Association of Sarcopenia and Gut Microbiota Composition in Older Patients with Advanced Chronic Kidney Disease, Investigation of the Interactions with Uremic Toxins, Inflammation and Oxidative Stress

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Population Characteristics
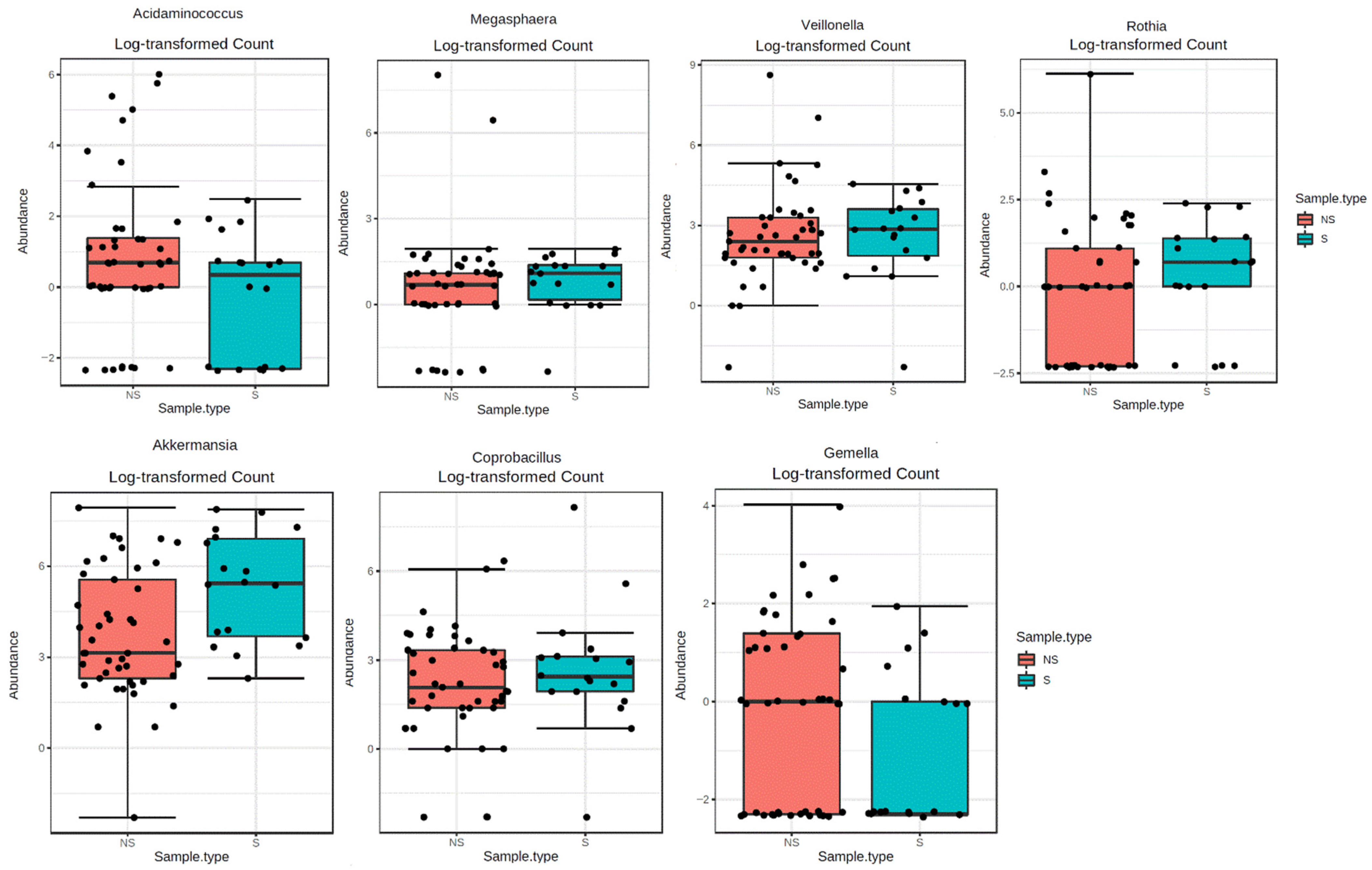
2.2. Sarcopenia and gMB Composition
2.2.1. Correlations of gMB with Uremic Toxins
2.2.2. Correlations of gMB with Inflammatory Cytokines
2.2.3. Correlations of gMB with Oxidative stress
2.3. Sarcopenia, Uremic Toxins, Oxidative Stress and Inflammatory Parameters
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Population Characteristics
5.2. Measurement of Uremic Toxins and Malondialdehyde
5.3. Measurement of Serum Cytokines
5.4. Bacterial DNA Extraction and V3–V4 Region Sequencing
5.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, H.; Enoki, Y.; Maruyama, T. Sarcopenia in chronic kidney disease: Factors, mechanisms, and therapeutic interventions. Biol. Pharm. Bull. 2019, 42, 1437–1445. [Google Scholar] [CrossRef] [Green Version]
- Nataatmadja, M.; Cho, Y.; Campbell, K.; Johnson, D.W. The roles of indoxyl sulphate and p-cresyl sulphate in patients with chronic kidney disease: A review of therapeutic options. In Chronic Kidney Disease—From Pathophysiology to Clinical Improvements; Rath, T., Ed.; InTech: London, UK, 2018. [Google Scholar]
- Mair, R.D.; Sirich, T.L.; Plummer, N.S.; Meyer, T.W. Characteristics of colon-derived uremic solutes. Clin. J. Am. Soc. Nephrol. 2018, 13, 1398–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evenepoel, P.; Meijers, B.; Bammens, B.; Verbeke, K. Uremic toxins originating from colonic microbial metabolism. Kidney Int. 2009, 76, S12–S19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreto, F.C.; Barreto, D.V.; Liabeuf, S.; Meert, N.; Glorieux, G.; Temmar, M.; Choukroun, G.; Vanholder, R.; Massy, Z.A.; European Uremic Toxin Work Group (EUTox). Serum indoxyl sulfate is associated with vascular disease and mortality in chronic kidney disease patients. Clin. J. Am. Soc. Nephrol. 2009, 4, 1551–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koppe, L.; Pillon, N.; Vella, R.E.; Croze, M.L.; Pelletier, C.C.; Chambert, S.; Massy, Z.; Glorieux, G.; Vanholder, R.; Dugenet, Y.; et al. P-cresyl sulfate promotes insulin resistance associated with CKD. J. Am. Soc. Nephrol. 2012, 24, 88–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liabeuf, S.; Barreto, D.V.; Barreto, F.C.; Meert, N.; Glorieux, G.; Schepers, E.; Temmar, M.; Choukroun, G.; Vanholder, R.; Massy, Z.A.; et al. Free p-cresyl sulphate is a predictor of mortality in patients at different stages of chronic kidney disease. Nephrol. Dial. Transplant. 2009, 25, 1183–1191. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Tsuruoka, S.; Ioka, T.; Ando, H.; Ito, C.; Akimoto, T.; Fujimura, A.; Asano, Y.; Kusano, E. Indoxyl sulfate stimulates proliferation of rat vascular smooth muscle cells. Kidney Int. 2006, 69, 1780–1785. [Google Scholar] [CrossRef] [Green Version]
