Exposure to Low Zearalenone Doses and Changes in the Homeostasis and Concentrations of Endogenous Hormones in Selected Steroid-Sensitive Tissues in Pre-Pubertal Gilts



Abstract
:1. Introduction
2. Results
2.1. Experimental Feed
2.2. Results of Laboratory Analyses
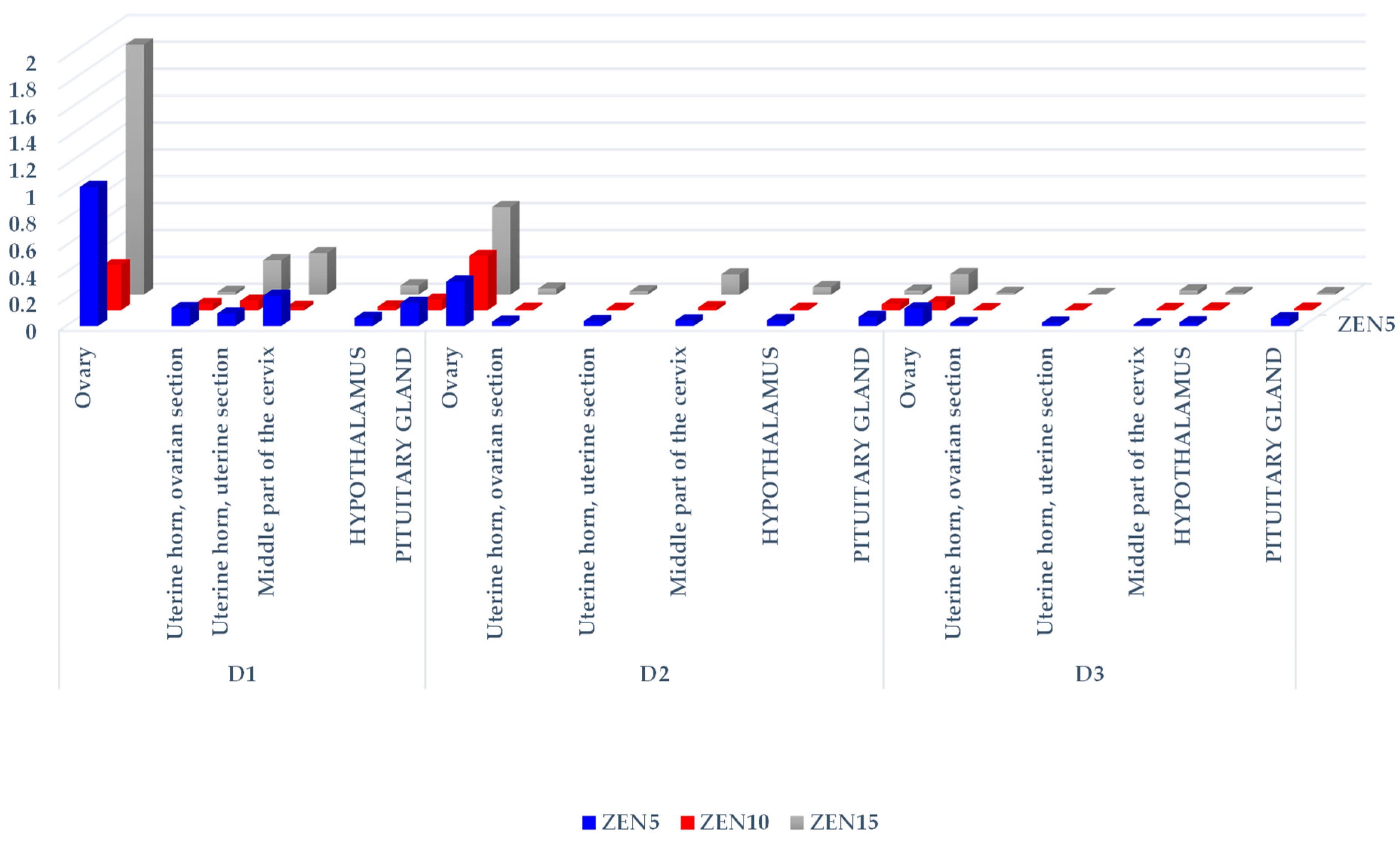
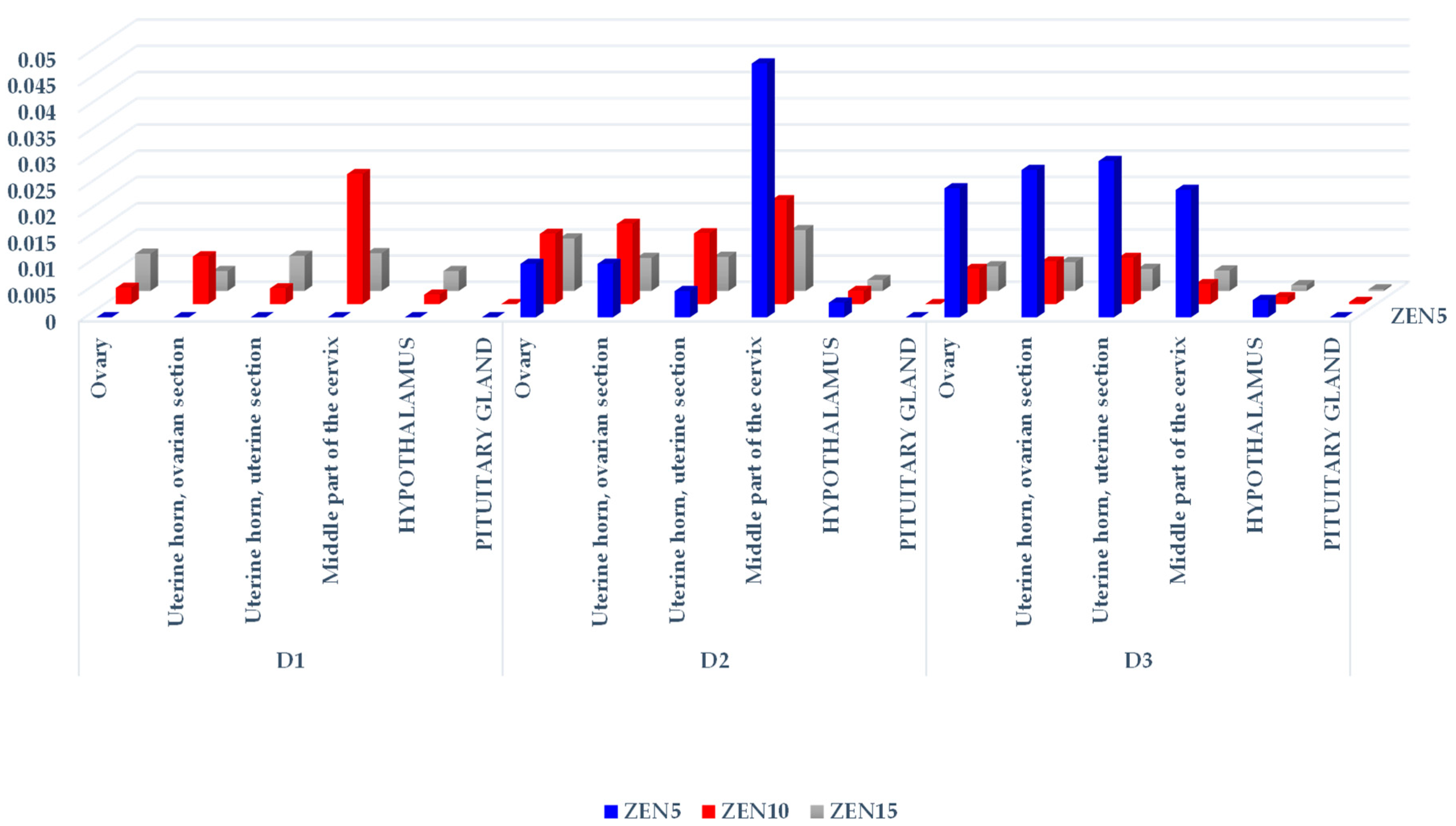
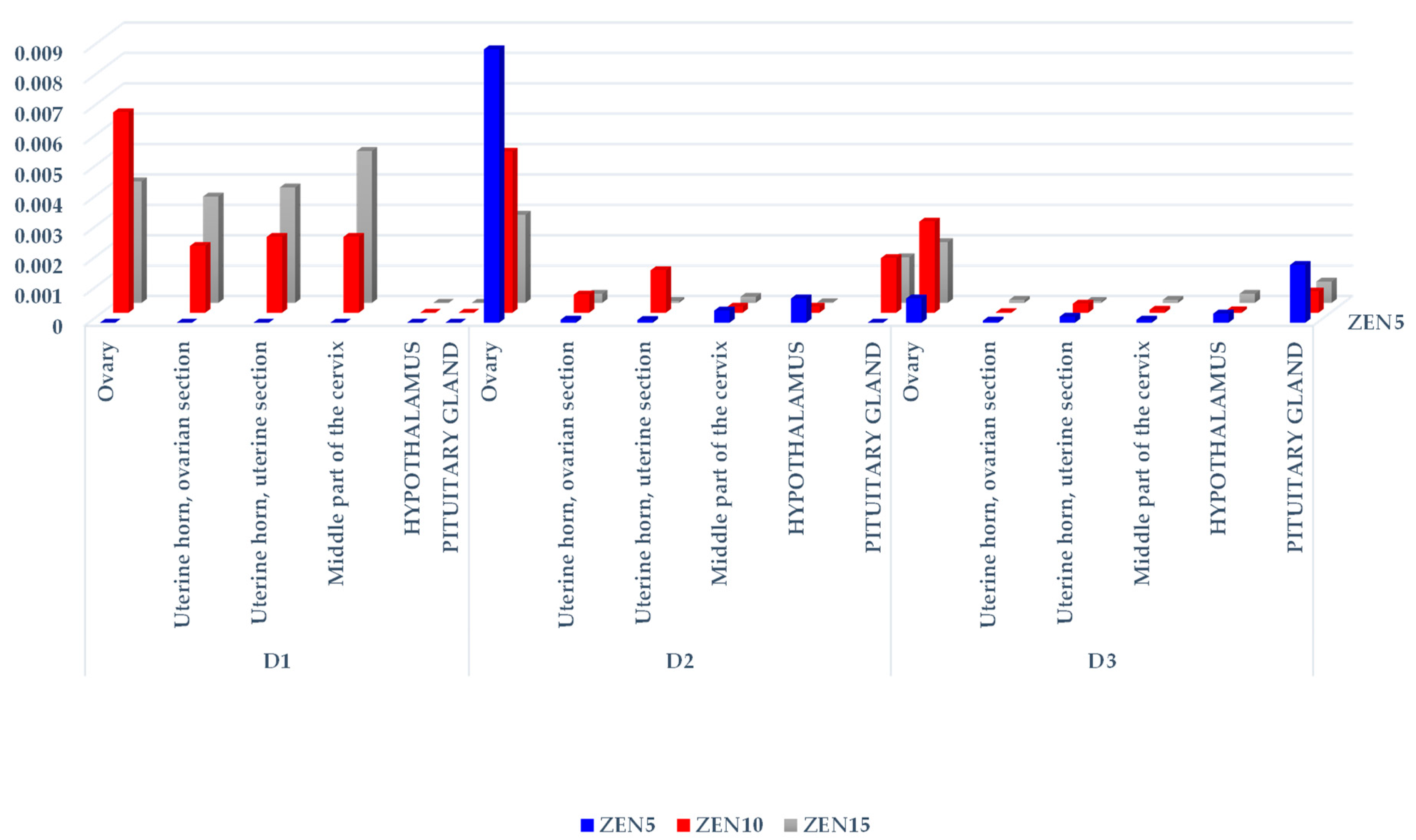
2.2.1. Concentrations of ZEN and Its Metabolites in Selected Tissues
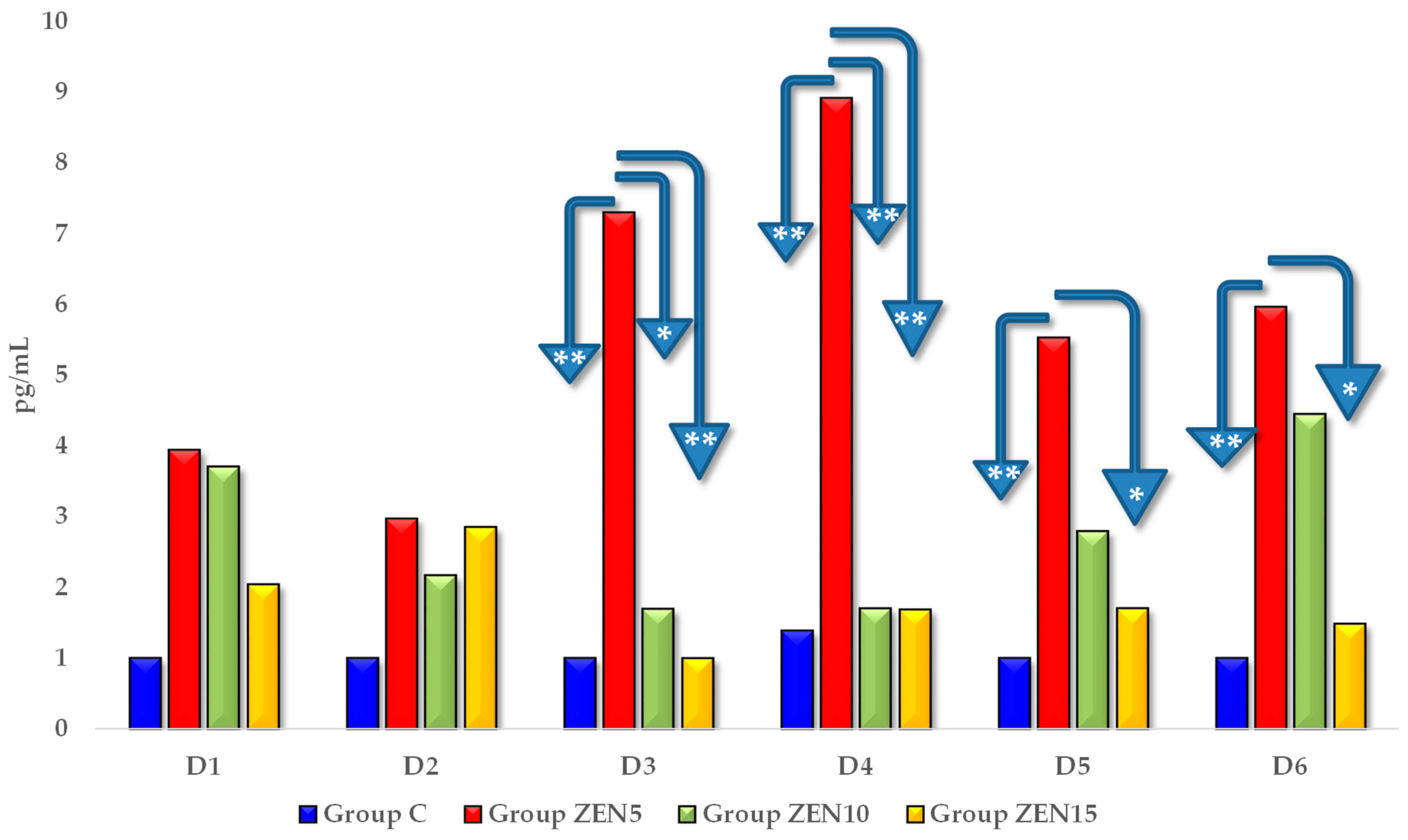
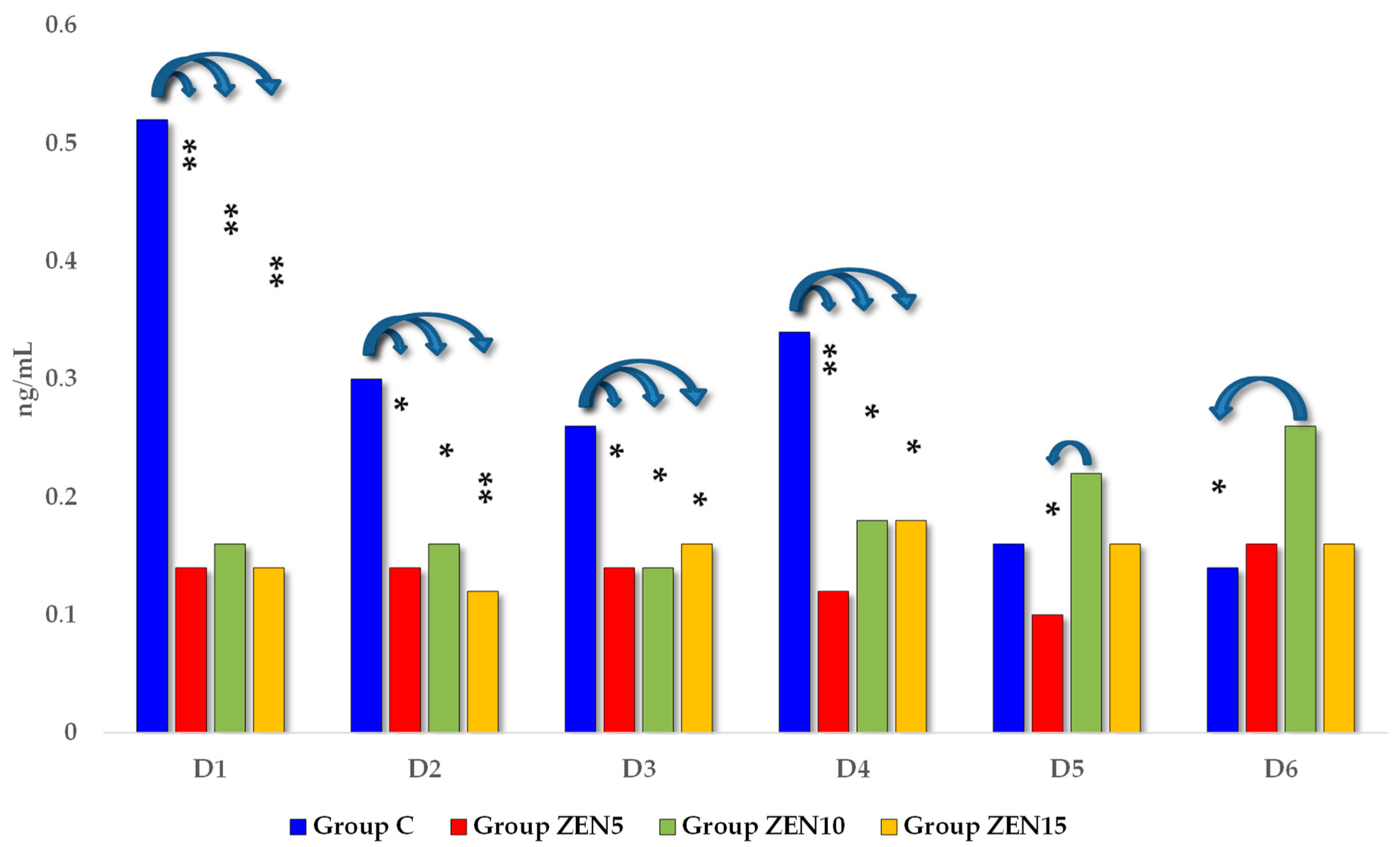
2.2.2. Blood Concentrations of estradiol and progesterone
3. Discussion
4. Summary
5. Materials and Methods
5.1. General Information
5.2. Experimental Feed
Toxicological Analysis of Feed
5.3. Experimental Animals
5.4. Toxicological Studies of Reproductive, Hypothalamic, and Pituitary Gland Tissues
5.4.1. Tissue Samples
5.4.2. Extraction Procedure
5.4.3. Chromatographic Quantification of Zearalenone and Its Metabolites
5.4.4. Mass Spectrometry Conditions
5.4.5. CF
5.4.6. Statistical Analysis
5.5. Toxicological Studies of Blood
Blood samples collection
5.6. Determination of Plasma Hormone Concentrations
5.6.1. Estradiol
5.6.2. Progesterone
5.6.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thielecke, F.; Nugent, A.P. Contaminants in Grain—A Major Risk for Whole Grain Safety? Nutrients 2018, 10, 1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleetwood, J.; Rahman, S.; Holland, D.; Millson, D.; Thomson, L.; Poppy, G. As clean as they look? Food hygiene inspection scores, microbiological contamination, and foodborne illness. Food Control 2019, 96, 76–86. [Google Scholar] [CrossRef]
- Alassane-Kpembi, I.; Pinton, P.; Oswald, I.P. Effects of Mycotoxins on the Intestine. Toxins 2019, 11, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piotrowska, M.; Sliżewska, K.; Nowak, A.; Zielonka, Ł.; Żakowska, Z.; Gajęcka, M.; Gajęcki, M. The effect of experimental fusarium mycotoxicosis on microbiota diversity in porcine ascending colon contents. Toxins 2014, 6, 2064–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zachariasova, M.; Dzumana, Z.; Veprikova, Z.; Hajkovaa, K.; Jiru, M.; Vaclavikova, M.; Zachariasova, A.; Pospichalova, M.; Florian, M.; Hajslova, J. Occurrence of multiple mycotoxins in European feeding stuffs, assessment of dietary intake by farm animals. Anim. Feed Sci. Technol. 2014, 193, 124–140. [Google Scholar] [CrossRef]
- Knutsen, H.-K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; et al. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, 4851. [Google Scholar] [CrossRef] [Green Version]
