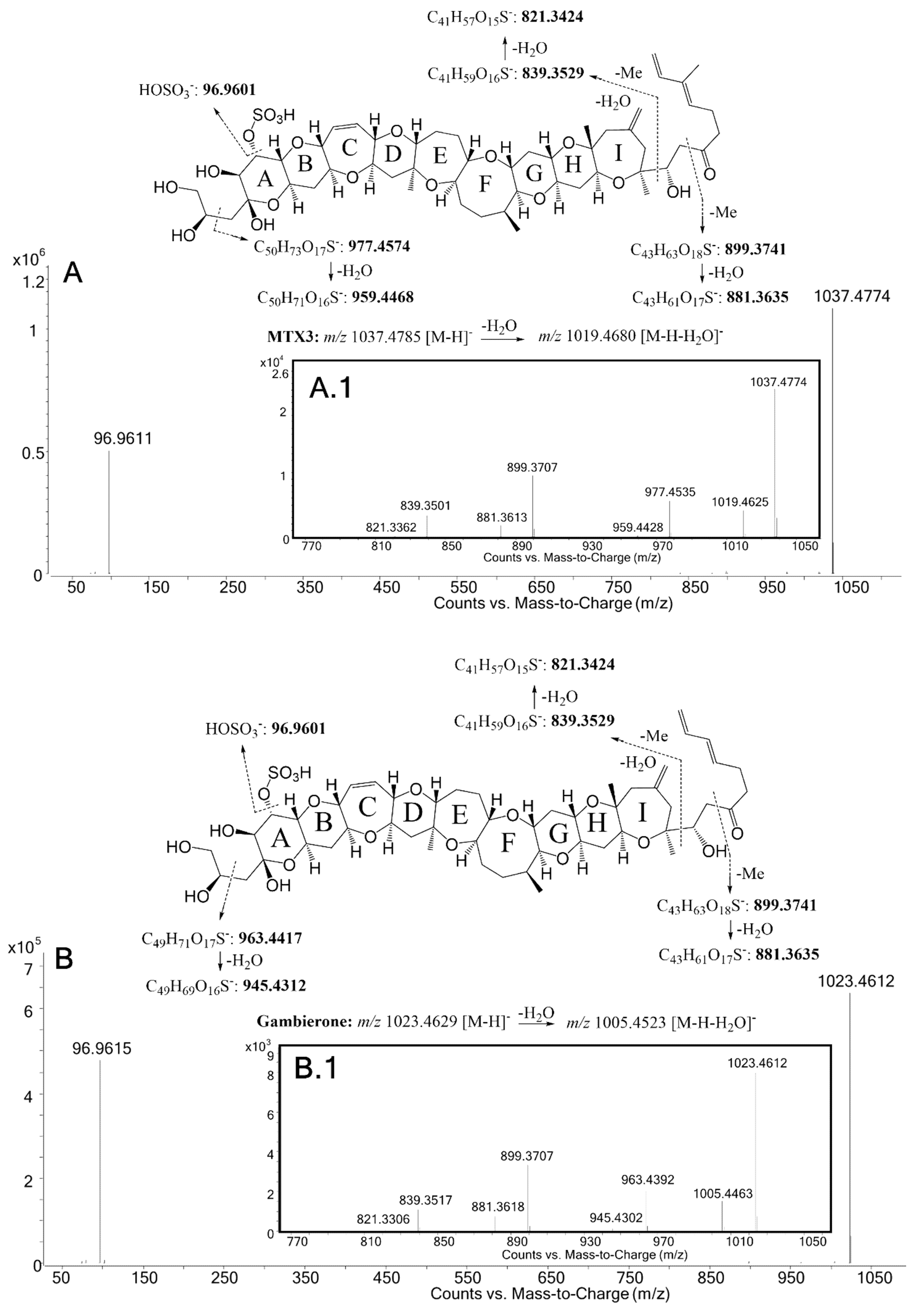
| C50H73O17S− | 977.4535 | 977.4574 | −4.0 | C49H71O17S− | 963.4392 | 963.4417 | −2.6 | |
| H2O | C50H71O16S− | 959.4428 | 959.4468 | −4.2 | C49H69O16S− | 945.4302 | 945.4312 | −1.0 |
| C43H63O18S− | 899.3707 | 899.3741 | −3.7 | C43H63O18S− | 899.3707 | 899.3741 | −3.7 | |
| -H2O | C43H61O17S− | 881.3613 | 881.3635 | −2.5 | C43H61O17S− | 881.3618 | 881.3635 | −1.9 |
| C41H59O16S− | 839.3501 | 839.3529 | −3.4 | C41H59O16S− | 839.3517 | 839.3529 | −1.5 | |
| -H2O | C41H57O15S− | 821.3362 | 821.3424 | −7.5 | C41H57O15S− | 821.3306 | 821.3424 | −14.3 |
| [HOSO3]− | HOSO3− | 96.9611 | 96.9601 | 10.3 | HOSO3− | 96.9615 | 96.9601 | 14.4 |
| ESI+ | ||||||||
| [M+H]+ | C52H79O19S+ | 1039.4911 | 1039.4931 | −1.9 | C51H77O19S+ | 1025.4749 | 1025.4774 | −2.4 |
| [M+H-H2O]+ | C52H77O18S+ | 1021.4795 | 1021.4825 | −2.9 | C51H75O18S+ | 1007.4636 | 1007.4668 | −3.2 |
| [M+H-2H2O]+ | C52H75O17S+ | 1003.4692 | 1003.4720 | −2.8 | C51H73O17S+ | 989.4528 | 989.4563 | −3.5 |
| [M-SO3+H]+ | C52H79O16+ | 959.5333 | 959.5363 | −3.1 | C51H77O16+ | 945.5171 | 945.5206 | −3.7 |
| [M-SO3-H2O+H]+ | C52H77O15+ | 941.5232 | 941.5257 | −2.7 | C51H75O15+ | 927.5070 | 927.5100 | −3.2 |
| [M-SO3-2H2O+H]+ | C52H75O14+ | 923.5125 | 923.5152 | −2.9 | C51H73O14+ | 909.4968 | 909.4995 | −3.0 |
