Combining Nanopore Sequencing with Recombinase Polymerase Amplification Enables Identification of Dinoflagellates from the Alexandrium Genus, Providing a Rapid, Field Deployable Tool

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Method Development
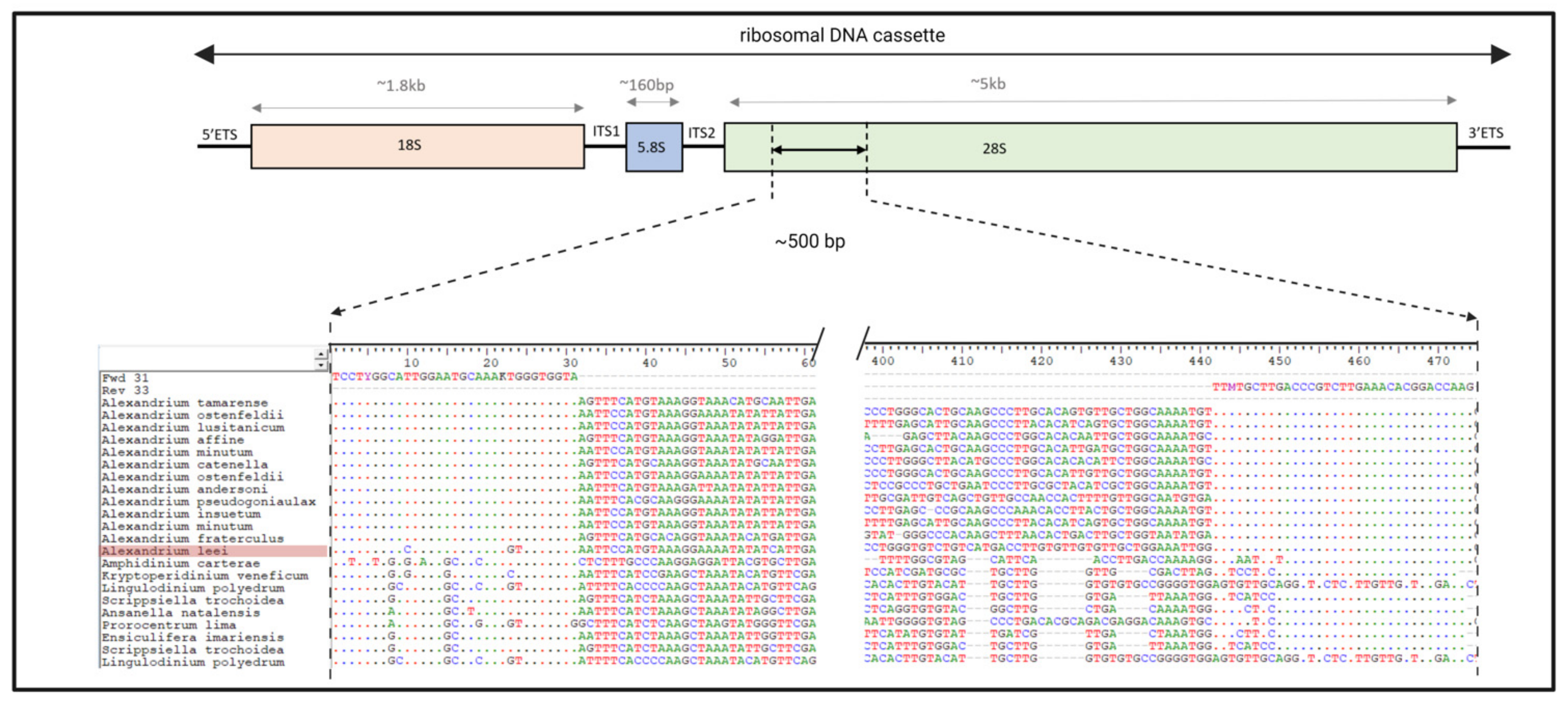
2.1.1. Primer Design
2.1.2. RPA Performance
2.2. Method Optimization
2.2.1. Sensitivity
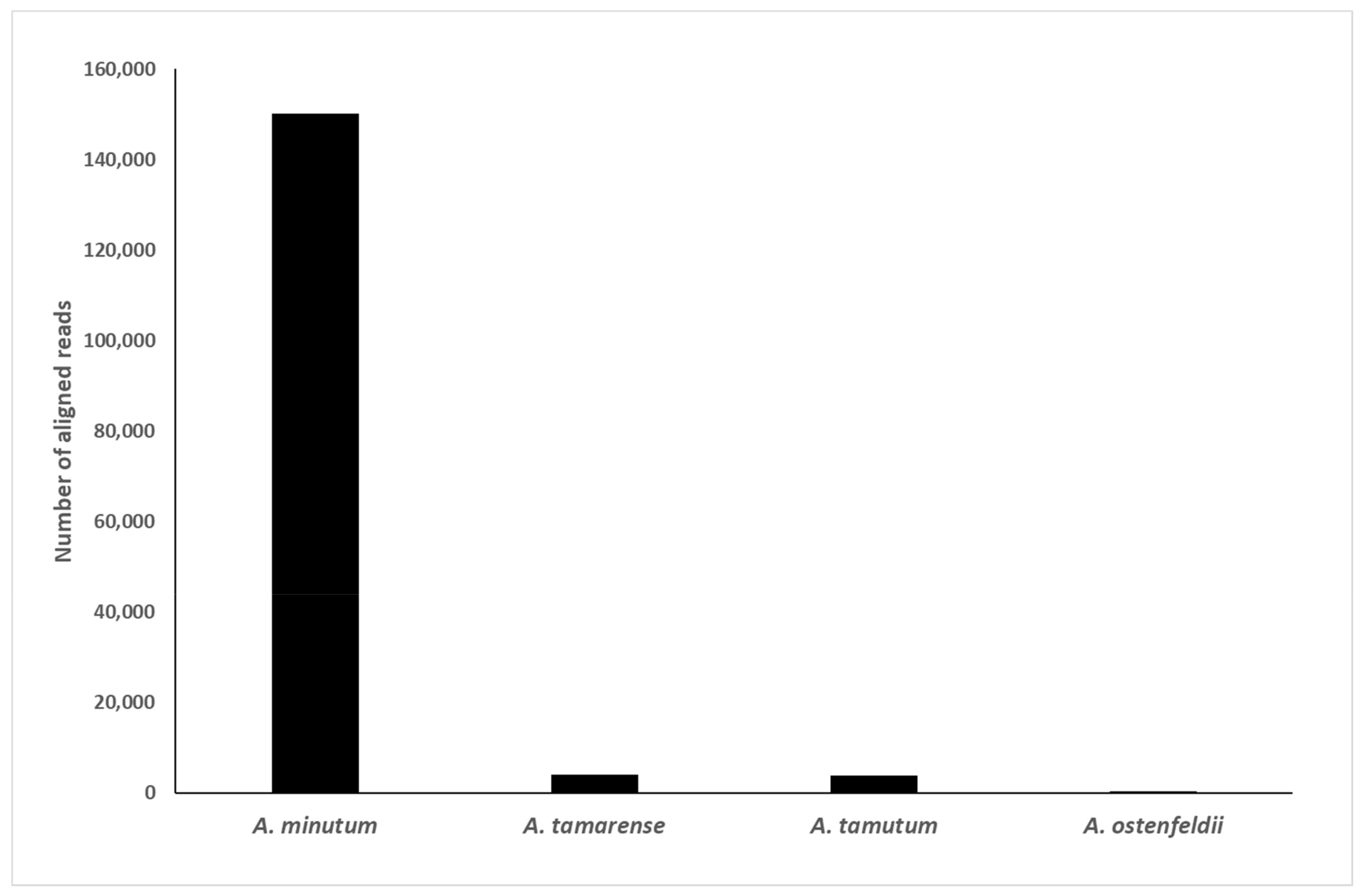
2.2.2. Specificity and Selectivity
2.3. Real Time Alignment and Phylogenetic Tree Analysis
2.3.1. Real Time Alignment
2.3.2. Phylogenetic Tree Analysis
2.4. Library Preparation and Multiplexing
2.4.1. Oxford Nanopore Field Sequencing Kit
2.4.2. Oxford Nanopore Rapid PCR Barcode Kit
2.4.3. VolTRAX Multiplexing Kit
2.5. Field Deployable Tools
2.5.1. In Field Cell Lysis
2.5.2. Bento Lab
2.5.3. The XavION Sequencer
2.6. Environmental Samples
3. Materials and Methods
3.1. Biological Material and Sample Preparation
3.1.1. Reference Materials
3.1.2. Sample Concentration
3.1.3. Performance Testing
3.1.4. Cell Lysis
3.2. RPA Assay
3.2.1. Primer Design
3.2.2. RPA Reactions
3.2.3. DNA Quantitation and Electrophoretic Analysis
3.2.4. Specificity Testing
3.3. Library Preparation and Nanopore Sequencing
3.3.1. Library Preparation Tools
3.3.2. Sequencing Device
3.3.3. Field Sequencing Library Preparation Kit
3.3.4. Rapid PCR Barcode Library Preparation Kit
3.3.5. VolTRAX Multiplexing Library Preparation Kit
3.4. Case Study: Environmental Samples
3.5. Bioinformatic and Statistical Analysis
3.5.1. Bioinformatics
“NGSpeciesID - m 400 --s 200 --mapped_threshold 0.8 --aligned_threshold 0.8 --rc_identity_threshold 0.8 --abundance_ratio 0.01 --ont --consensus --t 20 --fastq input.fastq --outfolder output_folder --medaka --medaka_model r941_min_high_g360”