- Faisal, Z.; Vörös, V.; Fliszár-Nyúl, E.; Lemli, B.; Kunsági-Máté, S.; Poór, M. Interactions of zearalanone, α-zearalanol, β-zearalanol, zearalenone-14-sulfate, and zearalenone-14-glucoside with serum albumin. Mycotoxin Res. 2020, 36, 389–397. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormesis: Path and Progression to Significance. Int. J. Mol. Sci. 2018, 19, 2871. [Google Scholar] [CrossRef] [Green Version]
- Freire, L.; Sant’Ana, A.S. Modified mycotoxins: An updated review on their formation, detection, occurrence, and toxic effects. Food Chem. Toxicol. 2018, 111, 189–205. [Google Scholar] [CrossRef]
- Rykaczewska, A.; Gajęcka, M.; Dąbrowski, M.; Wiśniewska, A.; Szcześniewska, J.; Gajęcki, M.T.; Zielonka, Ł. Growth performance, selected blood biochemical parameters and body weight of pre-pubertal gilts fed diets supplemented with different doses of zearalenone (ZEN). Toxicon 2018, 152, 84–94. [Google Scholar] [CrossRef]
- Cieplińska, K.; Gajęcka, M.; Dąbrowski, M.; Rykaczewska, A.; Zielonka, Ł.; Lisieska-Żołnierczyk, S.; Bulińska, M.; Gajęcki, M.T. Time-dependent changes in the intestinal microbiome of gilts exposed to low zearalenone doses. Toxins 2019, 11, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieplińska, K.; Gajęcka, M.; Nowak, A.; Dąbrowski, M.; Zielonka, Ł.; Gajęcki, M.T. The gentoxicity of caecal water in gilts exposed to low doses of zearalenone. Toxins 2018, 10, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajęcka, M.; Majewski, M.S.; Zielonka, Ł.; Grzegorzewski, W.; Onyszek, E.; Lisieska-Zołnierczyk, S.; Juśkiewicz, J.; Babuchowski, A.; Gajęcki, M.T. Concentration of Zearalenone, Alpha-Zearalenol and Beta-Zearalenol in the Myocardium and the Results of Isometric Analyses of the Coronary Artery in Prepubertal Gilts. Toxins 2021, 13, 396. [Google Scholar] [CrossRef]
- Mróz, M.; Gajęcka, M.; Przybyłowicz, K.E.; Sawicki, T.; Lisieska-Żołnierczyk, S.; Zielonk, Ł.; Gajęcki, M.T. The Effect of Low Doses of Zearalenone (ZEN) on the Bone Marrow Microenvironment and Haematological Parameters of Blood Plasma in Pre-Pubertal Gilts. Toxins 2022, 14, 105. [Google Scholar] [CrossRef] [PubMed]
- Mróz, M.; Gajęcka, M.; Brzuzan, P.; Lisieska-Żołnierczyk, S.; Leski, D.; Zielonka, Ł.; Gajęcki, M.T. Carry-Over of Zearalenone and Its Metabolites to Intestinal Tissues and the Expression of CYP1A1 and GSTπ1 in the Colon of Gilts before Puberty. Toxins 2022, 14, 354. [Google Scholar] [CrossRef]
- Zhou, J.; Zhao, L.; Huang, S.; Liu, Q.; Ao, X.; Lei, Y.; Ji, C.; Ma, Q. Zearalenone toxicosis on reproduction as estrogen receptor selective modulator and alleviation of zearalenone biodegradative agent in pregnant sows. J. Anim. Sci. Biotechnol. 2022, 13, 36. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. Activity of zearalenone in the porcine intestinal tract. Molecules 2017, 22, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celi, P.; Verlhac, V.; Pérez, C.E.; Schmeisser, J.; Kluenter, A.M. Biomarkers of gastrointestinal functionality in animal nutrition and health. Anim. Feed Sci. Technol. 2019, 250, 9–31. [Google Scholar] [CrossRef]
