Toxicity of the Diatom Genus Pseudo-nitzschia (Bacillariophyceae): Insights from Toxicity Tests and Genetic Screening in the Northern Adriatic Sea



Abstract
:1. Introduction
2. Results
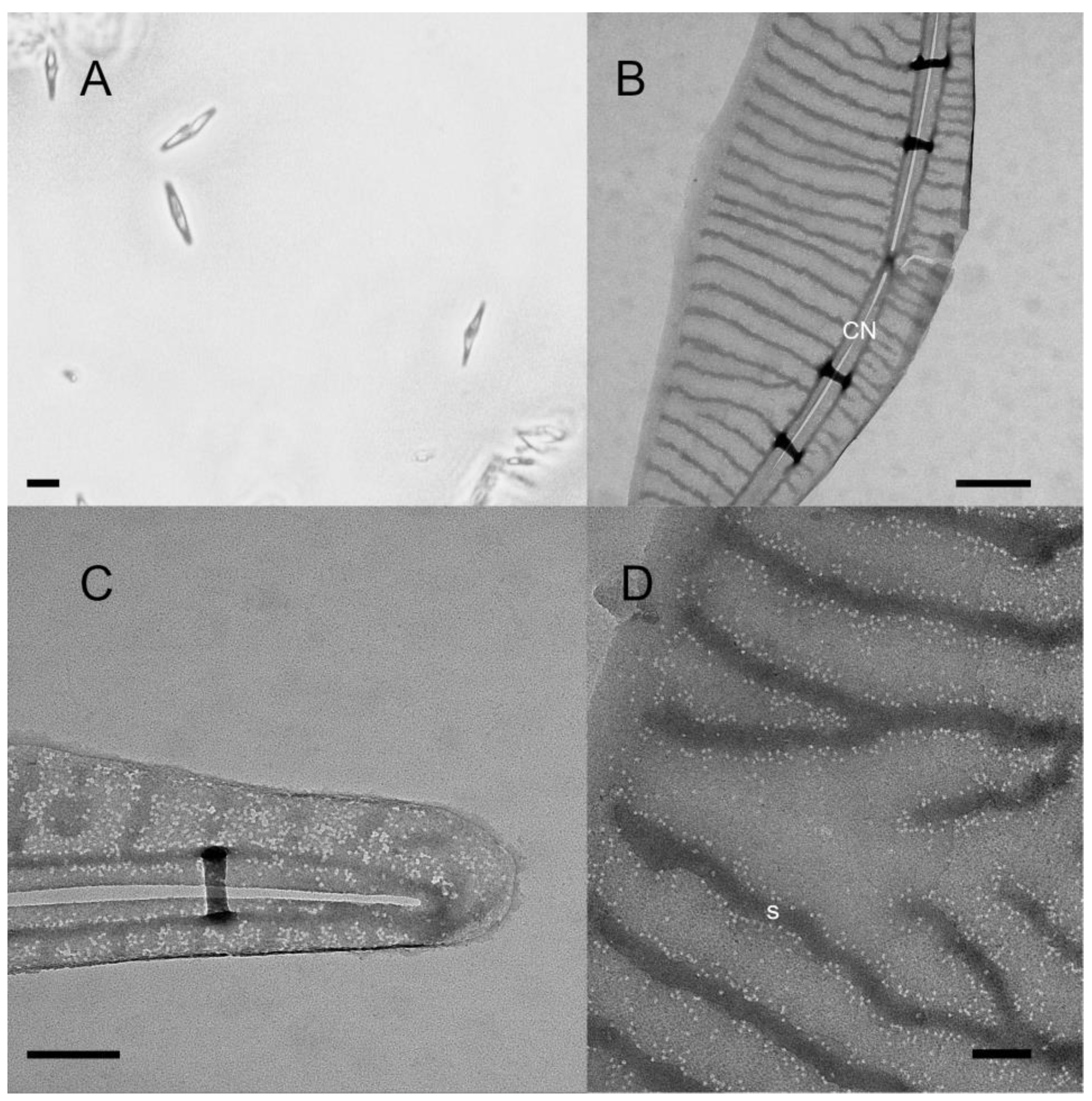
2.1. Toxicity of Individual Strains
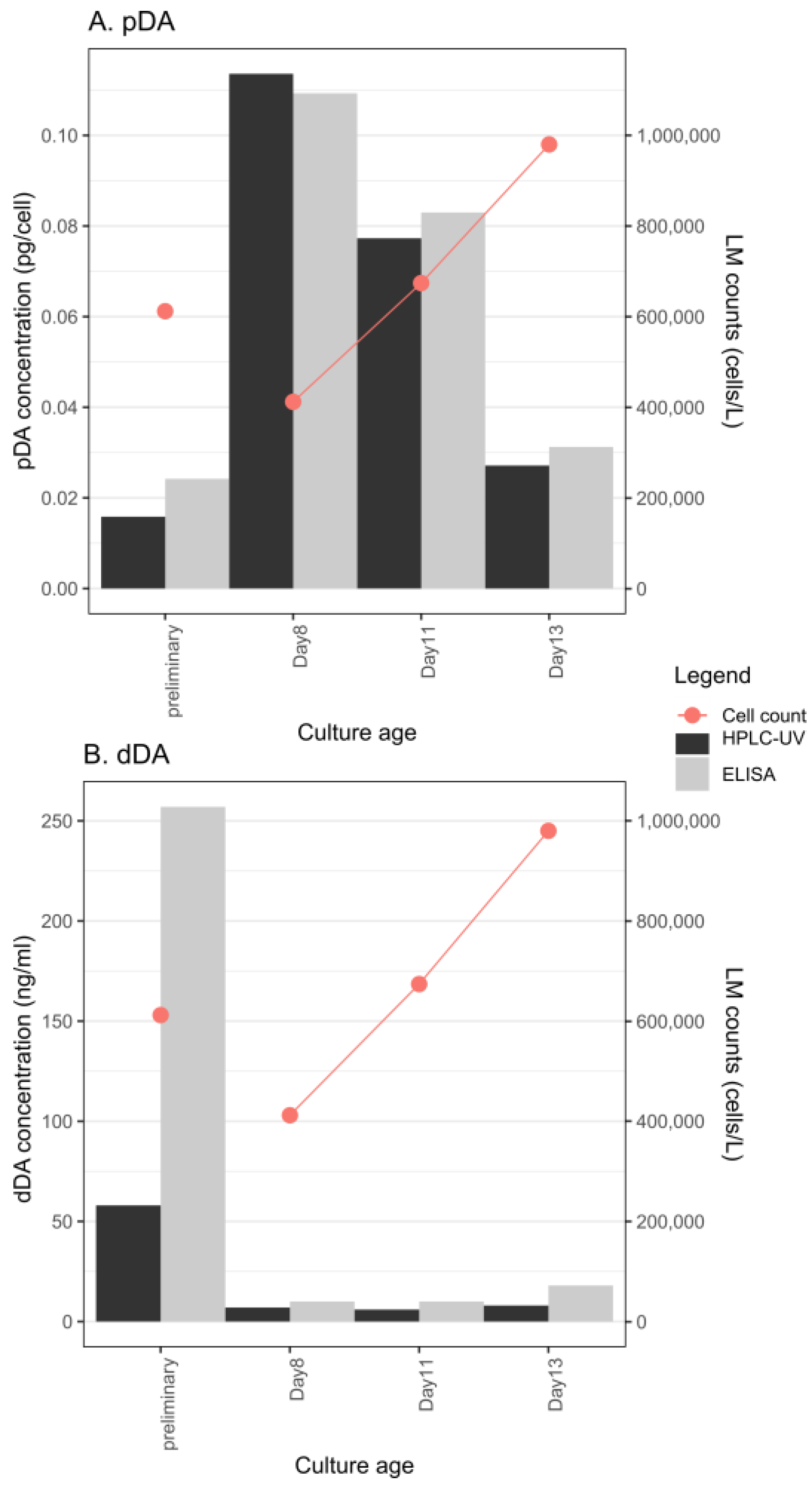
2.2. DA Production in Different Growth Phases
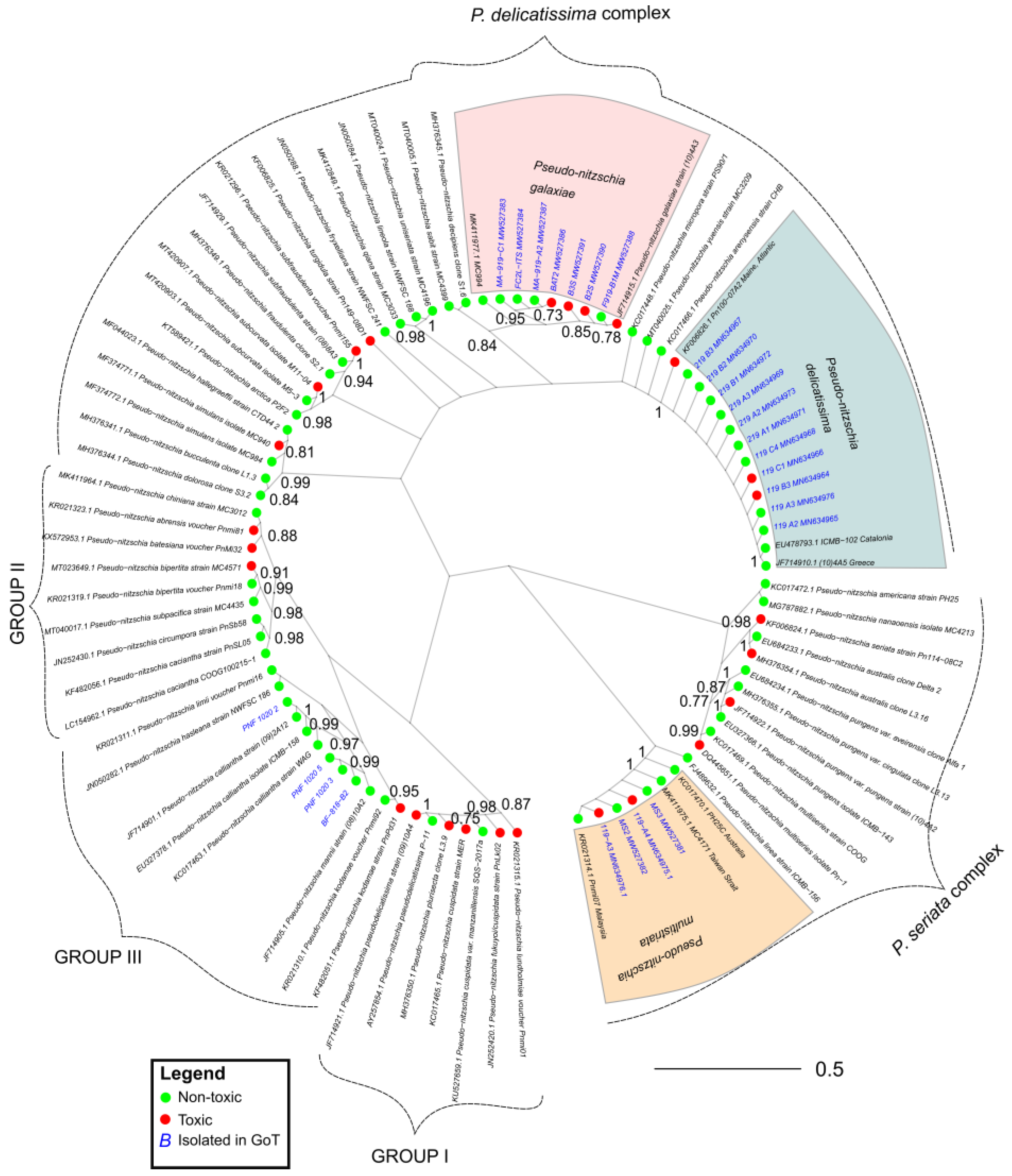
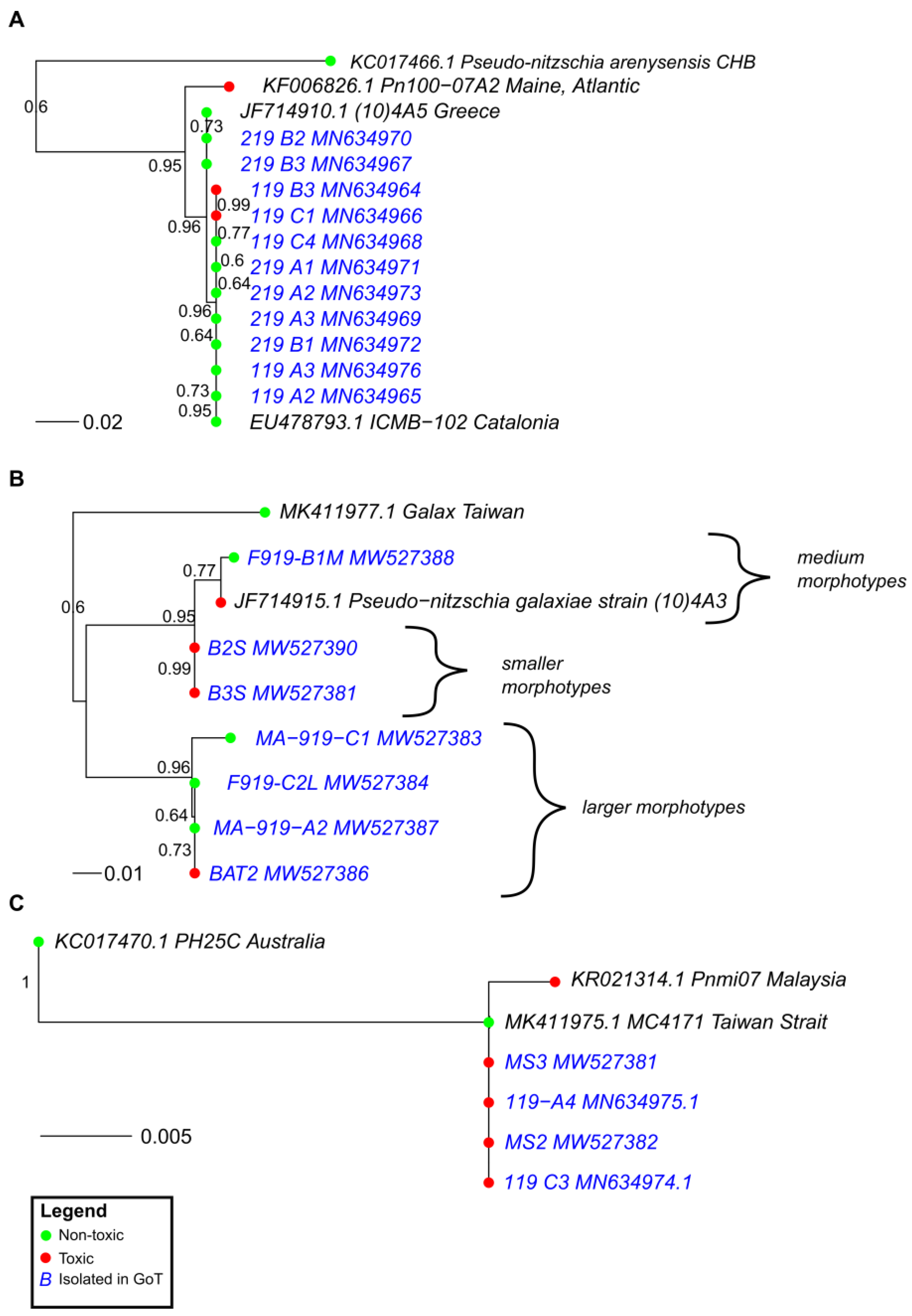
2.3. Phylogeny and Toxicity
2.4. dabA Gene Screening in Toxic and Non-Toxic Strains
3. Discussion
3.1. Toxicity
3.2. Phylogenetic Relationships between Toxic and Non-Toxic Strains
3.3. DabA in Selected Strains
4. Materials and Methods
4.1. Species Cultures
4.2. Domoic Acid in Cultures
4.3. Direct Competitive ELISA
4.4. HPLC-UV Method for Selected Cultures
4.5. LC-MS/MS for Selected Cultures
4.6. ITS-2 Phylogeny Reconstruction with Tested Strains
4.7. Amplification and Sequence Analysis of the dabA Gene
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bates, S.S.; Hubbard, K.A.; Lundholm, N.; Montresor, M.; Leaw, C.P. Pseudo-nitzschia, Nitzschia, and domoic acid: New research since 2011. Harmful Algae 2018, 1–41. [Google Scholar] [CrossRef]
- Huang, C.X.; Dong, H.C.; Lundholm, N.; Teng, S.T.; Zheng, G.C.; Tan, Z.J.; Lim, P.T.; Li, Y. Species composition and toxicity of the genus Pseudo-nitzschia in Taiwan Strait, including P. chiniana sp. nov. and P. qiana sp. nov. Harmful Algae 2019, 84, 195–209. [Google Scholar] [CrossRef]
- Lelong, A.; Hégaret, H.; Soudant, P.; Bates, S.S. Pseudo-nitzschia (Bacillariophyceae) species, domoic acid and amnesic shellfish poisoning: Revisiting previous paradigms. Phycologia 2012, 51, 168–216. [Google Scholar] [CrossRef] [Green Version]
- Rue, E.; Bruland, K. Domoic acid binds iron and copper: A possible role for the toxin produced by the marine diatom Pseudo-nitzschia. Mar. Chem. 2001, 76, 127–134. [Google Scholar] [CrossRef]
- Maldonado, M.T.; Hughes, M.P.; Rue, E.L.; Wells, M.L. The effect of Fe and Cu on growth and domoic acid production by Pseudo-nitzschia multiseries and Pseudo-nitzschia australis. Limnol. Oceanogr. 2002, 47, 515–526. [Google Scholar] [CrossRef]