| [M-SO3-3H2O+H]+ | C52H73O13+ | 905.5022 | 905.5046 | −2.7 | C51H71O13+ | 891.4849 | 891.4889 | −4.5 |
| [M-SO3-4H2O+H]+ | C52H71O12+ | 887.4911 | 887.4940 | −3.3 | C51H69O12+ | 873.4743 | 873.4783 | −4.6 |
| [M-SO3-5H2O+H]+ | C52H69O11+ | 869.4812 | 869.4835 | −2.6 | C51H67O11+ | 855.4656 | 855.4678 | −2.6 |
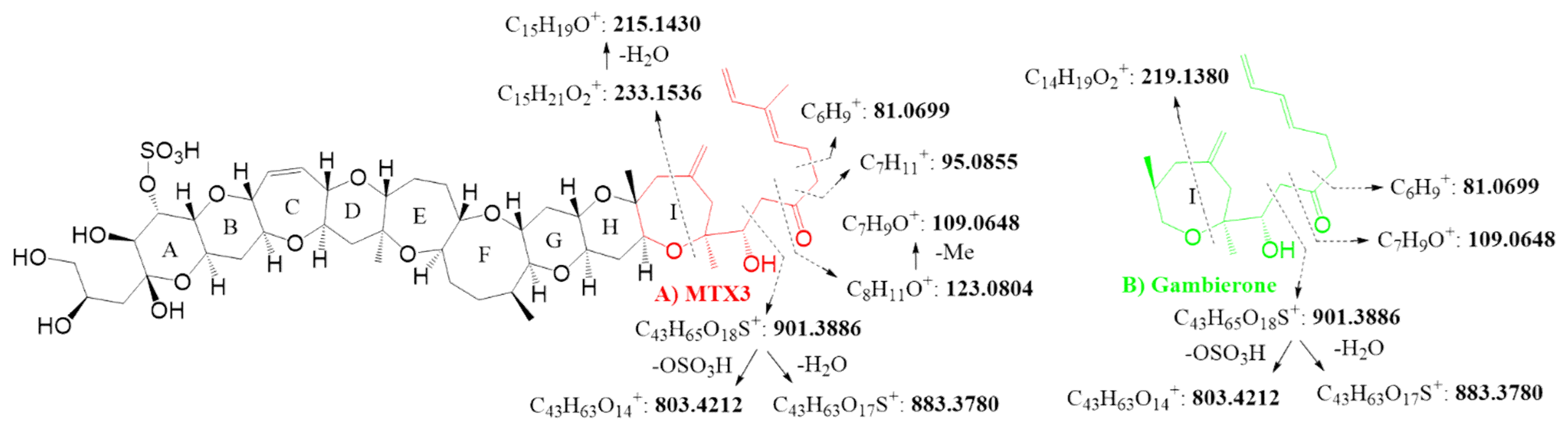
| C43H65O18S+ | 901.3854 | 901.3886 | −3.6 | C43H65O18S+ | 901.3884 | 901.3886 | −0.2 | |
| -H2O | C43H63O17S+ | 883.3756 | 883.3780 | −2.7 | C43H63O17S+ | 883.3744 | 883.3780 | −4.1 |
| -OSO3H | C43H63O14+ | 803.4182 | 803.4212 | −3.7 | C43H63O14+ | 803.4173 | 803.4212 | −4.9 |
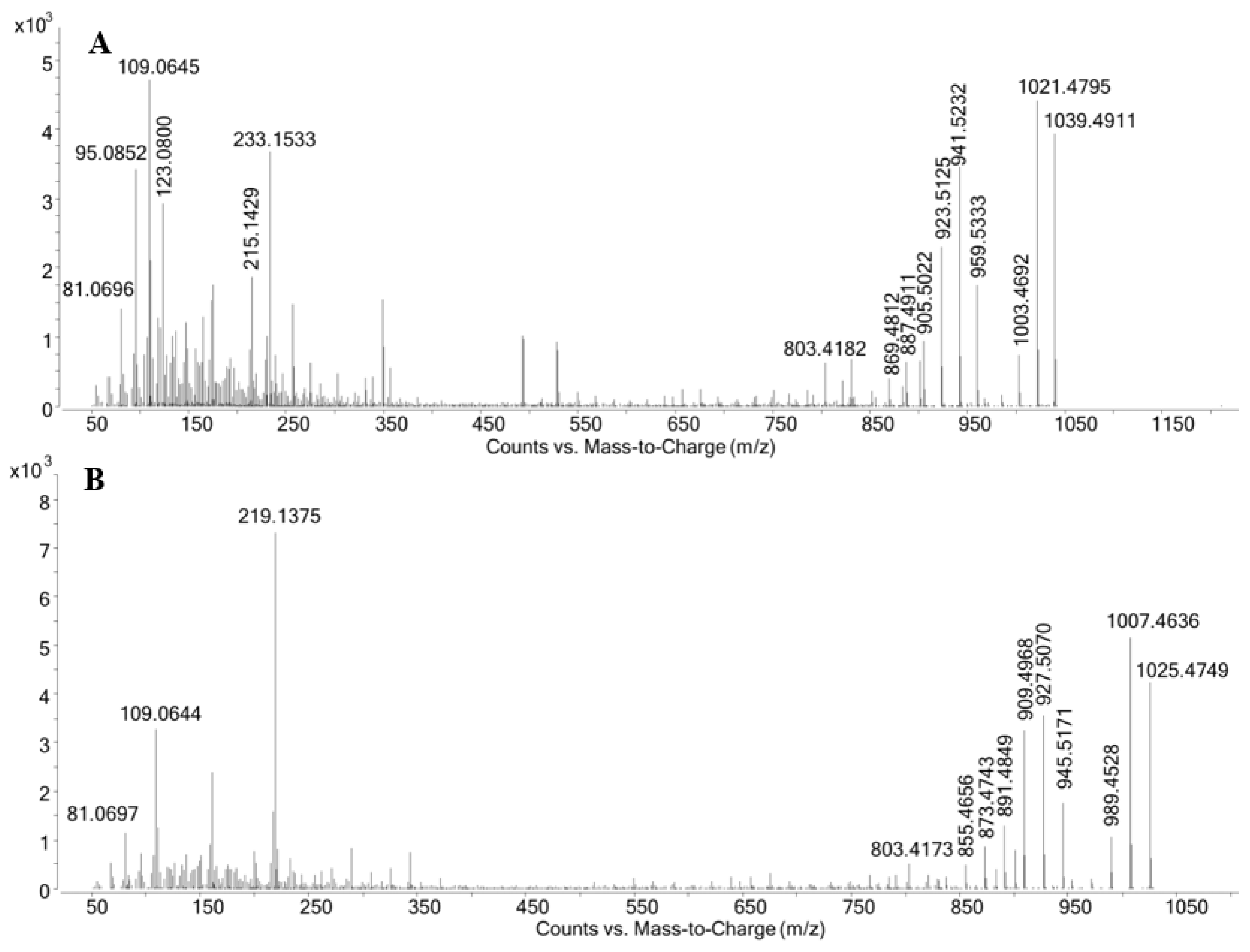
| C15H21O2+ | 233.1533 | 233.1536 | −1.3 | C14H19O2+ | 219.1375 | 219.1380 | −2.1 | |
| C15H19O+ | 215.1429 | 215.1430 | −0.5 | C7H9O+ | 109.0644 | 109.0648 | −3.6 | |
| C8H11O+ | 123.0800 | 123.0804 | −3.2 | C6H9+ | 81.0697 | 81.0699 | −2.2 | |
| C7H9O+ | 109.0645 | 109.0648 | −2.8 | |||||
| C7H11+ | 95.0852 | 95.0855 | −3.2 | |||||
| C6H9+ | 81.0696 | 81.0699 | −3.7 | |||||
| Species | Strain Code | Location | Number of Cells Extracted | Volume of Culture (L) | CTX-like (fg CTX1B Equiv./Cell) |
|---|---|---|---|---|---|
| G. australes | IRTA-SMM-17-189 | Torret, Menorca, Balearic Islands, Spain | 17 134 000 | 20 | 83 ± 12 a |