- Dąbrowski, M.; Obremski, K.; Gajęcka, M.; Gajęcki, M.; Zielonka, Ł. Changes in the subpopulations of porcine peripheral blood lymphocytes induced by exposure to low doses of zearalenone (ZEN) and deoxynivalenol (DON). Molecules 2016, 21, 557. [Google Scholar] [CrossRef] [Green Version]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Llorens, P.; Herrera, M.; Juan-García, A.; Payá, J.J.; Moltó, J.C.; Ariño, A.; Juan, C. Biomarkers of Exposure to Zearalenone in In Vivo and In Vitro Studies. Toxins 2022, 14, 291. [Google Scholar] [CrossRef] [PubMed]
- Szeliga, A.; Meczekalski, B. Kisspeptin Modulation of Reproductive Function. Endocrines 2022, 3, 367–374. [Google Scholar] [CrossRef]
- Wan, B.; Yuan, X.; Yang, W.; Jiao, N.; Li, Y.; Liu, F.; Liu, M.; Yang, Z.; Huang, L.; Jiang, S. The Effects of Zearalenone on the Localization and Expression of Reproductive Hormones in the Ovaries of Weaned Gilts. Toxins 2021, 13, 626. [Google Scholar] [CrossRef]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lathe, R.; Kotelevtsev, Y. Steroid signaling: Ligand-binding promiscuity molecular symmetry, and the need for gating. Steroids 2014, 82, 14–22. [Google Scholar] [CrossRef]
- Kramer, H.J.; van den Ham, W.A.; Slob, W.; Pieters, M.N. Conversion Factors Estimating Indicative Chronic No-Observed-Adverse-Effect Levels from Short-Term Toxicity Data. Regul. Toxicol. Pharm. 1996, 23, 249–255. [Google Scholar] [CrossRef]
- Pastoor, T.P.; Bachman, A.N.; Bell, D.R.; Cohen, S.M.; Dellarco, M.; Dewhurst, I.C.; Doe, J.E.; Doerrer, N.G.; Embry, M.R.; Hines, R.N.; et al. A 21st century roadmap for human health risk assessment. Crit. Rev. Toxicol. 2014, 44, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Suh, H.Y.; Peck, C.C.; Yu, K.S.; Lee, H. Determination of the starting dose in the first-in-human clinical trials with monoclonal antibodies: A systematic review of papers published between 1990 and 2013. Drug Des. Dev. Ther. 2016, 10, 4005–4016. [Google Scholar] [CrossRef] [Green Version]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.; Lee, D.-H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef]
- Gonkowski, S.; Gajęcka, M.; Makowska, K. Mycotoxins and the Enteric Nervous system. Toxins 2020, 12, 461. [Google Scholar] [CrossRef]
- Lephart, E.D.; Thompson, J.M.; Setchell, K.D.; Adlercreutz, H.; Weber, K.S. Phytoestrogens decrease brain calcium-binding proteins but do not alter hypothalamic androgen metabolizing enzymes in adult male rats. Brain Res. 2000, 859, 123–131. [Google Scholar] [CrossRef]