3.5.2. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brierley, A.S. Plankton. Curr. Biol. 2017, 27, R478–R483. [Google Scholar] [CrossRef] [PubMed]
- Nwankwegu, A.S.; Li, Y.; Huang, Y.; Wei, J.; Norgbey, E.; Sarpong, L.; Lai, Q.; Wang, K. Harmful algal blooms under changing climate and constantly increasing anthropogenic actions: The review of management implications. 3 Biotech 2019, 9, 449. [Google Scholar] [CrossRef] [PubMed]
- Hallegraeff, G.; Enevoldsen, H.; Zingone, A. Global harmful algal bloom status reporting. Harmful Algae 2021, 102, 101992. [Google Scholar] [CrossRef] [PubMed]
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. Harmful algal blooms: A global overview. Man. Harmful Mar. Microalgae 2003, 33, 1–22. [Google Scholar] [CrossRef]
- Batista, F.M.; Hatfield, R.; Powell, A.; Baker-Austin, C.; Lowther, J.; Turner, A.D. Methodological advances in the detection of biotoxins and pathogens affecting production and consumption of bivalve molluscs in a changing environment. Curr. Opin. Biotechnol. 2023, 80, 102896. [Google Scholar] [CrossRef]
- Klemm, K.; Cembella, A.; Clarke, D.; Cusack, C.; Arneborg, L.; Karlson, B.; Liu, Y.; Naustvoll, L.; Siano, R.; Gran-Stadniczeñko, S.; et al. Apparent biogeographical trends in Alexandrium blooms for northern Europe: Â identifying links to climate change and effective adaptive actions. Harmful Algae 2022, 119, 102335. [Google Scholar] [CrossRef]
- Mertens, K.N.; Adachi, M.; Anderson, D.M.; Band-Schmidt, C.J.; Bravo, I.; Brosnahan, M.L.; Bolch, C.J.; Calado, A.J.; Carbonell-Moore, M.C.; Chomérat, N.; et al. Morphological and phylogenetic data do not support the split of Alexandrium into four genera. Harmful Algae 2020, 98, 101902. [Google Scholar] [CrossRef]