- Wells, M.L.; Trick, C.G.; Cochlan, W.P.; Hughes, M.P.; Trainer, V.L. Domoic acid: The synergy of iron, copper, and the toxicity of diatoms. Limnol. Oceanogr. 2005, 50, 1908–1917. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gu, Y.; Lou, Y.; Wang, G. Response mechanisms of domoic acid in Pseudo-nitzschia multiseries under copper stress. Environ. Pollut. 2021, 272, 115578. [Google Scholar] [CrossRef] [PubMed]
- Geuer, J.K.; Trimborn, S.; Koch, F.; Brenneis, T.; Krock, B.; Koch, B.P. Dissolved Domoic Acid Does Not Improve Growth Rates and Iron Content in Iron-Stressed Pseudo-nitzschia subcurvata. Front. Mar. Sci. 2020, 7, 478. [Google Scholar] [CrossRef]
- Brunson, J.K.; McKinnie, S.M.K.; Chekan, J.R.; McCrow, J.P.; Miles, Z.D.; Bertrand, E.M.; Bielinski, V.A.; Luhavaya, H.; Oborník, M.; Smith, G.J.; et al. Biosynthesis of the neurotoxin domoic acid in a bloom-forming diatom. Science 2018, 361, 1356–1358. [Google Scholar] [CrossRef] [Green Version]
- Tammilehto, A.; Nielsen, T.G.; Krock, B.; Møller, E.F.; Lundholm, N. Induction of domoic acid production in the toxic diatom Pseudo-nitzschia seriata by calanoid copepods. Aquat. Toxicol. 2015, 159, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Haroardóttir, S.; Wohlrab, S.; Hjort, D.M.; Krock, B.; Nielsen, T.G.; John, U.; Lundholm, N. Transcriptomic responses to grazing reveal the metabolic pathway leading to the biosynthesis of domoic acid and highlight different defense strategies in diatoms. BMC Mol. Biol. 2019, 20, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zheng, T.; Lundholm, N.; Huang, X.; Jiang, X.; Li, A.; Li, Y. Chemical and morphological defenses of Pseudo-nitzschia multiseries in response to zooplankton grazing. Harmful Algae 2021, 104, 102033. [Google Scholar] [CrossRef]
- Turk Dermastia, T.; Cerino, F.; Stanković, D.; Francé, J.; Ramšak, A.; Žnidarič Tušek, M.; Beran, A.; Natali, V.; Cabrini, M.; Mozetič, P. Ecological time series and integrative taxonomy unveil seasonality and diversity of the toxic diatom Pseudo-nitzschia H. Peragallo in the northern Adriatic Sea. Harmful Algae 2020, 93, 101773. [Google Scholar] [CrossRef] [PubMed]