- Venkataramana, M.; Chandra Nayaka, S.; Anand, T.; Rajesh, R.; Aiyaz, M.; Divakara, S.T.; Murali, H.S.; Prakash, H.S.; Lakshmana Rao, P.V. Zearalenone induced toxicity in SHSY-5Y cells: The role of oxidative stress evidenced by N-acetyl cysteine. Food Chem. Toxicol. 2014, 65, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Guo, Z.; Xiang, W.; Wang, P. The neuroendocrine pathways and mechanisms for the control of the reproduction in female pigs. Anim. Reprod. 2021, 18, e20210063. [Google Scholar] [CrossRef] [PubMed]
- Genchi, V.A.; Rossi, E.; Lauriola, C.; D’Oria, R.; Palma, G.; Borrelli, A.; Caccioppoli, C.; Giorgino, F.; Cignarelli, A. Adipose Tissue Dysfunction and Obesity-Related Male Hypogonadism. Int. J. Mol. Sci. 2022, 23, 8194. [Google Scholar] [CrossRef] [PubMed]
- Marín-García, P.J.; Llobat, L. How does protein nutrition affect the epigenetic changes in pig? A review. Animals 2021, 11, 544. [Google Scholar] [CrossRef]
- Muro, B.B.D.; Leal, D.F.; Carnevale, R.F.; Torres, M.A.; Mendonça, M.V.; Nakasone, D.H.; Martinez, C.H.G.; Ravagnani, G.M.; Monteiro, M.S.; Poor, A.P.; et al. Altrenogest during early pregnancy modulates uterine glandular epithelium and endometrial growth factor expression at the time implantation in pigs. Anim. Reprod. 2021, 18, e20200431. [Google Scholar] [CrossRef]
- Rykaczewska, A.; Gajęcka, M.; Onyszek, E.; Cieplińska, K.; Dąbrowski, M.; Lisieska-Żołnierczyk, S.; Bulińska, M.; Babuchowski, A.; Gajęcki, M.T.; Zielonka, Ł. Imbalance in the Blood Concentrations of Selected Steroids in Prepubertal Gilts Depending on the Time of Exposure to Low Doses of Zearalenone. Toxins 2019, 11, 561. [Google Scholar] [CrossRef] [Green Version]
- Lawrenz, B.; Melado, L.; Fatemi, H. Premature progesterone rise in ART-cycles. Reprod. Biol. 2018, 18, 1–4. [Google Scholar] [CrossRef]
- Li, X.; Lin, B.; Zhang, X.; Shen, X.; Ouyang, H.; Wu, Z.; Tian, Y.; Fang, L.; Huang, Y. Comparative transcriptomics in the hypothalamic-pituitary-gonad axis of mammals and poultry. Genomics 2022, 114, 110396. [Google Scholar] [CrossRef]
- Zheng, W.; Feng, N.; Wang, Y.; Noll, L.; Xu, S.; Liu, X.; Lu, N.; Zou, H.; Gu, J.; Yuan, Y.; et al. Effects of zearalenone and its derivatives on the synthesis and secretion of mammalian sex steroid hormones: A review. Food Chem. Toxicol. 2019, 126, 262–276. [Google Scholar] [CrossRef]