- Fraga, S.; Sampedro, N.; Larsen, J.; Moestrup, Ø.; Calado, A.J. Arguments against the proposal 2302 by John & al. To reject the name Gonyaulax catenella (Alexandrium catenella). Taxon 2015, 64, 634–635. [Google Scholar] [CrossRef]
- John, U.; Litaker, R.W.; Montresor, M.; Murray, S.; Brosnahan, M.L.; Anderson, D.M. Formal revision of the alexandrium tamarense species complex (dinophyceae) taxonomy: The introduction of five species with emphasis on molecular-based (rDNA) classification. Protist 2014, 165, 779–804. [Google Scholar] [CrossRef]
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The globally distributed genus Alexandrium: Multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef]
- Llewellyn, L.E. Saxitoxin, a toxic marine natural product that targets a multitude of receptors. Nat. Prod. Rep. 2006, 23, 200. [Google Scholar] [CrossRef]
- Grattan, L.M.; Holobaugh, S.; Morris, J.G. Harmful algal blooms and public health. Harmful Algae 2016, 57, 2–8. [Google Scholar] [CrossRef]
- Sierra, M.A.; Martínez-Álvarez, R. Ricin and Saxitoxin: Two Natural Products That Became Chemical Weapons. J. Chem. Educ. 2020, 97, 1707–1714. [Google Scholar] [CrossRef]
- Stauffer, B.A.; Bowers, H.A.; Buckley, E.; Davis, T.W.; Johengen, T.H.; Kudela, R.; McManus, M.A.; Purcell, H.; Smith, G.J.; Woude, A.V.; et al. Considerations in harmful algal bloom research and monitoring: Perspectives from a consensus-building workshop and technology testing. Front. Mar. Sci. 2019, 6, 399. [Google Scholar] [CrossRef]
- Turner, A.D.; Hatfield, R.G.; Maskrey, B.H.; Algoet, M.; Lawrence, J.F. Evaluation of the new European Union reference method for paralytic shellfish toxins in shellfish: A review of twelve years regulatory monitoring using pre-column oxidation LC-FLD. TrAC Trends Anal. Chem. 2019, 113, 124–139. [Google Scholar] [CrossRef]