- Adijiang, A.; Goto, S.; Uramoto, S.; Nishijima, F.; Niwa, T. Indoxyl sulphate promotes aortic calcification with expression of osteoblast-specific proteins in hypertensive rats. Nephrol. Dial. Transplant. 2008, 23, 1892–1901. [Google Scholar] [CrossRef] [Green Version]
- Sato, E.; Mori, T.; Mishima, E.; Suzuki, A.; Sugawara, S.; Kurasawa, N.; Saigusa, D.; Miura, D.; Morikawa-Ichinose, T.; Saito, R.; et al. Metabolic alterations by indoxyl sulfate in skeletal muscle induce uremic sarcopenia in chronic kidney disease. Sci. Rep. 2016, 6, 36618. [Google Scholar] [CrossRef]
- Changchien, C.-Y.; Lin, Y.-H.; Cheng, Y.-C.; Chang, H.-H.; Peng, Y.-S.; Chen, Y. Indoxyl sulfate induces myotube atrophy by ROS-ERK and JNK-MAFbx cascades. Chem. Interact. 2019, 304, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Caldiroli, L.; Armelloni, S.; Eskander, A.; Messa, P.; Rizzo, V.; Margiotta, E.; Cesari, M.; Vettoretti, S. Association between the uremic toxins indoxyl-sulfate and p-cresyl-sulfate with sarcopenia and malnutrition in elderly patients with advanced chronic kidney disease. Exp. Gerontol. 2021, 147, 111266. [Google Scholar] [CrossRef]
- Briskey, D.; Tucker, P.; Johnson, D.W.; Coombes, J. The role of the gastrointestinal tract and microbiota on uremic toxins and chronic kidney disease development. Clin. Exp. Nephrol. 2017, 21, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Andersen, K.; Kesper, M.S.; Marschner, J.A.; Konrad, L.; Ryu, M.; Vr, S.K.; Kulkarni, O.P.; Mulay, S.R.; Romoli, S.; Dem-leitner, J.; et al. Intestinal dysbiosis, barrier dysfunction, and bacterial translocation account for CKD-related systemic in-flammation. J. Am. Soc. Nephrol. 2017, 28, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margiotta, E.; Miragoli, F.; Callegari, M.L.; Vettoretti, S.; Caldiroli, L.; Meneghini, M.; Zanoni, F.; Messa, P. Gut microbiota composition and frailty in elderly patients with chronic kidney disease. PLoS ONE 2020, 15, e0228530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vettoretti, S.; Caldiroli, L.; Armelloni, S.; Ferrari, C.; Cesari, M.; Messa, P. Sarcopenia is associated with malnutrition but not with systemic inflammation in older persons with advanced CKD. Nutrients 2019, 11, 1378. [Google Scholar] [CrossRef] [Green Version]
- Ntemiri, A. Effect of Dairy Based Food Ingredients on the Gut Microbiota of Older Consumers. Ph.D. Thesis, University College Cork, Cork, Ireland, 2019. [Google Scholar]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.A.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef] [Green Version]
- Myers, B.; Brownstone, N.; Reddy, V.; Chan, S.; Thibodeaux, Q.; Truong, A.; Bhutani, T.; Chang, H.-W.; Liao, W. The gut microbiome in psoriasis and psoriatic arthritis. Best Pract. Res. Clin. Rheumatol. 2019, 33, 101494. [Google Scholar] [CrossRef]
