Full Neutralization of Centruroides sculpturatus Scorpion Venom by Combining Two Human Antibody Fragments

 ,
, 
Abstract
:1. Introduction
2. Results
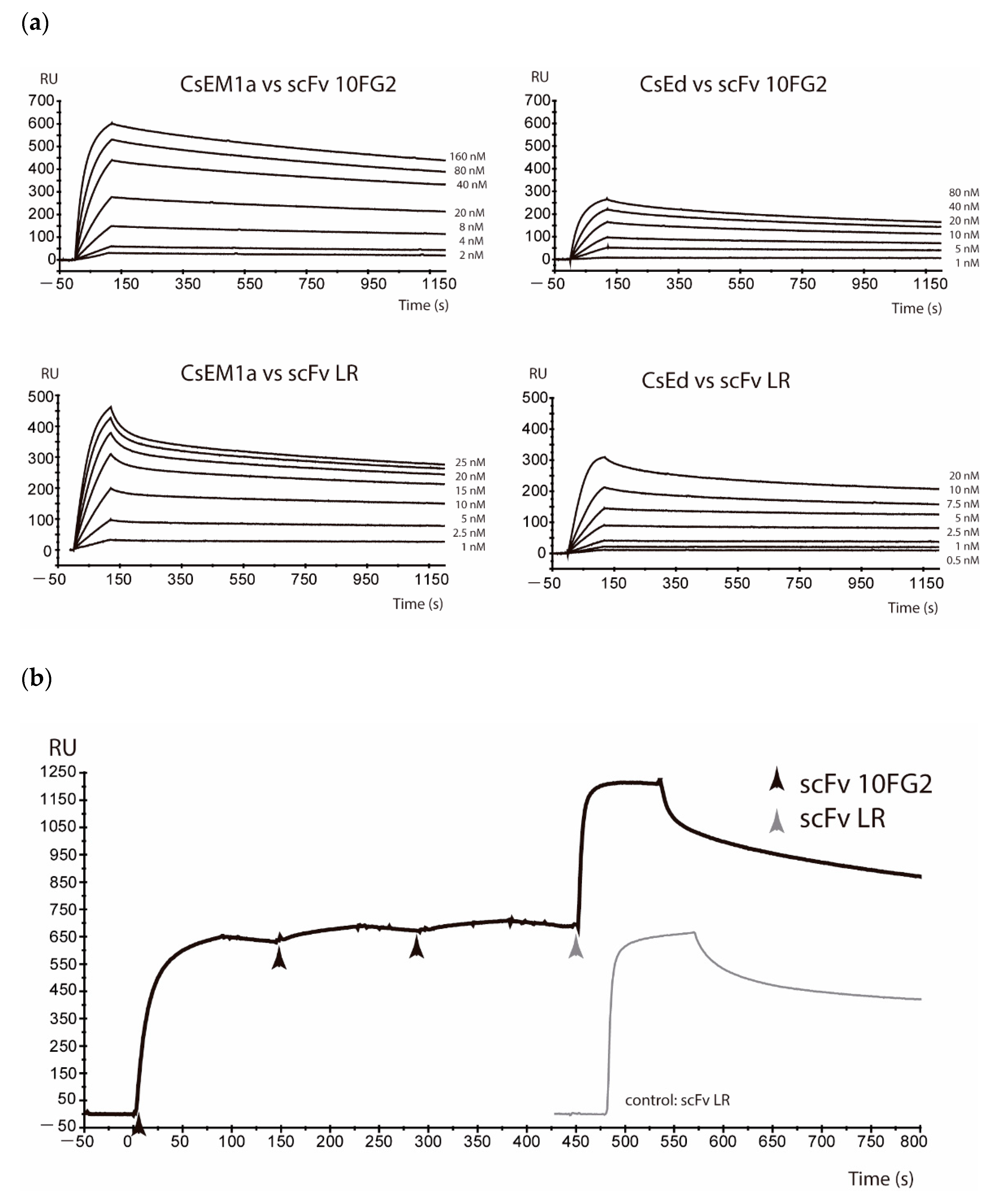
2.1. Neutralization Assays of C. sculpturatus Venom
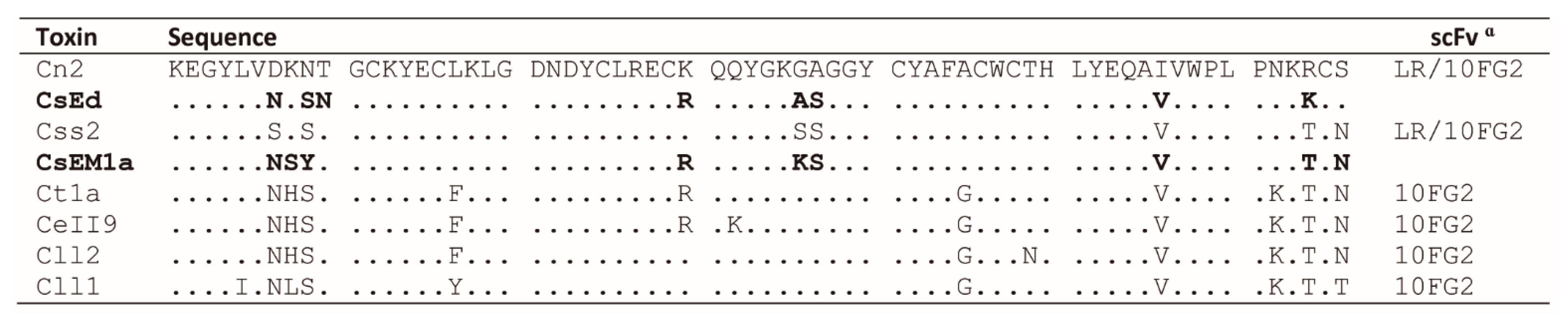
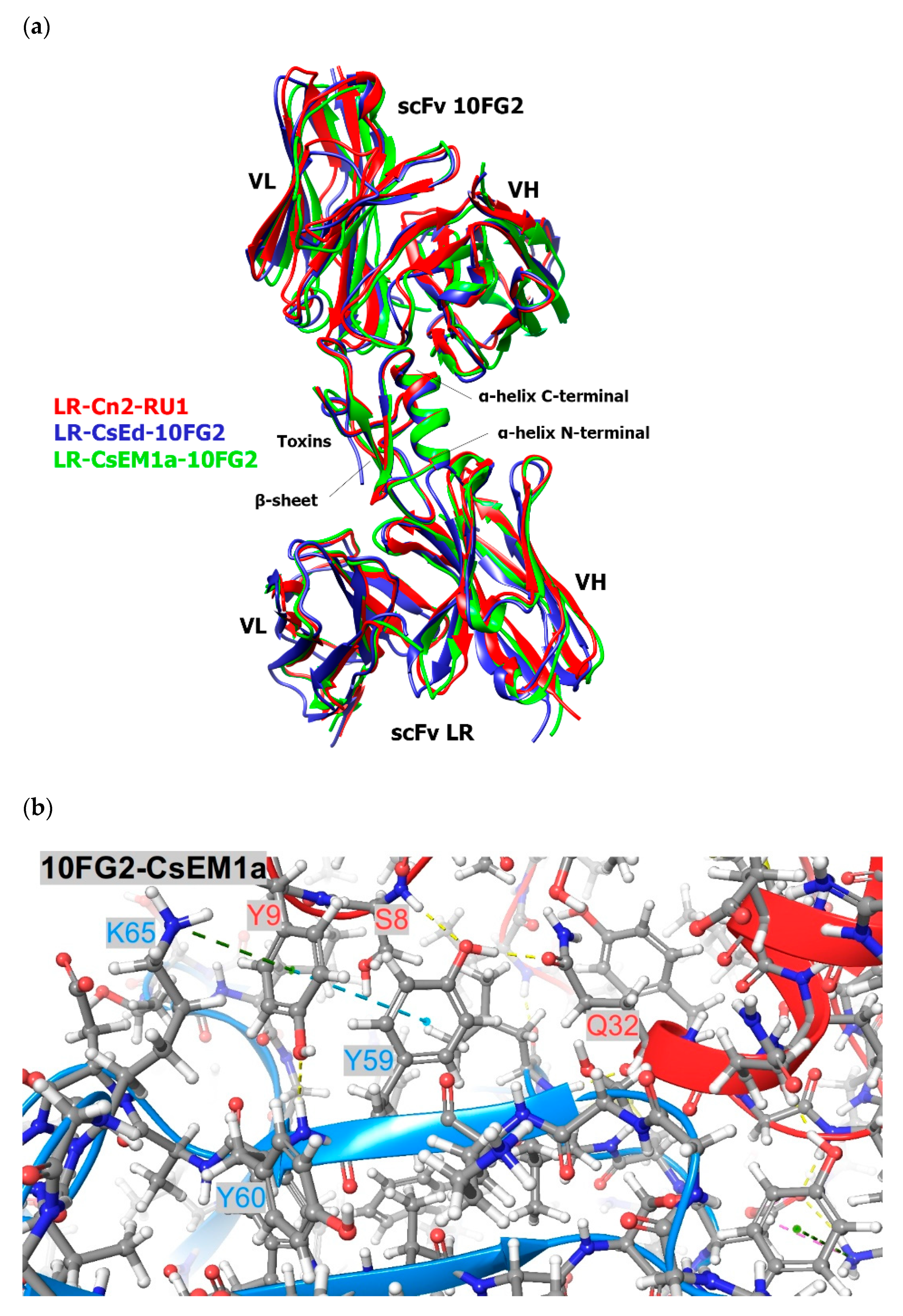
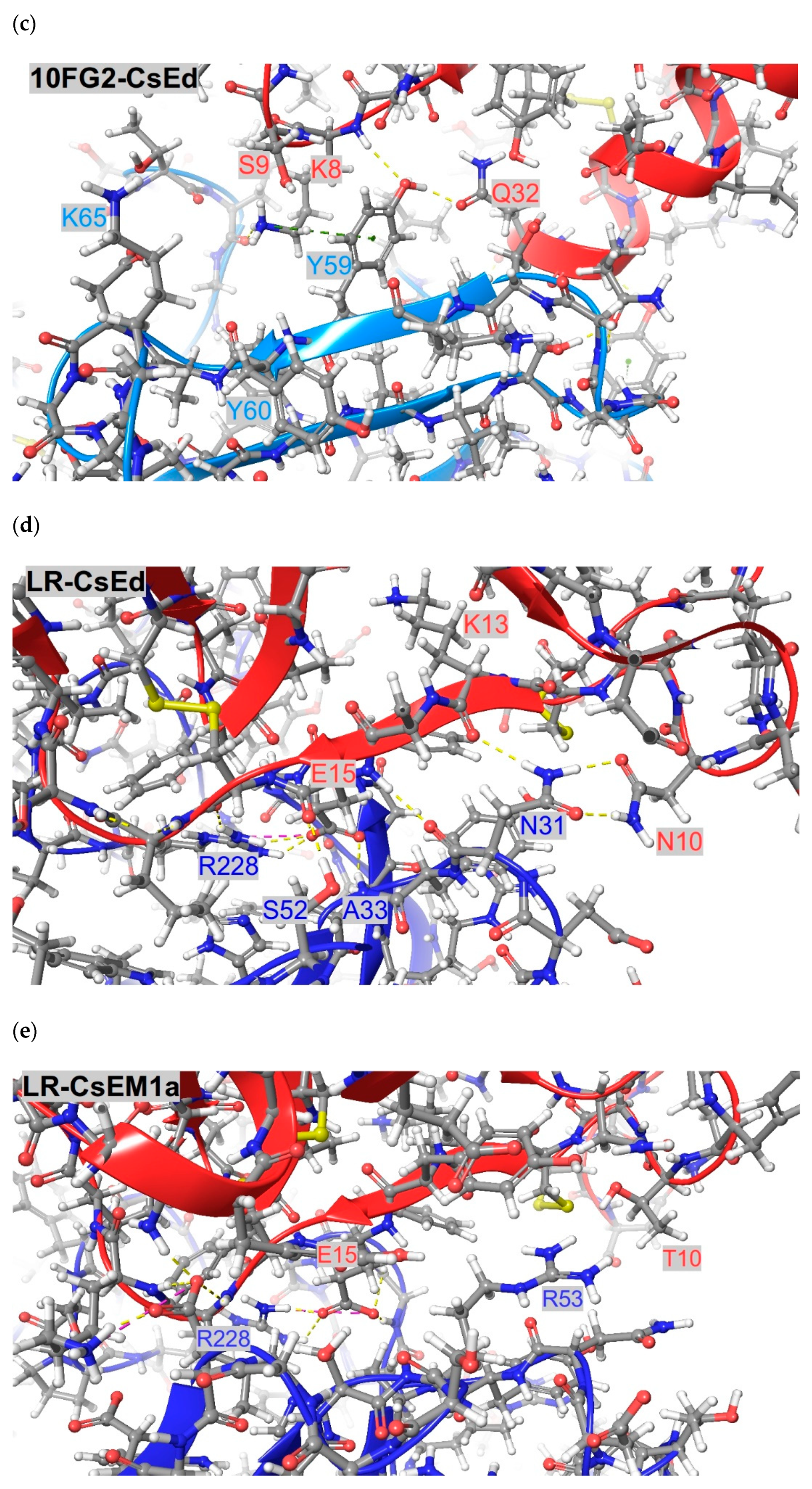
2.2. Structural Analyses of scFvs 10FG2 and LR in Complex with CsEM1a and CsEd Toxins
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Venom and Toxins
5.2. Expression of scFvs 10FG2 and LR
5.3. Surface Plasmon Resonance Recognition and Affinity Determinations
5.4. Competition Assays by Means of Surface Plasmon Resonance
5.5. Venom Neutralization Tests
5.5.1. Mixed Test
5.5.2. Rescue Test
5.6. Modelling and Structural Analyses of scFv 10FG2-CsEM1a, scFv 10FG2-CsEd, scFv LR-CsEM1a, and scFv LR-CsEd Complexes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gonzalez-Santillan, E.; Possani, L.D. North American scorpion species of public health importance with a reappraisal of historical epidemiology. Acta Trop. 2018, 187, 264–274. [Google Scholar] [CrossRef]
- Kang, A.M.; Brooks, D.E. Geographic Distribution of Scorpion Exposures in the United States, 2010–2015. Am. J. Public Health 2017, 107, 1958–1963. [Google Scholar] [CrossRef]
- Gummin, D.D.; Mowry, J.B.; Beuhler, M.C.; Spyker, D.A.; Brooks, D.E.; Dibert, K.W.; Rivers, L.J.; Pham, N.P.T.; Ryan, M.L. 2019 Annual Report of the American Association of Poison Control Centers’ National Poison Data System (NPDS): 37th Annual Report. Clin. Toxicol. 2020, 58, 1360–1541. [Google Scholar] [CrossRef] [PubMed]