- Zingone, A.; Escalera, L.; Aligizaki, K.; Fernández-Tejedor, M.; Ismael, A.; Montresor, M.; Mozetič, P.; Taş, S.; Totti, C. Toxic marine microalgae and noxious blooms in the Mediterranean Sea: A contribution to the Global HAB Status Report. Harmful Algae 2021, 102, 101843. [Google Scholar] [CrossRef] [PubMed]
- UVHVVR. Yearly Report On Zoonosis and Their Causative Organisms; 2019. Available online: https://www.gov.si/assets/organi-v-sestavi/UVHVVR/Varna-hrana/Porocila-bioloska-varnost/Nacionalno-porocilo-monitoringa-zoonoz-2019.pdf (accessed on 19 December 2021). (In Slovenian)
- Arapov, J.; Skejić, S.; Bužančić, M.; Bakrač, A.; Vidjak, O.; Bojanić, N.; Ujević, I.; Gladan, Ž.N. Taxonomical diversity of Pseudo-nitzschia from the Central Adriatic Sea. Phycol. Res. 2017, 65, 280–290. [Google Scholar] [CrossRef]
- Penna, A.; Casabianca, S.; Perini, F.; Bastianini, M.; Riccardi, E.; Pigozzi, S.; Scardi, M. Toxic Pseudo-nitzschia spp. in the northwestern Adriatic Sea: Characterization of species composition by genetic and molecular quantitative analyses. J. Plankton Res. 2013, 35, 352–366. [Google Scholar] [CrossRef] [Green Version]
- Caroppo, C.; Congestri, R.; Bracchini, L.; Albertano, P. On the presence of Pseudo-nitzschia calliantha Lundholm, Moestrup et Hasle and Pseudo-nitzschia delicatissima (Cleve) Heiden in the Southern Adriatic Sea (Mediterranean Sea, Italy). J. Plankton Res. 2005, 27, 763–774. [Google Scholar] [CrossRef]
- Marić, D.; Ljubešić, Z.; Godrijan, J.; Viličić, D.; Ujević, I.; Precali, R. Blooms of the potentially toxic diatom Pseudo-nitzschia calliantha Lundholm, Moestrup & Hasle in coastal waters of the northern Adriatic Sea (Croatia). Estuar. Coast. Shelf Sci. 2011, 92, 323–331. [Google Scholar] [CrossRef]
- Giulietti, S.; Romagnoli, T.; Siracusa, M.; Bacchiocchi, S.; Totti, C.; Accoroni, S. Integrative taxonomy of the Pseudo-nitzschia (Bacillariophyceae) populations in the NW Adriatic Sea, with a focus on a novel cryptic species in the P. delicatissima species complex. Phycologia 2021, 60, 1–18. [Google Scholar] [CrossRef]
- Pistocchi, R.; Guerrini, F.; Pezzolesi, L.; Riccardi, M.; Vanucci, S.; Ciminiello, P.; Dell’Aversano, C.; Forino, M.; Fattorusso, E.; Tartaglione, L.; et al. Toxin levels and profiles in microalgae from the North-Western Adriatic Sea - 15 Years of studies on cultured species. Mar. Drugs 2012, 10, 140–162. [Google Scholar] [CrossRef] [Green Version]