- Merli, M.; Iebba, V.; Giusto, M. What is new about diet in hepatic encephalopathy. Metab. Brain Dis. 2016, 31, 1289–1294. [Google Scholar] [CrossRef]
- Yang, X.; Feng, H.; Zhan, X.; Zhang, C.; Cui, R.; Zhong, L.; Ying, S.; Chen, Z. Early-life vancomycin treatment promotes airway inflammation and impairs microbiome homeostasis. Aging 2019, 11, 2071–2081. [Google Scholar] [CrossRef]
- Biagi, E.; Nylund, L.; Candela, M.; Ostan, R.; Bucci, L.; Pini, E.; Nikkïla, J.; Monti, D.; Satokari, R.; Franceschi, C.; et al. Through ageing, and beyond: Gut microbiota and inflammatory status in seniors and centenarians. PLoS ONE 2010, 5, e10667. [Google Scholar] [CrossRef]
- Kaczmarek, J.L.; Musaad, S.M.; Holscher, H.D. Time of day and eating behaviors are associated with the composition and function of the human gastrointestinal microbiota. Am. J. Clin. Nutr. 2017, 106, ajcn156380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, M.A.; Jeffery, I.B.; Beaumont, M.; Bell, J.T.; Clark, A.G.; Ley, R.E.; O’Toole, P.W.; Spector, T.D.; Steves, C.J. Signatures of early frailty in the gut microbiota. Genome Med. 2016, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Maffei, V.J.; Kim, S.; Blanchard, E.; Luo, M.; Jazwinski, S.M.; Taylor, C.; Welsh, D.A. Biological aging and the human gut microbiota. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2017, 72, 1474–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzo, S.; Mulè, F.; Caldara, G.F.; Baldassano, S.; Puleio, R.; Vitale, M.; Cassata, G.; Ferrantelli, V.; Amato, A. Pistachio consumption alleviates inflammation and improves gut microbiota composition in mice fed a high-fat diet. Int. J. Mol. Sci. 2020, 21, 365. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, L.; Abete, P.; Bellelli, G.; Bo, M.; Cherubini, A.; Corica, F.; Di Bari, M.; Maggio, M.; Manca, G.M.; Rizzo, M.R.; et al. Prevalence and clinical correlates of sarcopenia, identified according to the EWGSOP definition and diagnostic algorithm, in hospitalized older people: The GLISTEN study. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2017, 72, 1575–1581. [Google Scholar] [CrossRef]
- Peters, B.A.; Shapiro, J.A.; Church, T.R.; Miller, G.; Trinh-Shevrin, C.; Yuen, E.; Friedlander, C.; Hayes, R.B.; Ahn, J. A taxonomic signature of obesity in a large study of American adults. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Barlow, G.M.; Lin, E.A.; Mathur, R. An overview of the roles of the gut microbiome in obesity and diabetes. In Nutritional and Therapeutic Interventions for Diabetes and Metabolic Syndrome; Bagchi, D., Ed.; Elsevier: London, UK, 2018; pp. 65–91. [Google Scholar]
- Dhakal, S.; McCormack, L.; Dey, M. Association of the gut microbiota with weight-loss response within a retail weight-management program. Microorganisms 2020, 8, 1246. [Google Scholar] [CrossRef]