- Riano-Umbarila, L.; Rodriguez-Rodriguez, E.R.; Santibanez-Lopez, C.E.; Guereca, L.; Uribe-Romero, S.J.; Gomez-Ramirez, I.V.; Carcamo-Noriega, E.N.; Possani, L.D.; Becerril, B. Updating knowledge on new medically important scorpion species in Mexico. Toxicon 2017, 138, 130–137. [Google Scholar] [CrossRef]
- Carcamo-Noriega, E.N.; Olamendi-Portugal, T.; Restano-Cassulini, R.; Rowe, A.; Uribe-Romero, S.J.; Becerril, B.; Possani, L.D. Intraspecific variation of Centruroides sculpturatus scorpion venom from two regions of Arizona. Arch. Biochem. Biophys. 2018, 638, 52–57. [Google Scholar] [CrossRef]
- Gomez-Ramirez, I.V.; Riano-Umbarila, L.; Olamendi-Portugal, T.; Restano-Cassulini, R.; Possani, L.D.; Becerril, B. Biochemical, electrophysiological and immunological characterization of the venom from Centruroides baergi, a new scorpion species of medical importance in Mexico. Toxicon 2020, 184, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Olamendi-Portugal, T.; Restano-Cassulini, R.; Riano-Umbarila, L.; Becerril, B.; Possani, L.D. Functional and immuno-reactive characterization of a previously undescribed peptide from the venom of the scorpion Centruroides limpidus. Peptides 2017, 87, 34–40. [Google Scholar] [CrossRef]