- Arapov, J.; Ujević, I.; Straka, M.; Skejić, S.; Bužančić, M.; Bakrač, A.; Gladan, Ž.N. First evidence of domoic acid production in Pseudo-nitzschia calliantha cultures from the central Adriatic Sea. Acta Adriat. 2020, 61, 135–144. [Google Scholar] [CrossRef]
- Arapov, J.; Ujevic, I.; Pfannkuchen, D.M.; Godrijan, J.; Bakrac, A.; Gladan, Z.N.; Marasovic, I. Domoic acid in phytoplankton net samples and shellfish from the Krka river estuary in the central Adriatic Sea. Mediterr. Mar. Sci. 2016, 17, 340–350. [Google Scholar] [CrossRef]
- Cerino, F.; Orsini, L.; Sarno, D.; Dell’Aversano, C.; Tartaglione, L.; Zingone, A. The alternation of different morphotypes in the seasonal cycle of the toxic diatom Pseudo-nitzschia galaxiae. Harmful Algae 2005, 4, 33–48. [Google Scholar] [CrossRef]
- Lim, H.C.; Tan, S.N.; Teng, S.T.; Lundholm, N.; Orive, E.; David, H.; Quijano-Scheggia, S.; Leong, S.C.Y.; Wolf, M.; Bates, S.S.; et al. Phylogeny and species delineation in the marine diatom Pseudo-nitzschia (Bacillariophyta) using cox1, LSU, and ITS2 rRNA genes: A perspective in character evolution. J. Phycol. 2018, 54, 234–248. [Google Scholar] [CrossRef]
- Basu, S.; Patil, S.; Mapleson, D.; Russo, M.T.; Vitale, L.; Fevola, C.; Maumus, F.; Casotti, R.; Mock, T.; Caccamo, M.; et al. Finding a partner in the ocean: Molecular and evolutionary bases of the response to sexual cues in a planktonic diatom. New Phytol. 2017, 215, 140–156. [Google Scholar] [CrossRef] [Green Version]
- Trainer, V.L.; Bates, S.S.; Lundholm, N.; Thessen, A.E.; Cochlan, W.P.; Adams, N.G.; Trick, C.G. Pseudo-nitzschia physiological ecology, phylogeny, toxicity, monitoring and impacts on ecosystem health. Harmful Algae 2012, 14, 271–300. [Google Scholar] [CrossRef] [Green Version]
- HAEDAT Harmful Algae Event Database. Available online: http://haedat.iode.org/eventSearch.php?searchtext%5BcountryID%5D=58 (accessed on 8 June 2021).
- Tenorio, C.; Álvarez, G.; Quijano-Scheggia, S.; Perez-Alania, M.; Arakaki, N.; Araya, M.; Álvarez, F.; Blanco, J.; Uribe, E. First report of domoic acid production from Pseudo-nitzschia multistriata in Paracas bay (Peru). Toxins 2021, 13, 408. [Google Scholar] [CrossRef]