- Freedman, S.N.; Shahi, S.K.; Mangalam, A.K. The “gut feeling”: Breaking down the role of gut microbiome in multiple sclerosis. Neurotherapeutics 2018, 15, 109–125. [Google Scholar] [CrossRef] [Green Version]
- Rogosa, M. Acidaminococcus gen. n., Acidaminococcus fermentans sp. n., anaerobic gram-negative diplococci using amino acids as the sole energy source for growth. J. Bacteriol. 1969, 98. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Wu, K.; Pan, W.; Zeng, Y.; Hu, K.; Chen, D.; Huang, X.; Zhang, Q. Intestinal flora alterations in patients with early chronic kidney disease: A case-control study among the Han population in southwestern China. J. Int. Med. Res. 2020, 48. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhang, Y.; Wang, X.; Yang, R.; Zhu, X.; Zhang, Y.; Chen, C.; Yuan, H.; Yang, Z.; Sun, L. Gut bacteria Akkermansia is associated with reduced risk of obesity: Evidence from the American gut project. Nutr. Metab. 2020, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pisanu, S.; Palmas, V.; Madau, V.; Casula, E.; Deledda, A.; Cusano, R.; Uva, P.; Vascellari, S.; Boi, F.; Loviselli, A.; et al. Impact of a moderately hypocaloric Mediterranean diet on the gut microbiota composition of Italian obese patients. Nutrients 2020, 12, 2707. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Wong, J.; Pahl, M.; Piceno, Y.; Yuan, J.; DeSantis, T.Z.; Ni, Z.; Nguyen, T.-H.; Andersen, G.L. Chronic kidney disease alters intestinal microbial flora. Kidney Int. 2013, 83, 308–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gryp, T.; Huys, G.R.; Joossens, M.; Van Biesen, W.; Glorieux, G.; Vaneechoutte, M. Isolation and quantification of uremic toxin precursor-generating gut bacteria in chronic kidney disease patients. Int. J. Mol. Sci. 2020, 21, 1986. [Google Scholar] [CrossRef] [Green Version]
- Brook, I. Anaerobic infections in childhood. Clin. Infect. Dis. 1984, 6, S187–S192. [Google Scholar] [CrossRef]
- Zhang, L.S.; Davies, S.S. Microbial metabolism of dietary components to bioactive metabolites: Opportunities for new therapeutic interventions. Genome Med. 2016, 8, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Barrios, C.; Beaumont, M.; Pallister, T.; Villar, J.; Goodrich, J.K.; Clark, A.; Pascual, J.; Ley, R.; Spector, T.D.; Bell, J.T.; et al. Gut-microbiota-metabolite axis in early renal function decline. PLoS ONE 2015, 10, e0134311. [Google Scholar] [CrossRef] [Green Version]
- Terrier-Lenglet, A.; Nollet, A.; Liabeuf, S.; Barreto, D.V.; Brazier, M.; Lemke, H.D.; Vanholder, R.; Choukroun, G.; Massy, Z.A.; Groupe EUTox (European Uremic toxin). Plasma malondialdehyde may not predict mortality in patient with chronic kidney disease. Nephrol. Ther. 2011, 7, 219–224. [Google Scholar] [CrossRef]