| G. australes | IRTA-SMM-17-162 | St. Adeodat, Menorca, Balearic Islands, Spain | 27 811 000 | 20 | 101 ± 7.5 |
| G. australes | IRTA-SMM-17-164 | St. Adeodat, Menorca, Balearic Islands, Spain | 4 257 000 | 20 | >62.5 (NQ) |
| G. australes | IRTA-SMM-17-271 | Macarella, Menorca, Balearic Islands, Spain | 14 007 000 | 20 | 271 ± 29 |
| F. paulensis | IRTA-SMM-17-209 | Sacaleta, Menorca, Balearic Islands, Spain | 6 964 000 | 20 | 16 ± 1.7 a |
| G. australes | IRTA-SMM-17-253 | Anguila, Menorca, Balearic Islands, Spain | 13 735 000 | 20 | 164 ± 16 |
| G. australes | IRTA-SMM-17-244 | Camp de Mar, Mallorca, Balearic Islands, Spain | 4 121 000 | 5 | 155 ± 25 |
| Gambierdiscus sp.2 | 0010G-CR-CCAUTH | Kolimpari, Crete, Greece | 2 300 000 | 5 | NQ |
| G. excentricus | IRTA-SMM-17-407 | Playa de vueltas, La Gomera, Canary Islands, Spain | 6 084 000 | 5 | >794 (NQ) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estevez, P.; Sibat, M.; Leão-Martins, J.M.; Tudó, A.; Rambla-Alegre, M.; Aligizaki, K.; Diogène, J.; Gago-Martinez, A.; Hess, P. Use of Mass Spectrometry to Determine the Diversity of Toxins Produced by Gambierdiscus and Fukuyoa Species from Balearic Islands and Crete (Mediterranean Sea) and the Canary Islands (Northeast Atlantic). Toxins 2020, 12, 305. https://doi.org/10.3390/toxins12050305