- Joint FAO-IOC-IAEA technical guidance for the implementation of early warning systems for harmful algal blooms. FAO Fish. Aquac. Technol. Pap. 2023, 690, I-202. [CrossRef]
- AOAC. AOAC Official Method 2005.06 Paralytic Shellfish Poisoning Toxins in Shellfish. AOAC: Rockville, MD, USA, 2005; pp. 1714–1727. [Google Scholar]
- DeGrasse, S.L.; van de Riet, J.; Hatfield, R.; Turner, A. Pre- versus post-column oxidation liquid chromatography fluorescence detection of paralytic shellfish toxins. Toxicon 2011, 57, 619–624. [Google Scholar] [CrossRef]
- Hatfield, R.G.; Turner, A.D. Rapid liquid chromatography for paralytic shellfish toxin analysis using superficially porous chromatography with AOAC official methodSM 2005.06. J. AOAC Int. 2012, 95, 1089–1096. [Google Scholar] [CrossRef]
- Turner, A.D.; Dhanji-Rapkova, M.; Fong, S.Y.T.; Hungerford, J.; McNabb, P.S.; Boundy, M.J.; Harwood, D.T.; Aanrud, S.; Alfonso, C.; Alvarez, M.; et al. Ultrahigh-Performance Hydrophilic Interaction Liquid Chromatography with Tandem Mass Spectrometry Method for the Determination of Paralytic Shellfish Toxins and Tetrodotoxin in Mussels, Oysters, Clams, Cockles, and Scallops: Collaborative Study. J. AOAC Int. 2020, 103, 533–562. [Google Scholar] [CrossRef]
- Utermöhl, H. Neue Wege in der quantitativen Erfassung des Plankton. (Mit besonderer Berücksichtigung des Ultraplanktons.). SIL Proc. 1922–2010 1931, 5, 567–596. [Google Scholar] [CrossRef]
- UK-NRL PhytoplanktonWG. Standard Operating Procedure for the Identification and Enumeration of Potential Toxin-Producing Phytoplankton Species in Samples Collected from UK Coastal Waters Using the Utermöhl Method. 2008. Vol. Version 4, no. March, pp. 1–10. Available online: https://www.afbini.gov.uk (accessed on 25 April 2023).