- Orsini, L.; Sarno, D.; Procaccini, G.; Poletti, R.; Dahlmann, J.; Montresor, M. Toxic Pseudo-nitzschia multistriata (Bacillariophyceae) from the Gulf of Naples: Morphology, toxin analysis and phylogenetic relationships with other Pseudo-nitzschia species. Eur. J. Phycol. 2002, 37, 247–257. [Google Scholar] [CrossRef]
- Amato, A.; Lüdeking, A.; Kooistra, W.H.C.F. Intracellular domoic acid production in Pseudo-nitzschia multistriata isolated from the Gulf of Naples (Tyrrhenian Sea, Italy). Toxicon 2010, 55, 157–161. [Google Scholar] [CrossRef]
- Ruggiero, M.V.; Sarno, D.; Barra, L.; Kooistra, W.H.C.F.; Montresor, M.; Zingone, A. Diversity and temporal pattern of Pseudo-nitzschia species (Bacillariophyceae) through the molecular lens. Harmful Algae 2015, 42, 15–24. [Google Scholar] [CrossRef]
- Moschandreou, K.K.; Baxevanis, A.D.; Katikou, P.; Papaefthimiou, D.; Nikolaidis, G.; Abatzopoulos, T.J. Inter- and intra-specific diversity of Pseudo-nitzschia (Bacillariophyceae) in the northeastern Mediterranean. Eur. J. Phycol. 2012, 47, 321–339. [Google Scholar] [CrossRef]
- Amato, A.; Montresor, M. Morphology, phylogeny, and sexual cycle of Pseudo-nitzschia mannii sp. nov. (Bacillariophyceae): A pseudo-cryptic species within the P. pseudodelicatissima complex. Phycologia 2008, 47, 487–497. [Google Scholar] [CrossRef]
- Pan, Y.; Parsons, M.L.; Busman, M.; Moeller, P.D.R.; Dortch, Q.; Powell, C.L.; Doucette, G.J. Pseudo-nitzschia sp. cf. pseudodelicatissima-A confirmed producer of domoic acid from the northern Gulf of Mexico. Mar. Ecol. Prog. Ser. 2001, 220, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Lundholm, N.; Hansen, P.J.; Kotaki, Y. Effect of pH on growth and domoic acid production by potentially toxic diatoms of the genera Pseudo-nitzschia and Nitzschia. Mar. Ecol. Prog. Ser. 2004, 1–15. [Google Scholar] [CrossRef]