- Yang, D.; Jiang, T.; Lin, P.; Chen, H.; Wang, L.; Wang, N.; Zhao, F.; Tang, K.; Zhou, D.; Wang, A.; et al. Apoptosis inducing factor gene depletion inhibits zearalenone-induced cell death in a goat Leydig cell line. Reprod. Toxicol. 2017, 67, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Piastowska-Ciesielska, A. Zearalenone as an endocrine disruptor in humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Benagiano, M.; Bianchi, P.; D’Elios, M.M.; Brosens, I.; Benagiano, G. Autoimmune diseases: Role of steroid hormones. Best Pract. Res. Clin. Obstet. Gynaecol. 2019, 60, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Kiss, D.S.; Ioja, E.; Toth, I.; Barany, Z.; Jocsak, G.; Bartha, T.; Horvath, T.L.; Zsarnovszky, A. Comparative Analysis of Zearalenone Effects on Thyroid Receptor Alpha (TRα) and Beta (TRβ) Expression in Rat Primary Cerebellar Cell Cultures. Int. J. Mol. Sci. 2018, 19, 1440. [Google Scholar] [CrossRef] [Green Version]
- Mucci, A.; Clemente, E. The Role of Genetics in Central Precocious Puberty: Confirmed and Potential Neuroendocrine Genetic and Epigenetic Contributors and Their Interactions with Endocrine Disrupting Chemicals (EDCs). Endocrines 2022, 3, 433–451. [Google Scholar] [CrossRef]
- Gajęcka, M.; Otrocka-Domagała, I. Immunocytochemical expression of 3β- and 17β-hydroxysteroid dehydrogenase in bitch ovaries exposed to low doses of zearalenone. Pol. J. Vet. Sci. 2013, 16, 55–62. [Google Scholar] [CrossRef]
- Liu, X.L.; Wu, R.Y.; Sun, X.F.; Cheng, S.F.; Zhang, R.Q.; Zhang, T.T.; Zhang, X.F.; Zhao, Y.; Shen, W.; Li, L. Mycotoxin zearalenone exposure impairs genomic stability of swine follicular granulosa cells in vitro. Int. J. Biol. Sci. 2018, 14, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Hennig-Pauka, I.; Koch, F.J.; Schaumberger, S.; Woechtl, B.; Novak, J.; Sulyok, M.; Nagl, V. Current challenges in the diagnosis of zearalenone toxicosis as illustrated by a field case of hyperestrogenism in suckling piglets. Porc. Health Manag. 2018, 4, 1–9. [Google Scholar] [CrossRef]
- He, J.; Wei, C.; Li, Y.; Liu, Y.; Wang, Y.; Pan, J.; Liu, J.; Wu, Y.; Cui, S. Zearalenone and alpha-zearalenol inhibit the synthesis and secretion of pig follicle stimulating hormone via the non-classical estrogen membrane receptor GPR30. Mol. Cell. Endocrinol. 2018, 461, 43–54. [Google Scholar] [CrossRef]
- Gellerich, F.N.; Gizatullina, Z.; Trumbeckaite, S.; Nguyen, H.P.; Pallas, T.; Arandarcikaite, O.; Vielhaber, S.; Seppet, E.; Striggow, F. The regulation of OXPHOS by extramitochondrial calcium. Biochim. Biophys. Acta (BBA)-Bioenerg. 2010, 1797, 1018–1027. [Google Scholar] [CrossRef]
- Gajęcka, M.; Przybylska-Gornowicz, B. The low doses effect of experimental zearalenone (ZEN) intoxication on the presence of Ca2+ in selected ovarian cells from pre-pubertal bitches. Pol. J. Vet. Sci. 2012, 15, 711–720. [Google Scholar] [CrossRef] [Green Version]
- Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. The Effect of Low Monotonic Doses of Zearalenone on Selected Reproductive Tissues in Pre-Pubertal Female Dogs—A Review. Molecules 2015, 20, 20669–20687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romejko, K.; Rymarz, A.; Sadownik, H.; Niemczyk, S. Testosterone Deficiency as One of the Major Endocrine Disorders in Chronic Kidney Disease. Nutrients 2022, 14, 3438. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Mróz, M.; Brzuzan, P.; Onyszek, E.; Zielonka, Ł.; Lipczyńska-Ilczuk, K.; Przybyłowicz, K.E.; Babuchowski, A.; Gajęcki, M.T. Correlations between Low Doses of Zearalenone, Its Carryover Factor and Estrogen Receptor Expression in Different Segments of the Intestines in Pre-Pubertal Gilts—A Study Protocol. Toxins 2021, 13, 379. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, Ł.; Waśkiewicz, A.; Beszterda, M.; Kostecki, M.; Dąbrowski, M.; Obremski, K.; Goliński, P.; Gajęcki, M. Zearalenone in the Intestinal Tissues of Immature Gilts Exposed per os to Mycotoxins. Toxins 2015, 7, 3210–3223. [Google Scholar] [CrossRef] [Green Version]
- Meerpoel, C.; Vidal, A.; Tangni, E.K.; Huybrechts, B.; Couck, L.; De Rycke, R.; De Bels, L.; De Saeger, S.; Van den Broeck, W.; Devreese, M.; et al. A Study of Carry-Over and Histopathological Effects after Chronic Dietary Intake of Citrinin in Pigs, Broiler Chickens and Laying Hens. Toxins 2020, 12, 719. [Google Scholar] [CrossRef]
- Kowalski, A.; Kaleczyc, J.; Gajęcki, M.; Zieliński, H. Adrenaline, noradrenaline and cortisol levels in pigs during blood collection. Med. Weter. 1996, 52, 716–718. (In Polish) [Google Scholar]
- Stanczyk, F.Z.; Xu, X.; Sluss, P.M.; Brinton, L.A.; McGlynn, K.A. Do metabolites account for higher serum steroid hormone levels measured by RIA compared to mass spectrometry? Clin. Chim. Acta 2018, 484, 223–225. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure Dates | Feed Intake [kg/day] | Total ZEN Doses in Groups [µg/kg BW] | Tissue | Group ZEN5 [ng/g] | Group ZEN10 [ng/g] | Group ZEN15 [ng/g] |
|---|---|---|---|---|---|---|
| D1 | 0.8 | 80.5/161.9/242.7 | Ovaries | 83.15 ± 99.60 | 54.29 ± 51.37 | 451.67 ± 433.14 |
| Uterine horn, ovarian section | 10.71 ± 5.13 xx | 7.46 ± 3.24 xx | 55.00 ± 17.00 | |||
| Uterine horn, uterine section | 7.47 ± 1.52 x | 11.44 ± 5.24 | 62.08 ± 51.33 | |||
| Middle part of the cervix | 18.06 ± 5.07 xx | 3.93 ± 0.93 xx yy | 75.19 ± 5.77 | |||
| Hypothalamus | 4.88 ± 2.13 x | 4.43 ± 2.54 x | 16.76 ± 9.44 | |||
| Pituitary gland | 13.77 | 12.66 | 14.26 | |||
| D2 | 1.1 | 101.01/196.9/298.2 | Ovary | 33.40 ± 26.09 | 79.67 ± 19.94 | 194.56 ± 138.81 |
| Uterine horn, ovarian section | 3.20 ± 3.32 a | 2.03 ± 0.88 a x | 13.40 ± 10.66 aa | |||