- Rodríguez-Ramos, T.; Dornelas, M.; Marañón, E.; Cermeño, P. Conventional sampling methods severely underestimate phytoplankton species richness. J. Plankton Res. 2014, 36, 334–343. [Google Scholar] [CrossRef]
- Rowland-Pilgrim, S.; Swan, S.C.; O’neill, A.; Johnson, S.; Coates, L.; Stubbs, P.; Dean, K.; Parks, R.; Harrison, K.; Alves, M.T.; et al. Variability of Amnesic Shellfish Toxin and Pseudo-nitzschia occurrence in bivalve molluscs and water samples–Analysis of ten years of the official control monitoring programme. Harmful Algae 2019, 87, 101623. [Google Scholar] [CrossRef] [PubMed]
- Eckford-Soper, L.K.; Bresnan, E.; Lacaze, J.-P.; Green, D.H.; Davidson, K. The competitive dynamics of toxic Alexandrium fundyense and non-toxic Alexandrium tamarense: The role of temperature. Harmful Algae 2016, 53, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhang, C.; Wang, Y.; Chen, G. A review of the current and emerging detection methods of marine harmful microalgae. Sci. Total Environ. 2022, 815, 152913. [Google Scholar] [CrossRef]
- Hatfield, R.G.; Bean, T.; Turner, A.D.; Lees, D.N.; Lowther, J.; Lewis, A.; Baker-Austin, C. Development of a TaqMan qPCR assay for detection of Alexandrium spp. and application to harmful algal bloom monitoring. Toxicon X 2019, 2, 100011. [Google Scholar] [CrossRef]
- Hatfield, R.G.; Batista, F.M.; Bean, T.P.; Fonseca, V.G.; Santos, A.; Turner, A.D.; Lewis, A.; Dean, K.J.; Martinez-Urtaza, J. The Application of Nanopore Sequencing Technology to the Study of Dinoflagellates: A Proof of Concept Study for Rapid Sequence-Based Discrimination of Potentially Harmful Algae. Front. Microbiol. 2020, 11, 844. [Google Scholar] [CrossRef]
- Opel, K.L.; Chung, D.; Mccord, B.R. A study of PCR inhibition mechanisms using real time PCR. J. Forensic Sci. 2010, 55, 25–33. [Google Scholar] [CrossRef]
- Lobato, I.M.; O’Sullivan, C.K. Recombinase polymerase amplification: Basics, applications and recent advances. TrAC Trends Anal. Chem. 2017, 98, 19–35. [Google Scholar] [CrossRef]
- Toldrà, A.; O’Sullivan, C.K.; Diogène, J.; Campàs, M. Detecting harmful algal blooms with nucleic acid amplification-based biotechnological tools. Sci. Total Environ. 2020, 749, 141605. [Google Scholar] [CrossRef]
- Valentini, A.; Taberlet, P.; Miaud, C.; Civade, R.; Herder, J.; Thomsen, P.F.; Bellemain, E.; Besnard, A.; Coissac, E.; Boyer, F.; et al. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Mol. Ecol. 2016, 25, 929–942. [Google Scholar] [CrossRef]
- Tan, B.F.; Ng, C.M.; Nshimyimana, J.P.; Loh, L.L.; Gin, K.Y.-H.; Thompson, J.R. Next-generation sequencing (NGS) for assessment of microbial water quality: Current progress, challenges, and future opportunities. Front. Microbiol. 2015, 6, 1027. [Google Scholar] [CrossRef]
- Esenkulova, S.; Sutherland, B.J.; Tabata, A.; Haigh, N.; Pearce, C.M.; Miller, K.M. Operational taxonomic unit comparing metabarcoding and morphological approaches to identify phytoplankton taxa associated with harmful algal blooms. Facets 2020, 5, 784–811. [Google Scholar] [CrossRef]
- Shendure, J.; Balasubramanian, S.; Church, G.M.; Gilbert, W.; Rogers, J.; Schloss, J.A.; Waterston, R.H. DNA sequencing at 40: Past, present and future. Nature 2017, 550, 345–353. [Google Scholar] [CrossRef]
- Mikheyev, A.S.; Tin, M.M.Y. A first look at the Oxford Nanopore MinION sequencer. Mol. Ecol. Resour. 2014, 14, 1097–1102. [Google Scholar] [CrossRef]