- Amin, S.A.; Parker, M.S.; Armbrust, E.V. Interactions between Diatoms and Bacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 667–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Maucher-Fuquay, J.; Fire, S.E.; Mikulski, C.M.; Haynes, B.; Doucette, G.J.; Ramsdell, J.S. Optimization of solid-phase extraction and liquid chromatography-tandem mass spectrometry for the determination of domoic acid in seawater, phytoplankton, and mammalian fluids and tissues. Anal. Chim. Acta 2012, 715, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.A.; Papapanagiotou, E.P.; Brown, N.A.; Stobo, L.A.; Gallacher, S.; Shanks, A.M. Effect of storage on amnesic shellfish poisoning (ASP) toxins in king scallops (Pecten maximus). Harmful Algae 2006, 5, 9–19. [Google Scholar] [CrossRef]
- Howard, M.D.A.; Smith, G.J.; Kudela, R.M. Phylogenetic relationships of yessotoxin-producing dinoflagellates, based on the large subunit and internal transcribed spacer ribosomal DNA domains. Appl. Environ. Microbiol. 2009, 75, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Lema, K.A.; Metegnier, G.; Quéré, J.; Latimier, M.; Youenou, A.; Lambert, C.; Fauchot, J.; Le Gac, M.; Costantini, M. Inter- and Intra-Specific Transcriptional and Phenotypic Responses of Pseudo-nitzschia under Different Nutrient Conditions. Genome Biol. Evol. 2019, 11, 731–747. [Google Scholar] [CrossRef]
- Pulido, O.M. Domoic acid toxicologic pathology: A review. Mar. Drugs 2008, 6, 180–219. [Google Scholar] [CrossRef]
- Tan, S.N.; Teng, S.T.; Lim, H.C.; Kotaki, Y.; Bates, S.S.; Leaw, C.P.; Lim, P.T. Diatom Nitzschia navis-varingica (Bacillariophyceae) and its domoic acid production from the mangrove environments of Malaysia. Harmful Algae 2016, 60, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Smida, D.B.; Lundholm, N.; Kooistra, W.H.C.F.; Sahraoui, I.; Ruggiero, M.V.; Kotaki, Y.; Ellegaard, M.; Lambert, C.; Mabrouk, H.H.; Hlaili, A.S. Morphology and molecular phylogeny of Nitzschia bizertensis sp. nov.-A new domoic acid-producer. Harmful Algae 2014, 32, 49–63. [Google Scholar] [CrossRef]
- Chekan, J.R.; McKinnie, S.M.K.; Noel, J.P.; Moore, B.S. Algal neurotoxin biosynthesis repurposes the terpene cyclase structural fold into an N-prenyltransferase. Proc. Natl. Acad. Sci. USA 2020, 117, 12799–12805. [Google Scholar] [CrossRef]