Estevez P, Sibat M, Leão-Martins JM, Tudó A, Rambla-Alegre M, Aligizaki K, Diogène J, Gago-Martinez A, Hess P. Use of Mass Spectrometry to Determine the Diversity of Toxins Produced by Gambierdiscus and Fukuyoa Species from Balearic Islands and Crete (Mediterranean Sea) and the Canary Islands (Northeast Atlantic). Toxins. 2020; 12(5):305. https://doi.org/10.3390/toxins12050305
Chicago/Turabian StyleEstevez, Pablo, Manoëlla Sibat, José Manuel Leão-Martins, Angels Tudó, Maria Rambla-Alegre, Katerina Aligizaki, Jorge Diogène, Ana Gago-Martinez, and Philipp Hess. 2020. "Use of Mass Spectrometry to Determine the Diversity of Toxins Produced by Gambierdiscus and Fukuyoa Species from Balearic Islands and Crete (Mediterranean Sea) and the Canary Islands (Northeast Atlantic)" Toxins 12, no. 5: 305. https://doi.org/10.3390/toxins12050305
APA StyleEstevez, P., Sibat, M., Leão-Martins, J. M., Tudó, A., Rambla-Alegre, M., Aligizaki, K., Diogène, J., Gago-Martinez, A., & Hess, P. (2020). Use of Mass Spectrometry to Determine the Diversity of Toxins Produced by Gambierdiscus and Fukuyoa Species from Balearic Islands and Crete (Mediterranean Sea) and the Canary Islands (Northeast Atlantic). Toxins, 12(5), 305. https://doi.org/10.3390/toxins12050305