- Schiavon, E.; Pedraza-Escalona, M.; Gurrola, G.B.; Olamendi-Portugal, T.; Corzo, G.; Wanke, E.; Possani, L.D. Negative-shift activation, current reduction and resurgent currents induced by beta-toxins from Centruroides scorpions in sodium channels. Toxicon 2012, 59, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Boyer, L.; Degan, J.; Ruha, A.M.; Mallie, J.; Mangin, E.; Alagon, A. Safety of intravenous equine F(ab’)2: Insights following clinical trials involving 1534 recipients of scorpion antivenom. Toxicon 2013, 76, 386–393. [Google Scholar] [CrossRef]
- Riano-Umbarila, L.; Juarez-Gonzalez, V.R.; Olamendi-Portugal, T.; Ortiz-Leon, M.; Possani, L.D.; Becerril, B. A strategy for the generation of specific human antibodies by directed evolution and phage display. An example of a single-chain antibody fragment that neutralizes a major component of scorpion venom. FEBS J. 2005, 272, 2591–2601. [Google Scholar] [CrossRef]
- Riano-Umbarila, L.; Gomez-Ramirez, I.V.; Ledezma-Candanoza, L.M.; Olamendi-Portugal, T.; Rodriguez-Rodriguez, E.R.; Fernandez-Taboada, G.; Possani, L.D.; Becerril, B. Generation of a Broadly Cross-Neutralizing Antibody Fragment against Several Mexican Scorpion Venoms. Toxins 2019, 11, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riano-Umbarila, L.; Contreras-Ferrat, G.; Olamendi-Portugal, T.; Morelos-Juarez, C.; Corzo, G.; Possani, L.D.; Becerril, B. Exploiting cross-reactivity to neutralize two different scorpion venoms with one single chain antibody fragment. J. Biol. Chem. 2011, 286, 6143–6151. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Giraldo, A.E.; Olamendi-Portugal, T.; Riano-Umbarila, L.; Becerril, B.; Possani, L.D.; Delepierre, M.; Del Rio-Portilla, F. The three-dimensional structure of the toxic peptide Cl13 from the scorpion Centruroides limpidus. Toxicon 2020, 184, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Pintar, A.; Possani, L.D.; Delepierre, M. Solution structure of toxin 2 from centruroides noxius Hoffmann, a beta-scorpion neurotoxin acting on sodium channels. J. Mol. Biol. 1999, 287, 359–367. [Google Scholar] [CrossRef]
- Saucedo, A.L.; del Rio-Portilla, F.; Picco, C.; Estrada, G.; Prestipino, G.; Possani, L.D.; Delepierre, M.; Corzo, G. Solution structure of native and recombinant expressed toxin CssII from the venom of the scorpion Centruroides suffusus suffusus, and their effects on Nav1.5 sodium channels. Biochim. Biophys. Acta 2012, 1824, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Riano-Umbarila, L.; Rojas-Trejo, V.M.; Romero-Moreno, J.A.; Costas, M.; Utrera-Espindola, I.; Olamendi-Portugal, T.; Possani, L.D.; Becerril, B. Comparative assessment of the VH-VL and VL-VH orientations of single-chain variable fragments of scorpion toxin-neutralizing antibodies. Mol. Immunol. 2020, 122, 141–147. [Google Scholar] [CrossRef]
- Riano-Umbarila, L.; Ledezma-Candanoza, L.M.; Serrano-Posada, H.; Fernandez-Taboada, G.; Olamendi-Portugal, T.; Rojas-Trejo, S.; Gomez-Ramirez, I.V.; Rudino-Pinera, E.; Possani, L.D.; Becerril, B. Optimal Neutralization of Centruroides noxius Venom Is Understood through a Structural Complex between Two Antibody Fragments and the Cn2 Toxin. J. Biol. Chem. 2016, 291, 1619–1630. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Rodriguez, E.R.; Olamendi-Portugal, T.; Serrano-Posada, H.; Arredondo-Lopez, J.N.; Gomez-Ramirez, I.; Fernandez-Taboada, G.; Possani, L.D.; Anguiano-Vega, G.A.; Riano-Umbarila, L.; Becerril, B. Broadening the neutralizing capacity of a family of antibody fragments against different toxins from Mexican scorpions. Toxicon 2016, 119, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Cerni, F.A.; Janke, R.; Bermudez-Mendez, E.; Ledsgaard, L.; Barbosa, J.E.; Laustsen, A.H. History of Envenoming Therapy and Current Perspectives. Front. Immunol. 2019, 10, 1598. [Google Scholar] [CrossRef]
- Knudsen, C.; Ledsgaard, L.; Dehli, R.I.; Ahmadi, S.; Sorensen, C.V.; Laustsen, A.H. Engineering and design considerations for next-generation snakebite antivenoms. Toxicon 2019, 167, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, C.; Casewell, N.R.; Lomonte, B.; Gutierrez, J.M.; Vaiyapuri, S.; Laustsen, A.H. Novel Snakebite Therapeutics Must Be Tested in Appropriate Rescue Models to Robustly Assess Their Preclinical Efficacy. Toxins 2020, 12, 528. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Maria Gutierrez, J.; Knudsen, C.; Johansen, K.H.; Bermudez-Mendez, E.; Cerni, F.A.; Jurgensen, J.A.; Ledsgaard, L.; Martos-Esteban, A.; Ohlenschlaeger, M.; et al. Pros and cons of different therapeutic antibody formats for recombinant antivenom development. Toxicon 2018, 146, 151–175. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger Release 2021-1; Maestro, Schrödinger, LLC: New York, NY, USA, 2021.