- Xu, K.-Y.; Xia, G.-H.; Lu, J.-Q.; Chen, M.-X.; Zhen, X.; Wang, S.; You, C.; Nie, J.; Zhou, H.-W.; Yin, J. Impaired renal function and dysbiosis of gut microbiota contribute to increased trimethylamine-N-oxide in chronic kidney disease patients. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Schippa, S.; Iebba, V.; Santangelo, F.; Gagliardi, A.; De Biase, R.V.; Stamato, A.; Bertasi, S.; Lucarelli, M.; Conte, M.P.; Quattrucci, S.; et al. Cystic fibrosis transmembrane conductance regulator (CFTR) allelic variants relate to shifts in faecal microbiota of cystic fibrosis patients. PLoS ONE 2013, 8, e61176. [Google Scholar] [CrossRef]
- Waters, J.L.; Ley, R.E. The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 2019, 17, 1–11. [Google Scholar] [CrossRef]
- Feng, Y.-L.; Cao, G.; Chen, D.-Q.; Vaziri, N.D.; Chen, L.; Zhang, J.; Wang, M.; Guo, Y.; Zhao, Y.-Y. Microbiome-metabolomics reveals gut microbiota associated with glycine-conjugated metabolites and polyamine metabolism in chronic kidney disease. Cell. Mol. Life Sci. 2019, 76, 4961–4978. [Google Scholar] [CrossRef] [Green Version]
- Saito, Y.; Sato, T.; Nomoto, K.; Tsuji, H. Identification of phenol- and p-cresol-producing intestinal bacteria by using media supplemented with tyrosine and its metabolites. FEMS Microbiol. Ecol. 2018, 76, 4961–4978. [Google Scholar] [CrossRef]
- Carson, T.; Lowe, J.; Ramaraj, T.; Leff, J.W.; Luo, L.; Bell, C.J.; Shah, V.O. Composition diversity and abundance of gut microbiome in prediabetes and type 2 diabetes. J. Diabetes Obes. 2015, 2, 1–7. [Google Scholar]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. 2016, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Liu, S.; Zhou, Y.; Yao, Z.; Zhang, D.; Cao, S.; Wei, Z.; Tan, B.; Li, Y.; Lian, Z.; et al. Evolutionary biologic changes of gut microbiota in an “adenoma-carcinoma sequence” mouse colorectal cancer model induced by 1, 2-Dimethylhydrazine. Oncotarget 2016, 8, 444–457. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Lv, L.; Wu, W.; Li, Y.; Shi, D.; Fang, D.; Guo, F.; Jiang, H.; Yan, R.; Ye, W.; et al. Butyrate protects mice against methionine–choline-deficient diet-induced non-alcoholic steatohepatitis by improving gut barrier function, attenuating inflammation and reducing endotoxin levels. Front. Microbiol. 2018, 9, 1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masood, M.I.; Qadir, M.I.; Shirazi, J.H.; Khan, I.U. Beneficial effects of lactic acid bacteria on human beings. Crit. Rev. Microbiol. 2010, 37, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Biagi, E.; Franceschi, C.; Rampelli, S.; Severgnini, M.; Ostan, R.; Turroni, S.; Consolandi, C.; Quercia, S.; Scurti, M.; Monti, D.; et al. Gut microbiota and extreme longevity. Curr. Biol. 2016, 26, 1480–1485. [Google Scholar] [CrossRef] [Green Version]