- Deamer, D.; Akeson, M.; Branton, D. Three decades of nanopore sequencing. Nat. Biotechnol. 2016, 34, 518–524. [Google Scholar] [CrossRef]
- Laver, T.; Harrison, J.; O’neill, P.; Moore, K.; Farbos, A.; Paszkiewicz, K.; Studholme, D. Assessing the performance of the Oxford Nanopore Technologies MinION. Biomol. Detect. Quantif. 2015, 3, 1–8. [Google Scholar] [CrossRef]
- Sanderson, N.; Kapel, N.; Rodger, G.; Webster, H.; Lipworth, S.; Stoesser, N. Comparison of R9.4.1/Kit10 and R10/Kit12 Oxford Nanopore flowcells and chemistries in bacterial genome reconstruction. Biorxiv Prepr. 2023, 9, mgen000910. [Google Scholar] [CrossRef]
- Johnson, S.S.; Zaikova, E.; Goerlitz, D.S.; Bai, Y.; Tighe, S.W. Real-time DNA sequencing in the antarctic dry valleys using the Oxford nanopore sequencer. J. Biomol. Tech. 2017, 28, 2–7. [Google Scholar] [CrossRef]
- Parker, J.; Helmstetter, A.J.; Devey, D.; Wilkinson, T.; Papadopulos, A.S.T. Field-based species identification of closely-related plants using real-time nanopore sequencing. Sci. Rep. 2017, 7, 8345. [Google Scholar] [CrossRef]
- Shikata, T.; Taniguchi, E.; Sakamoto, S.; Kitatsuji, S.; Yamasaki, Y.; Yoshida, M.; Oikawa, H. Phylogeny, growth and toxicity of the noxious red-tide dinoflagellate Alexandrium leei in Japan. Reg. Stud. Mar. Sci. 2020, 36, 101265. [Google Scholar] [CrossRef]
- Zhai, Y.; Li, R.; Liu, F.; Zhang, C.; Wang, Y.; Chen, G. Recombinase polymerase amplification combined with lateral flow dipstick for the rapid detection of Prorocentrum donghaiense. Mar. Biol. Res. 2021, 17, 646–657. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, C.; Liu, F.; Li, R.; Wang, Y.; Chen, G. Development of a recombinase polymerase amplification combined with lateral flow dipstick assay for rapid and sensitive detection of Heterosigma akashiwo. J. Appl. Phycol. 2021, 33, 3165–3178. [Google Scholar] [CrossRef]
- Fu, M.; Chen, G.; Zhang, C.; Wang, Y.; Sun, R.; Zhou, J. Rapid and sensitive detection method for Karlodinium veneficum by recombinase polymerase amplification coupled with lateral flow dipstick. Harmful Algae 2019, 84, 1–9. [Google Scholar] [CrossRef]
- Toldrà, A.; Jauset-Rubio, M.; Andree, K.B.; Fernández-Tejedor, M.; Diogène, J.; Katakis, I.; O’Sullivan, C.K.; Campàs, M. Detection and quantification of the toxic marine microalgae Karlodinium veneficum and Karlodinium armiger using recombinase polymerase amplification and enzyme-linked oligonucleotide assay. Anal. Chim. Acta 2018, 1039, 140–148. [Google Scholar] [CrossRef]
- Oosterbroek, S.; Doorenspleet, K.; Nijland, R.; Jansen, L. Decona: From demultiplexing to consensus for Nanopore amplicon data. ARPHA Conf. Abstr. 2021, 4, e65029. [Google Scholar] [CrossRef]
- Chen, P.; Sun, Z.; Wang, J.; Liu, X.; Bai, Y.; Chen, J.; Liu, A.; Qiao, F.; Chen, Y.; Yuan, C.; et al. Portable nanopore-sequencing technology: Trends in development and applications. Front. Microbiol. 2023, 14, 1043967. [Google Scholar] [CrossRef]
- Martin, S.; Heavens, D.; Lan, Y.; Horsfield, S.; Clark, M.D.; Leggett, R.M. Nanopore adaptive sampling: A tool for enrichment of low abundance species in metagenomic samples. Genome Biol. 2022, 23, 11. [Google Scholar] [CrossRef]
- Lin, Y.; Dai, Y.; Liu, Y.; Ren, Z.; Guo, H.; Li, Z.; Li, J.; Wang, K.; Yang, L.; Zhang, S.; et al. Rapid PCR-Based Nanopore Adaptive Sequencing Improves Sensitivity and Timeliness of Viral Clinical Detection and Genome Surveillance. Front. Microbiol. 2022, 13, e2102593. [Google Scholar] [CrossRef]