| Uterine horn, uterine section | 3.67 ± 1.95 a | 2.20 ± 0.53 aa, x | 7.41 ± 4.60 | |||
| Middle part of the cervix | 4.17 ± 3.11 aa xx | 4.46 ± 2.35 b | 45.35 ± 26.09 | |||
| Hypothalamus | 4.39 ± 2.81 x | 2.38 ± 0.81 x | 17.07 ± 11.52 | |||
| Pituitary gland | 6.81 | 8.63 | 9.02 | |||
| D3 | 1.6 | 128.3/481.4/716.7 | Ovary | 16.80 ± 15.69 xx | 29.06 ± 17.79 xx | 110.38 ± 26.91 |
| Uterine horn, ovarian section | 2.80 ± 1.71 a xx | 3.60 ± 1.84 x | 10.17 ± 2.37 aa | |||
| Uterine horn, uterine section | 3.17 ± 1.80 aa | 2.73 ± 1.19 a | 2.47 ± 1.51 | |||
| Middle part of the cervix | 1.51 ± 0.67 aa xx | 4.86 ± 2.89 xx | 23.17 ± 5.91 a | |||
| Hypothalamus | 3.45 ± 1.20 | 6.93 ± 5.65 | 10.02 ± 2.72 | |||
| Pituitary gland | 7.28 | 6.10 | 9.25 |
| Exposure Dates | Feed Intake [kg/day] | Total ZEN Doses in Groups [µg/kg BW] | Tissue | Group ZEN5 [ng/g] | Group ZEN10 [ng/g] | Group ZEN15 [ng/g] |
|---|---|---|---|---|---|---|
| D1 | 0.8 | 80.5/161.9/242.7 | Ovary | 0 | 0.51 ± 0.34 | 1.73 ± 0.45 |
| Uterine horn, ovarian section | 0 | 1.48 ± 0.98 | 0.94 ± 0.25 | |||
| Uterine horn, uterine section | 0 | 0.50 ± 0.28 | 1.65 ± 0.18 | |||
| Middle part of the cervix | 0 | 4.03 ± 0.16 | 1.77 ± 1.46 | |||
| Hypothalamus | 0 | 0.30 ± 0.20 | 0.94 ± 0.04 | |||
| Pituitary gland | 0 | 0 | 0 | |||
| D2 | 1.1 | 101.01/196.9/298.2 | Ovary | 1.04 ± 0.10 xx yy | 2.65 ± 0.55 aa | 3.00 ± 0.04 a |
| Uterine horn, ovarian section | 1.04 ± 0.42 yy | 3.02 ± 0.19 | 1.90 ± 0.87 | |||
| Uterine horn, uterine section | 0.51 ± 0.22 xx yy | 2.67 ± 0.42 aa | 1.95 ± 0.48 | |||
| Middle part of the cervix | 4.89 ± 0.60 x | 3.91 ± 0.08 | 3.48 ± 0.48 | |||
| Hypothalamus | 0.29 ± 0.20 x | 0.50 ± 0.03 | 0.65 ± 0.08 a | |||
| Pituitary gland | 0 | 0 | 0.246 | |||
| D3 | 1.6 | 128.3/481.4/716.7 | Ovary | 3.16 ± 0.73 | 3.23 ± 0.38 aa | 3.38 ± 0.54 aa |
| Uterine horn, ovarian section | 3.61 ± 0.16 | 3.91 ± 1.66 | 4.00 ± 0.32 aa, bb | |||
| Uterine horn, uterine section | 3.83 ± 0.17 | 4.24 ± 0.47 aa bb | 3.02 ± 1.80 | |||
| Middle part of the cervix | 3.13 ± 0.19 xx yy | 1.83 ± 0.17 aa bb | 2.81 ± 0.27 | |||
| Hypothalamus | 0.43 ± 0.29 x | 0.65 ± 0.08 | 0.79 ± 0.15 | |||
| Pituitary gland | 0 | 0.195 | 0.245 |
| Exposure Date | Feed Intake [kg/day] | Total ZEN Doses in Groups [µg/kg BW] | Tissue | Group ZEN5 [ng/g] | Group ZEN10 [ng/g] | Group ZEN15 [ng/g] |
|---|---|---|---|---|---|---|
| D1 | 0.8 | 80.5/161.9/242.7 | Ovary | 0 | 1.07 ± 0.29 | 0.98 ± 0.12 |
| Uterine horn, ovarian section | 0 | 0.36 ± 0.07 | 0.87 ± 0.14 | |||
| Uterine horn, uterine section | 0 | 0.41 ± 0.08 | 0.93 ± 0.03 | |||
| Middle part of the cervix | 0 | 0.42 ± 0.04 | 1.22 ± 0.02 | |||
| Hypothalamus | 0 | 0 | 0 | |||
| Pituitary gland | 0 | 0 | 0 | |||