- Vörös, K.; Szenthe, P.; Kaszás, E.; Cseh, K.; Gráf, L., Jr.; Prohászka, Z.; Gráf, L.; Böröcz, Z.; Kalabay, L. Serum fetuin-A in metabolic and inflammatory pathways in patients with myocardial infarction. Eur. J. Clin. Investig. 2011, 41, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, G.A.; Barrett-Connor, E.; Cummins, K.M.; Daniels, L.B.; Wassel, C.L.; Ix, J.H. Sex-specific association of fetuin-a with type 2 diabetes in older community-dwelling adults: The Rancho Bernardo study. Diabetes Care 2013, 36, 1994–2000. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. Human gut microbiota changes reveal the progression of glucose intolerance. PLoS ONE 2013, 8, e71108. [Google Scholar] [CrossRef] [PubMed]
- Ix, J.H.; Wassel, C.L.; Kanaya, A.M.; Vittinghoff, E.; Johnson, K.C.; Koster, A.; Cauley, J.A.; Harris, T.B.; Cummings, S.R.; Shlipak, M.J. Fetuin-A and incident diabetes mellitus in older persons. JAMA 2008, 300, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-T.N. Urea concentration in intestinal fluids in normal and uremic dogs. J. Surg. Oncol. 1971, 3, 163–168. [Google Scholar] [CrossRef]
- Pretorius, C.J.; McWhinney, B.C.; Sipinkoski, B.; Johnson, L.A.; Rossi, M.; Campbell, K.L.; Ungerer, J.P. Reference ranges and biological variation of free and total serum indoxyl- and p-cresyl sulphate measured with a rapid UPLC fluorescence detection method. Clin. Chim. Acta 2013, 419, 122–126. [Google Scholar] [CrossRef]
- Grune, T.; Siems, W.; Esterbauer, H. 077 comparison of different assays for malondialdehyde using thiobarbituric acid. Anal. Bioanal. Chem. 1992, 343, 135. [Google Scholar] [CrossRef]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variables | Overall Cohort (n = 63) | Sarcopenic (n = 18) | Not Sarcopenic (n = 45) | p |
|---|---|---|---|---|
| Age (years) | 80.7 ± 6.2 | 83.1 ± 5.7 | 79.7 ± 6.2 | 0.0528 |
| Males, n (%) | 44 (69) | 16 (89) | 28 (62) | 0.12 |
| eGFR (ml/min/1,73 m2) | 26 ± 11 | 23.9 ± 10 | 25.5 ± 10 | 0.5733 |
| Diabetes, n (%) | 37 (59%) | 11 (61%) | 26 (58%) | 0.75 |
| Frailty, n (%) | 38 (59) | 13 (72) | 24 (53) | 0.16 |
| BMI (kg/m2) | 28.4 ± 4.7 | 25.5 ± 2.6 | 29.3 ± 4.8 | 0.025 |
| MIS | 6 ± 4.7 | 6.7 ± 4.2 | 5.7 ± 4.9 | 0.48 |
| CRP, (mg/dl) | 0.3 ± 0.5 | 0.43 ± 0.9 | 0.28 ± 0.22 | 0.20 |
| HCO3– | 24.0 ± 3.5 | 24.0 ± 3.6 | 23.9 ± 2.9 | 0.93 |
| GENUS | Uremic Toxins | Anti-Inflammatory Cytokines | Pro-Inflammatory Cytokines | Oxidative Stress | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| IS µmol/L | PCs µmol/L | IL-10 pg/mL | IL-12 pg/mL | Fetuin A ng/mL | IL-6 pg/mL | IL-17 pg/mL | TNF-α pg/mL | MCP-1 pg/mL | MDA µmol/L | ||
| Alistipes | r p | 0.01 0.95 | 0.11 0.38 | −0.05 0.70 | −0.25 0.048 | −0.17 0.17 | 0.048 0.71 | −0.23 0.07 | 0.077 0.54 | −0.19 0.13 | 0.12 0.36 |
| Bacteroides | r p | 0.06 0.64 | −0.1 0.44 | −0.22 0.086 | −0.04 0.76 | −0.035 0.78 | 0.23 0.06 | 0.27 0.033 | −0.29 0.019 | 0.11 0.40 | |
| Collinsella | r p | 0.15 0.25 | 0.05 0.70 | 0.16 0.21 | −0.039 0.76 | 0.25 0.045 | −0.09 0.47 | 0.31 0.013 | −0.14 0.27 | 0.023 0.86 | 0.021 0.87 |
| Desulfovibrio | r p | −0.025 0.84 | 0.29 0.02 | −0.056 0.66 | 0.045 0.73 | 0.15 0.25 | −0.11 0.39 | −0.038 0.77 | −0.09 0.498 | 0.095 0.45 | 0.046 0.72 |