- Orr, R.J.S.; Stüken, A.; Murray, S.A.; Jakobsen, K.S. Evolution and distribution of saxitoxin biosynthesis in dinoflagellates. Mar. Drugs 2013, 11, 2814–2828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuhrman, J.A. Marine viruses and their biogeochemical and ecological effects. Nature 1999, 399, 541–548. [Google Scholar] [CrossRef]
- Casteleyn, G.; Adams, N.G.; Vanormelingen, P.; Debeer, A.-E.; Sabbe, K.; Vyverman, W. Natural Hybrids in the Marine Diatom Pseudo-nitzschia pungens (Bacillariophyceae): Genetic and Morphological Evidence. Protist 2009, 160, 343–354. [Google Scholar] [CrossRef]
- Tesson, S.V.M.; Legrand, C.; van Oosterhout, C.; Montresor, M.; Kooistra, W.H.C.F.; Procaccini, G. Mendelian Inheritance Pattern and High Mutation Rates of Microsatellite Alleles in the Diatom Pseudo-nitzschia multistriata. Protist 2013, 164, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Schliep, K.P. Phangorn: Phylogenetic analysis in R. Bioinformatics 2011, 27, 592–593. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Sampling Frequency 1 | Partition Tested | Test Method 2 | Result 3 | DA Concentration4 | ELISA Retest 5 |
|---|---|---|---|---|---|---|---|
| P. delicatissima | 219 A1 | P | pDA | E | - | ||
| 219 A2 | P | pDA | E | - | |||
| 219 A3 | P | pDA | E | - | |||
| 219 B1 | P | pDA | E | - | |||
| 219 B2 | P | pDA | E | - | |||
| 219 B3 | P | pDA | E | - | |||
| 219 B4 | P | pDA | E | - | |||
| 119 A2 | P | pDA | E | - | |||
| 119 A3 | P | pDA | E | - | |||
| 119 B3 | P & C | pDA and dDA | E&H | + (E) | 1.5 fg/cell | <quant | |
| 119 C1 | P | pDA and dDA | E&H | + (E) | NA | <quant | |
| 119 C4 | P | pDA | E | - | |||
| P. multistriata | 119 A4 | P & C | pDA and dDA | E&H | + (E&H) | pDA: 16–114 fg/cell; dDA: 6–257 ng/mL | |
| 119 C3 | P | pDA and dDA | E&H | + (E&H) | pDA: 121–160 fg/cell; dDA: 30–80 | ||
| MS2 | P & C | pDA and dDA | E; H; L | + (E) | pDA: 0–32.8 fg/cell; dDA: 20 ng/mL | <quant | |
| MS3 | P & C | pDA and dDA | E; H; L | + (E) | pDA: 1.42–20.7 fg/cell; dDA: 14.6–38.5 ng/mL | <quant | |
| PN0DB2131216-A | P | pDA | L | - | |||
| PN0DB2131216-B | P | pDA | L | - | |||
| PN0DB2131216-C | P | pDA | L | + | 0.217 fg/cell | ||
| P. galaxiae—large | MA-919-C1 | P | pDA and dDA | E&H | - | ||
| MA-919-A2 | P | pDA and dDA | E&H | - | |||
| F919-C2L | P | pDA and dDA | E&H | - | |||