- Brown, L.; Bresnan, E.; Graham, J.; Lacaze, J.-P.; Turrell, E.; Collins, C. Distribution, diversity and toxin composition of the genus alexandrium (dinophyceae) in Scottish waters. Eur. J. Phycol. 2010, 45, 375–393. [Google Scholar] [CrossRef]
- Van Reine, W.P. Report of the nomenclature committee for algae: 15. Taxon 2017, 66, 191–192. [Google Scholar] [CrossRef]
- Anon. Commission Regulation (EC) No 1664/2006 of 6 November 2006 amending Regulation (EC) No 2074/2005 as regards implementing measures for certain products of animal origin intended for human consumption and repealing certain implementing measures. Off. J. Eur. Union 2006, 320, 13–45. [Google Scholar]
- Turner, A.; Dhanji-Rapkova, M.; Baker, C.; Algoet, M. Assessment of a semiquantitative liquid chromatography-fluorescence detection method for the determination of paralytic shellfish poisoning toxin levels in bivalve molluscs from Great Britain. J. AOAC Int. 2014, 97, 492–497. [Google Scholar] [CrossRef]
- Montresor, M.; John, U.; Beran, A.; Medlin, L.K. Alexandrium tamutum sp. nov. (Dinophyceae): A new nontoxic species in the genus Alexandrium. J. Phycol. 2004, 40, 398–411. [Google Scholar] [CrossRef]
- Salgado, P.; Riobó, P.; Rodríguez, F.; Franco, J.M.; Bravo, I. Differences in the toxin profiles of Alexandrium ostenfeldii (Dinophyceae) strains isolated from different geographic origins: Evidence of paralytic toxin, spirolide, and gymnodimine. Toxicon 2015, 103, 85–98. [Google Scholar] [CrossRef]
- Lewis, A.M.; Coates, L.N.; Turner, A.D.; Percy, L.; Lewis, J. A review of the global distribution of Alexandrium minutum (Dinophyceae) and comments on ecology and associated paralytic shellfish toxin profiles, with a focus on Northern Europe. J. Phycol. 2018, 54, 581–598. [Google Scholar] [CrossRef]
- Lewis, A.M.; Dean, K.J.; Hartnell, D.M.; Percy, L.; Turner, A.D.; Lewis, J.M. The value of toxin profiles in the chemotaxonomic analysis of paralytic shellfish toxins in determining the relationship between British Alexandrium spp. and experimentally contaminated Mytilus sp. Harmful Algae 2022, 111, 102131. [Google Scholar] [CrossRef]
- Turner, A.D.; Stubbs, B.; Coates, L.; Dhanji-Rapkova, M.; Hatfield, R.G.; Lewis, A.M.; Rowland-Pilgrim, S.; O’neil, A.; Stubbs, P.; Ross, S.; et al. Variability of paralytic shellfish toxin occurrence and profiles in bivalve molluscs from Great Britain from official control monitoring as determined by pre-column oxidation liquid chromatography and implications for applying immunochemical tests. Harmful Algae 2014, 31, 87–99. [Google Scholar] [CrossRef]
- Smith, M.Q.R.P.; Ruxton, G.D. Effective use of the McNemar test. Behav. Ecol. Sociobiol. 2020, 74, 133. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Hargraves, P.E. Stichochrysis immobilis is a diatom, not a chrysophyte. Phycologia 1993, 32, 234–236. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. 1999, 41, 95–98. [Google Scholar]
- TwistDX Primer Design Protocol. Available online: https://www.twistdx.co.uk/wp-content/uploads/2021/04/twistamp-assay-design-manual-v2-5.pdf (accessed on 3 November 2022).
- Chen, Y.; Ye, W.; Zhang, Y.; Xu, Y. High speed BLASTN: An accelerated MegaBLAST search tool. Nucleic Acids Res. 2015, 43, 7762–7768. [Google Scholar] [CrossRef] [PubMed]
- Benton, M. Zenodo/Github, version 0.2.1, Nanopore Sequencing on Nvidia Jetson SoM boards. GitHub: San Francisco, CA, USA, 2021.