- Bowers, K.J.; Chow, D.E.; Xu, H.; Dror, R.O.; Eastwood, M.P.; Gregersen, B.A.; Klepeis, J.L.; Kolossvary, I.; Moraes, M.A.; Sacerdoti, F.D.; et al. Scalable algorithms for molecular dynamics simulations on commodity clusters. In Proceedings of the SC ‘06: Proceedings of the 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 11–17 November 2006; pp. 1–13. [Google Scholar]
- Tina, K.G.; Bhadra, R.; Srinivasan, N. PIC: Protein Interactions Calculator. Nucleic Acids Res. 2007, 35, W473–W476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| scFv | Toxin | kon (M−1s−1) × 105 | koff (s−1) × 10−4 | KD (M) | TR (min) |
|---|---|---|---|---|---|
| LR | CsEd | 3.85 | 2.35 | 6.1 × 10−10 | 70.9 |
| CsEM1a | 6.28 | 7.9 | 1.29 × 10−9 | 21 | |
| 10FG2 | CsEd | 1.85 | 3.4 | 1.84 × 10−9 | 49 |
| CsEM1a | 2.37 | 2.61 | 1.1 × 10−9 | 63 |
| scFv(s) | LD50 of Venom | Molar Ratio | Survivors/Total |
|---|---|---|---|
| LR | 1 | 1:10 | 9/10 ** |
| LR | 2 | 1:10 | 3/6 *** |
| 10FG2 | 2 | 1:10 | 6/6 * |
| LR + 10FG2 | 2 | 1:5 of each scFv | 6/6 |
| Control | 1 | - | 5/10 *** |
| Control | 2 | - | 0/6 *** |
| Survivors/Total | ||||
|---|---|---|---|---|
| Mix assays scFv(s) | LD/Molar ratios (toxin:scFv) | |||
| 2 LD50 1:5 | 3 LD50 1:3.3 | 4 LD50 1:2.5 | 5 LD50 1:2 | |
| 10FG2 | 6/6 | 6/6 | 6/6 * | 6/6 * |
| Comb. LR+10FG2 | - | - | 6/6 | 6/6 |
| Control 2 LD50 | 0/6 | |||
| Comb. LR+10FG2 | 6/6 | |||
| Control | 0/6 | |||
| Survivors/Total | |
|---|---|
| Comb. LR+10FG2 | 6/6 |
| Control | 0/6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riaño-Umbarila, L.; Romero-Moreno, J.A.; Ledezma-Candanoza, L.M.; Olamendi-Portugal, T.; Possani, L.D.; Becerril, B. Full Neutralization of Centruroides sculpturatus Scorpion Venom by Combining Two Human Antibody Fragments. Toxins 2021, 13, 708. https://doi.org/10.3390/toxins13100708
Riaño-Umbarila L, Romero-Moreno JA, Ledezma-Candanoza LM, Olamendi-Portugal T, Possani LD, Becerril B. Full Neutralization of Centruroides sculpturatus Scorpion Venom by Combining Two Human Antibody Fragments. Toxins. 2021; 13(10):708. https://doi.org/10.3390/toxins13100708
Chicago/Turabian StyleRiaño-Umbarila, Lidia, José Alberto Romero-Moreno, Luis M. Ledezma-Candanoza, Timoteo Olamendi-Portugal, Lourival D. Possani, and Baltazar Becerril. 2021. "Full Neutralization of Centruroides sculpturatus Scorpion Venom by Combining Two Human Antibody Fragments" Toxins 13, no. 10: 708. https://doi.org/10.3390/toxins13100708
APA StyleRiaño-Umbarila, L., Romero-Moreno, J. A., Ledezma-Candanoza, L. M., Olamendi-Portugal, T., Possani, L. D., & Becerril, B. (2021). Full Neutralization of Centruroides sculpturatus Scorpion Venom by Combining Two Human Antibody Fragments. Toxins, 13(10), 708. https://doi.org/10.3390/toxins13100708