| Eggerthella | r p | 0.007 0.95 | −0.09 0.50 | −0.13 0.29 | −0.26 0.04 | −0.08 0.50 | −0.07 0.69 | −0.04 0.74 | 0.16 0.21 | −0.29 0.022 | 0.17 0.18 |
| Lactobacillus | r p | 0.39 0.002 | −0.07 0.58 | 0.17 0.18 | 0.049 0.71 | 0.11 0.40 | −0.08 0.55 | −0.05 0.72 | −0.34 0.006 | 0.08 0.51 | −0.07 0.60 |
| Ruminococcus | r p | −0.007 0.96 | 0.09 0.49 | −0.12 0.34 | −0.18 0.15 | 0.095 0.45 | 0.02 0.86 | −0.15 0.25 | 0.08 0.53 | −0.33 0.007 | 0.17 0.19 |
| Streptococcus | r p | 0.18 0.16 | 0.06 0.64 | 0.17 0.17 | −0.19 0.14 | 0.14 0.26 | −0.08 0.54 | 0.05 0.71 | −0.27 0.03 | −0.24 0.06 | −0.15 0.24 |
| Unclass. Barnesiellaceae | r p | −0.015 0.91 | −0.007 0.96 | −0.06 0.64 | 0.04 0.77 | −0.015 0.91 | 0.14 0.29 | −0.12 0.34 | 0.3 0.016 | −0.34 0.0066 | 0.17 0.19 |
| Unclass. Coriobacteriaceae | r p | 0.16 0.22 | 0.29 0.02 | 0.11 0.37 | 0.013 0.92 | 0.16 0.20 | −0.15 0.23 | −0.09 0.49 | −0.067 0.60 | −0.15 0.24 | −0.053 0.68 |
| Unclass. Christensenellaceae | r p | 0.08 0.54 | 0.28 0.027 | −0.13 0.29 | 0.027 0.83 | 0.037 0.77 | 0.025 0.84 | −0.21 0.11 | 0.0052 0.97 | −0.091 0.47 | −0.052 0.69 |
| Unclass. Mogibacteriaceae | r p | 0.11 0.41 | 0.26 0.04 | 0.18 0.16 | −0.17 0.18 | 0.053 0.68 | −0.081 0.52 | −0.12 0.37 | 0.033 0.80 | −0.071 0.58 | 0.065 0.61 |
| Unclass. Rikenellaceae | r p | 0.061 0.64 | 0.1 0.44 | −0.15 0.23 | −0.1 0.43 | −0.07 0.59 | 0.095 0.45 | −0.15 0.25 | 0.27 0.03 | −0.2 0.11 | 0.18 0.15 |
| SPECIES | Uremic Toxins | Anti-Inflammatory Cytokines | Pro-Inflammatory Cytokines | Oxidative Stress | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| IS µmol/L | PCs µmol/L | IL-10 pg/mL | IL-12 pg/mL | Fetuin-A ng/mL | IL-6 pg/mL | IL-17 pg/mL | TNF-α pg/mL | MCP-1 pg/mL | MDA µmol/L | ||
| Lactobacillus Unclass.Lactobacillus | r p | 0.42 0.0005 | −0.04 0.76 | 0.21 0.10 | 0.098 0.45 | 0.2 0.12 | −0.11 0.39 | −0.02 0.88 | −0.3 0.016 | 0.09 0.48 | −0.042 0.74 |
| Bifidobacterium-longum | r p | 0.25 0.047 | 0.012 0.93 | 0.11 0.39 | −0.19 0.15 | 0.16 0.21 | −0.12 0.35 | 0.0056 0.97 | 0.11 0.40 | −0.12 0.35 | 0.092 0.47 |
| Eubacterium-biforme | r p | 0.061 0.64 | 0.35 0.006 | 0.12 0.33 | −0.043 0.74 | 0.13 0.32 | −0.016 0.89 | −0.24 0.06 | −0.019 0.88 | 0.11 0.39 | −0.089 0.49 |
| Oscillospira | r p | 0.18 0.16 | 0.28 0.029 | 0.083 0.51 | −0.096 0.46 | 0.017 0.90 | 0.079 0.54 | −0.034 0.80 | 0.087 0.49 | −0.14 0.28 | 0.045 0.72 |
| Bacteroides-Unclass. Bacteroides | r p | 0.056 0.66 | −0.1 0.43 | −0.26 0.04 | −0.019 0.88 | 0.00004 0.99 | 0.29 0.02 | −0.15 0.26 | 0.28 0.026 | −0.26 0.042 | 0.093 0.47 |
| Collinsella-aerofaciens | r p | 0.12 0.37 | 0.092 0.48 | 0.15 0.23 | 0.01 0.94 | 0.28 0.02 | −0.12 0.36 | 0.28 0.028 | −0.14 0.27 | 0.07 0.58 | 0.052 0.69 |
| Clostridium-difficile | r p | 0.13 0.30 | 0.059 0.65 | 0.14 0.27 | −0.095 0.46 | 0.2 0.12 | −0.024 0.85 | −0.099 0.45 | −0.058 0.65 | 0.054 0.67 | −0.16 0.22 |