| P. galaxiae— medium | F919-B1M | P | pDA and dDA | E&H | - | ||
| P. galaxiae—bat-like | BAT2 | P | pDA and dDA | E&H | o (E) | dDA: 13 ng/mL | <quant |
| P. galaxiae—small | B3S | P | pDA and dDA | E&H | + (E) | dDA: 5–24.8 ng/mL; pDA > quant | >quant |
| B2S | P | pDA and dDA | E&H | o (E) | dDA: 12.6 ng/mL | <quant | |
| P. mannii | BF-819-B2 | P | pDA and dDA | E&H | - | ||
| BF-819-A4 | P | pDA and dDA | E&H | - | |||
| BD-919-A3 | P | pDA and dDA | E&H | - | |||
| PNF_1020_5 | P | pDA and dDA | E | o | <quant | ||
| P. subfraudulenta | PNF_1020_1 | P | pDA and dDA | E | - | ||
| P. calliantha | PNF_1020_2 | P | pDA and dDA | E | o | <quant | |
| PN00BF281016-2A | P | pDA | L | - |
| Primer Name | Primer-Sequence | Type |
|---|---|---|
| DabA_multF | ATGAAATTTGCAACGTCCATTGTC | PCR |
| DabA_degF | ATGAARTTTGCAACRTCCATYGTC | PCR |
| N1_R | TCCAAAAACGCTTTCATCAA | PCR |
| N2_R | AACGCTTTCATCAATGGTTTGTGG | PCR |
| Internal_multistriataF | CGATTGGATGAAGATCCCTTCA | Sequencing |
| Internal_multistriataR | GCAGAAGTCGACCATCCA | Sequencing |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turk Dermastia, T.; Dall’Ara, S.; Dolenc, J.; Mozetič, P. Toxicity of the Diatom Genus Pseudo-nitzschia (Bacillariophyceae): Insights from Toxicity Tests and Genetic Screening in the Northern Adriatic Sea. Toxins 2022, 14, 60. https://doi.org/10.3390/toxins14010060
Turk Dermastia T, Dall’Ara S, Dolenc J, Mozetič P. Toxicity of the Diatom Genus Pseudo-nitzschia (Bacillariophyceae): Insights from Toxicity Tests and Genetic Screening in the Northern Adriatic Sea. Toxins. 2022; 14(1):60. https://doi.org/10.3390/toxins14010060
Chicago/Turabian StyleTurk Dermastia, Timotej, Sonia Dall’Ara, Jožica Dolenc, and Patricija Mozetič. 2022. "Toxicity of the Diatom Genus Pseudo-nitzschia (Bacillariophyceae): Insights from Toxicity Tests and Genetic Screening in the Northern Adriatic Sea" Toxins 14, no. 1: 60. https://doi.org/10.3390/toxins14010060
APA StyleTurk Dermastia, T., Dall’Ara, S., Dolenc, J., & Mozetič, P. (2022). Toxicity of the Diatom Genus Pseudo-nitzschia (Bacillariophyceae): Insights from Toxicity Tests and Genetic Screening in the Northern Adriatic Sea. Toxins, 14(1), 60. https://doi.org/10.3390/toxins14010060