| D2 | 1.1 | 101.01/196.9/298.2 | Ovary | 0.92 ± 0.10 | 1.05 ± 0.53 | 0.87 ± 0.06 xx |
| Uterine horn, ovarian section | 0.02 ± 0.02 | 0.12 ± 0.08 aa | 0.09 ± 0.01 aa | |||
| Uterine horn, uterine section | 0.01 ± 0.01 | 0.29 ± 0.57 | 0.02 ± 0.02 aa | |||
| Middle part of the cervix | 0.05 ± 0.01 | 0.04 ± 0.01 aa | 0.06 ± 0.02 aa | |||
| Hypothalamus | 0.09 ± 0.13 | 0.04 ± 0.009 | 0.006 ± 0.01 | |||
| Pituitary gland | 0 | 0.372 | 0.462 | |||
| D3 | 1.6 | 128.3/481.4/716.7 | Ovary | 0.11 ± 0.12 xx yy | 1.48 ± 0.25 | 1.49 ± 0.27 a |
| Uterine horn, ovarian section | 0.01 ± 0.005 xx yy | 0.01 ± 0.002 aa | 0.14 ± 0.02 aa | |||
| Uterine horn, uterine section | 0.03 ± 0.01 | 0.14 ± 0.18 | 0.05 ± 0.03 aa | |||
| Middle part of the cervix | 0.02 ± 0.01 x yy | 0.07 ± 0.02 aa z | 0.12 ± 0.02 aa | |||
| Hypothalamus | 0.04 ± 0.03 yy | 0.04 ± 0.01 zz | 0.21 ± 0.03 | |||
| Pituitary gland | 0.253 | 0.362 | 0.536 |
| Parameters | Composition Declared by The Manufacturer (%) |
|---|---|
| Soybean meal | 16 |
| Wheat | 55 |
| Barley | 22 |
| Wheat bran | 4.0 |
| Limestone | 0.3 |
| Zitrosan | 0.2 |
| Vitamin-mineral premix 1 | 2.5 |
| Analyte | Precursor | Quantification Ion | Confirmation | Ion LOD (ng mL−1) | LOQ (ng mL−1) | Linearity (%R2) |
|---|---|---|---|---|---|---|
| ZEN | 317.1 | 273.3 | 187.1 | 0.03 | 0.1 | 0.999 |
| α-ZEL | 319.2 | 275.2 | 160.1 | 0.3 | 0.9 | 0.997 |
| β-ZEL | 319.2 | 275.2 | 160.1 | 0.3 | 1 | 0.993 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gajęcka, M.; Zielonka, Ł.; Babuchowski, A.; Gajęcki, M.T. Exposure to Low Zearalenone Doses and Changes in the Homeostasis and Concentrations of Endogenous Hormones in Selected Steroid-Sensitive Tissues in Pre-Pubertal Gilts. Toxins 2022, 14, 790. https://doi.org/10.3390/toxins14110790
Gajęcka M, Zielonka Ł, Babuchowski A, Gajęcki MT. Exposure to Low Zearalenone Doses and Changes in the Homeostasis and Concentrations of Endogenous Hormones in Selected Steroid-Sensitive Tissues in Pre-Pubertal Gilts. Toxins. 2022; 14(11):790. https://doi.org/10.3390/toxins14110790
Chicago/Turabian StyleGajęcka, Magdalena, Łukasz Zielonka, Andrzej Babuchowski, and Maciej Tadeusz Gajęcki. 2022. "Exposure to Low Zearalenone Doses and Changes in the Homeostasis and Concentrations of Endogenous Hormones in Selected Steroid-Sensitive Tissues in Pre-Pubertal Gilts" Toxins 14, no. 11: 790. https://doi.org/10.3390/toxins14110790
APA StyleGajęcka, M., Zielonka, Ł., Babuchowski, A., & Gajęcki, M. T. (2022). Exposure to Low Zearalenone Doses and Changes in the Homeostasis and Concentrations of Endogenous Hormones in Selected Steroid-Sensitive Tissues in Pre-Pubertal Gilts. Toxins, 14(11), 790. https://doi.org/10.3390/toxins14110790