| Unclass. Christensenellaceae | r p | 0.074 0.57 | 0.29 0.021 | −0.15 0.24 | 0.034 0.79 | 0.037 0.77 | 0.03 0.82 | −0.2 0.13 | −0.001 0.99 | −0.064 0.61 | −0.052 0.68 |
| Unclass. Clostridiaceae | r p | 0.21 0.098 | 0.34 0.007 | −0.01 0.94 | −0.04 0.76 | 0.15 0.22 | 0.064 0.62 | −0.22 0.096 | 0.045 0.73 | −0.018 0.88 | −0.09 0.46 |
| Unclass. Mogibacteriaceae | rp | 0.1 0.44 | 0.29 0.023 | 0.15 0.22 | −0.16 0.21 | 0.054 0.67 | −0.072 0.57 | −0.11 0.40 | 0.023 0.86 | −0.038 0.76 | 0.063 0.62 |
| Variables | Sarcopenic (n = 18) | Not Sarcopenic (n = 45) | p |
|---|---|---|---|
| Uremic toxins | |||
| PCs (µmol/L) | 0.65 ± 0 | 0.79 ± 0.18 | 0.24 |
| IS (µmol/L) | 1.83 ± 2.61 | 2.2 ± 3.13 | 0.43 |
| Pro-Inflammatory cytokines | |||
| IL-6 (pg/mL) | 2.4 ± 4.93 | 2.88 ± 4.83 | 0.70 |
| TNFα (pg/mL) | 14.63 ± 8.84 | 14.14 ± 8.65 | 0.99 |
| MCP-1 (pg/mL) | 428.88 ± 316.49 | 447.17 ± 199.71 | 0.36 |
| IL17 (pg/mL) | 2.4 ± 4.93 | 2.88 ± 4.83 | 0.70 |
| Anti-Inflammatory cytokines | |||
| IL12p70 (pg/mL) | 0.605 ± 1.80 | 1.471 ± 2.13 | 0.45 |
| IL-10 (pg/mL) | 1.08 ± 1.66 | 2.4 ± 5.87 | 0.039 |
| Fetuin-A (ng/mL) | 0.56 ± 0.65 | 0.64 ± 0.56 | 0.56 |
| Oxidative Stress | |||
| MDA (µmol/L) | 0.275 ± 0.31 | 0.346 ± 0.18 | 0.47 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Margiotta, E.; Caldiroli, L.; Callegari, M.L.; Miragoli, F.; Zanoni, F.; Armelloni, S.; Rizzo, V.; Messa, P.; Vettoretti, S. Association of Sarcopenia and Gut Microbiota Composition in Older Patients with Advanced Chronic Kidney Disease, Investigation of the Interactions with Uremic Toxins, Inflammation and Oxidative Stress. Toxins 2021, 13, 472. https://doi.org/10.3390/toxins13070472
Margiotta E, Caldiroli L, Callegari ML, Miragoli F, Zanoni F, Armelloni S, Rizzo V, Messa P, Vettoretti S. Association of Sarcopenia and Gut Microbiota Composition in Older Patients with Advanced Chronic Kidney Disease, Investigation of the Interactions with Uremic Toxins, Inflammation and Oxidative Stress. Toxins. 2021; 13(7):472. https://doi.org/10.3390/toxins13070472
Chicago/Turabian StyleMargiotta, Elisabetta, Lara Caldiroli, Maria Luisa Callegari, Francesco Miragoli, Francesca Zanoni, Silvia Armelloni, Vittoria Rizzo, Piergiorgio Messa, and Simone Vettoretti. 2021. "Association of Sarcopenia and Gut Microbiota Composition in Older Patients with Advanced Chronic Kidney Disease, Investigation of the Interactions with Uremic Toxins, Inflammation and Oxidative Stress" Toxins 13, no. 7: 472. https://doi.org/10.3390/toxins13070472
APA StyleMargiotta, E., Caldiroli, L., Callegari, M. L., Miragoli, F., Zanoni, F., Armelloni, S., Rizzo, V., Messa, P., & Vettoretti, S. (2021). Association of Sarcopenia and Gut Microbiota Composition in Older Patients with Advanced Chronic Kidney Disease, Investigation of the Interactions with Uremic Toxins, Inflammation and Oxidative Stress. Toxins, 13(7), 472. https://doi.org/10.3390/toxins13070472