- Hatfield, R.G.; Punn, R.; Algoet, M.; Turner, A.D. A rapid method for the analysis of paralytic shellfish toxins utilizing standard pressure HPLC: Refinement of AOAC 2005.06. J. AOAC Int. 2016, 99, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Sahlin, K.; Lim, M.C.W.; Prost, S. NGSpeciesID: DNA barcode and amplicon consensus generation from long-read sequencing data. Ecol. Evol. 2021, 11, 1392–1398. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primers: | |
| Fwd36: 5’ CACAYTCCTYGGCATTGGAATGCAAAKTGGGTGGTA 3’ | |
| Fwd35: 5’ ACAYTCCTYGGCATTGGAATGCAAAKTGGGTGGTA 3’ | |
| Fwd34: 5’ CAYTCCTYGGCATTGGAATGCAAAKTGGGTGGTA 3’ | |
| Fwd33: 5’ AYTCCTYGGCATTGGAATGCAAAKTGGGTGGTA 3’ | |
| Fwd32: 5’ YTCCTYGGCATTGGAATGCAAAKTGGGTGGTA 3’ | |
| * | Fwd31: 5’ TCCTYGGCATTGGAATGCAAAKTGGGTGGTA 3’ |
| Fwd30: 5’ CCTYGGCATTGGAATGCAAAKTGGGTGGTA 3’ | |
| Reverse Primers: | |
| Rev36: 5’ CTTGGTCCGTGTTTCAAGACGGGTCAAGCAKAADCA 3’ | |
| Rev35: 5’ CTTGGTCCGTGTTTCAAGACGGGTCAAGCAKAADC 3’ | |
| Rev34: 5’ CTTGGTCCGTGTTTCAAGACGGGTCAAGCAKAAD 3’ | |
| * | Rev33: 5’ CTTGGTCCGTGTTTCAAGACGGGTCAAGCAKAA 3’ |
| Rev32: 5’ CTTGGTCCGTGTTTCAAGACGGGTCAAGCAKA 3’ | |
| Rev31: 5’ CTTGGTCCGTGTTTCAAGACGGGTCAAGCAK 3’ | |
| Rev30: 5’ CTTGGTCCGTGTTTCAAGACGGGTCAAGCA 3’ |
| Experiment | Concentration Method | Mag Bead Clean Up | Alexandrium Cells Spiked | Amplification | ||
|---|---|---|---|---|---|---|
| A | B | C | ||||
| Sensitivity testing | Centrefugation | yes | 1000 | + | + | + |
| yes | 100 | + | + | + | ||
| yes | 10 | + | + | + | ||
| yes | 1 | + | + | − | ||
| yes | 0 | − | − | − | ||
| Filtration | yes | 1000 | + | + | + | |
| yes | 100 | + | + | + | ||
| yes | 10 | + | + | + | ||
| yes | 1 | + | + | + | ||
| yes | 0 | − | − | − | ||
| None * | yes | 1 | + | + | + | |
| yes | 0 | − | − | − | ||
| Live Cells | Centrifugation | yes | 100 | + | + | + |
| Mock community | Centrifugation | yes | 100 | + | ||
| yes | 0 | − | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatfield, R.G.; Ryder, D.; Tidy, A.M.; Hartnell, D.M.; Dean, K.J.; Batista, F.M. Combining Nanopore Sequencing with Recombinase Polymerase Amplification Enables Identification of Dinoflagellates from the Alexandrium Genus, Providing a Rapid, Field Deployable Tool. Toxins 2023, 15, 372. https://doi.org/10.3390/toxins15060372
Hatfield RG, Ryder D, Tidy AM, Hartnell DM, Dean KJ, Batista FM. Combining Nanopore Sequencing with Recombinase Polymerase Amplification Enables Identification of Dinoflagellates from the Alexandrium Genus, Providing a Rapid, Field Deployable Tool. Toxins. 2023; 15(6):372. https://doi.org/10.3390/toxins15060372
Chicago/Turabian StyleHatfield, Robert G., David Ryder, Annabel M. Tidy, David M. Hartnell, Karl J. Dean, and Frederico M. Batista. 2023. "Combining Nanopore Sequencing with Recombinase Polymerase Amplification Enables Identification of Dinoflagellates from the Alexandrium Genus, Providing a Rapid, Field Deployable Tool" Toxins 15, no. 6: 372. https://doi.org/10.3390/toxins15060372
APA StyleHatfield, R. G., Ryder, D., Tidy, A. M., Hartnell, D. M., Dean, K. J., & Batista, F. M. (2023). Combining Nanopore Sequencing with Recombinase Polymerase Amplification Enables Identification of Dinoflagellates from the Alexandrium Genus, Providing a Rapid, Field Deployable Tool. Toxins, 15(6), 372. https://doi.org/10.3390/toxins15060372