Ciguatera Fish Poisoning in the Caribbean Sea and Atlantic Ocean: Reconciling the Multiplicity of Ciguatoxins and Analytical Chemistry Approach for Public Health Safety



Abstract
:1. Introduction
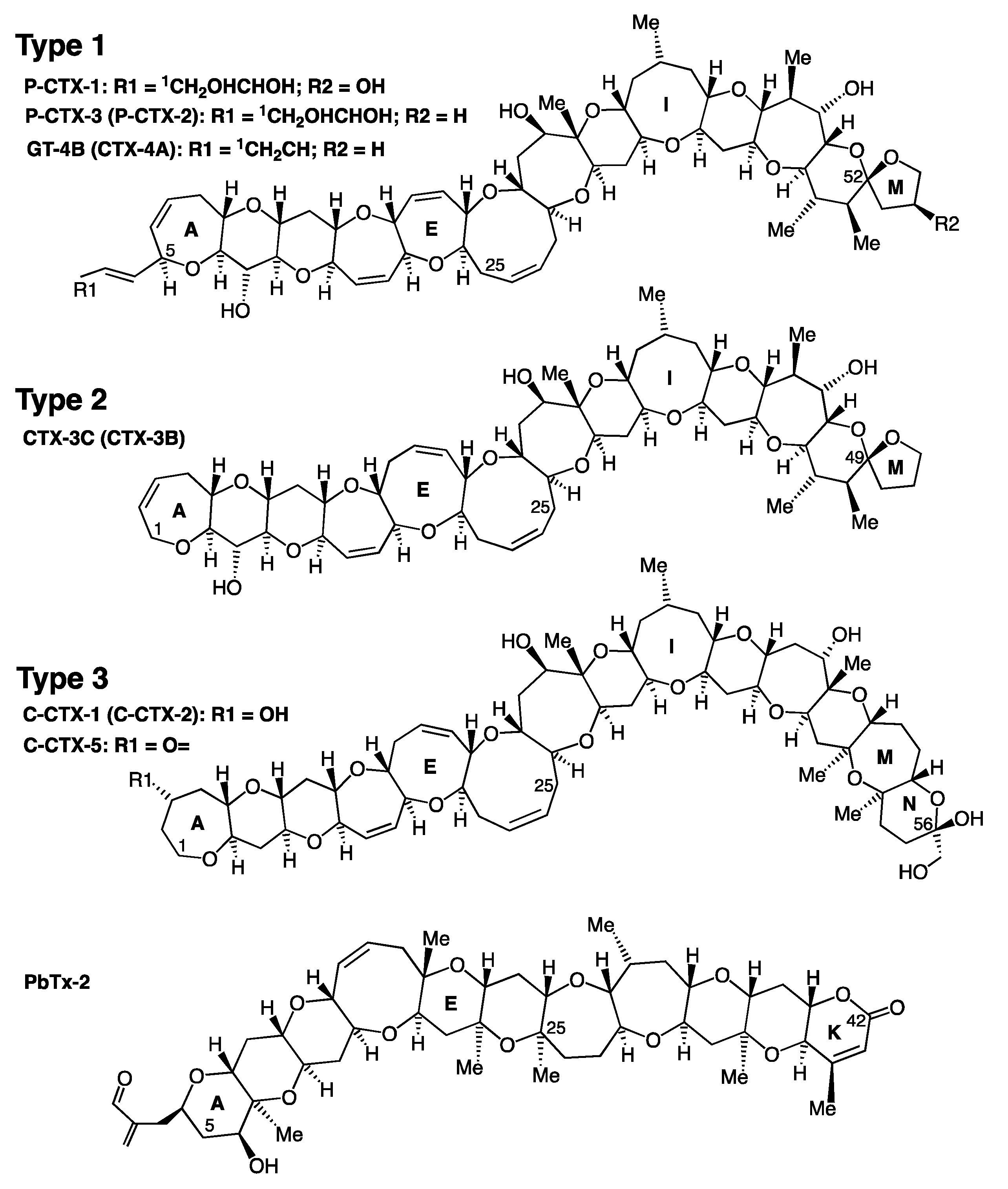
2. Ciguatoxins Structures and Reference Standards
2.1. Structures of the Three Main Types of Ciguatoxins
2.2. Availability of Ciguatoxin Reference Standards
3. Links between the Chemistry and Biochemistry and Toxicology of CTXs
3.1. Lipophilicity of CTXs
3.2. Distribution of C-CTXs in Fish
3.3. Stability of CTXs
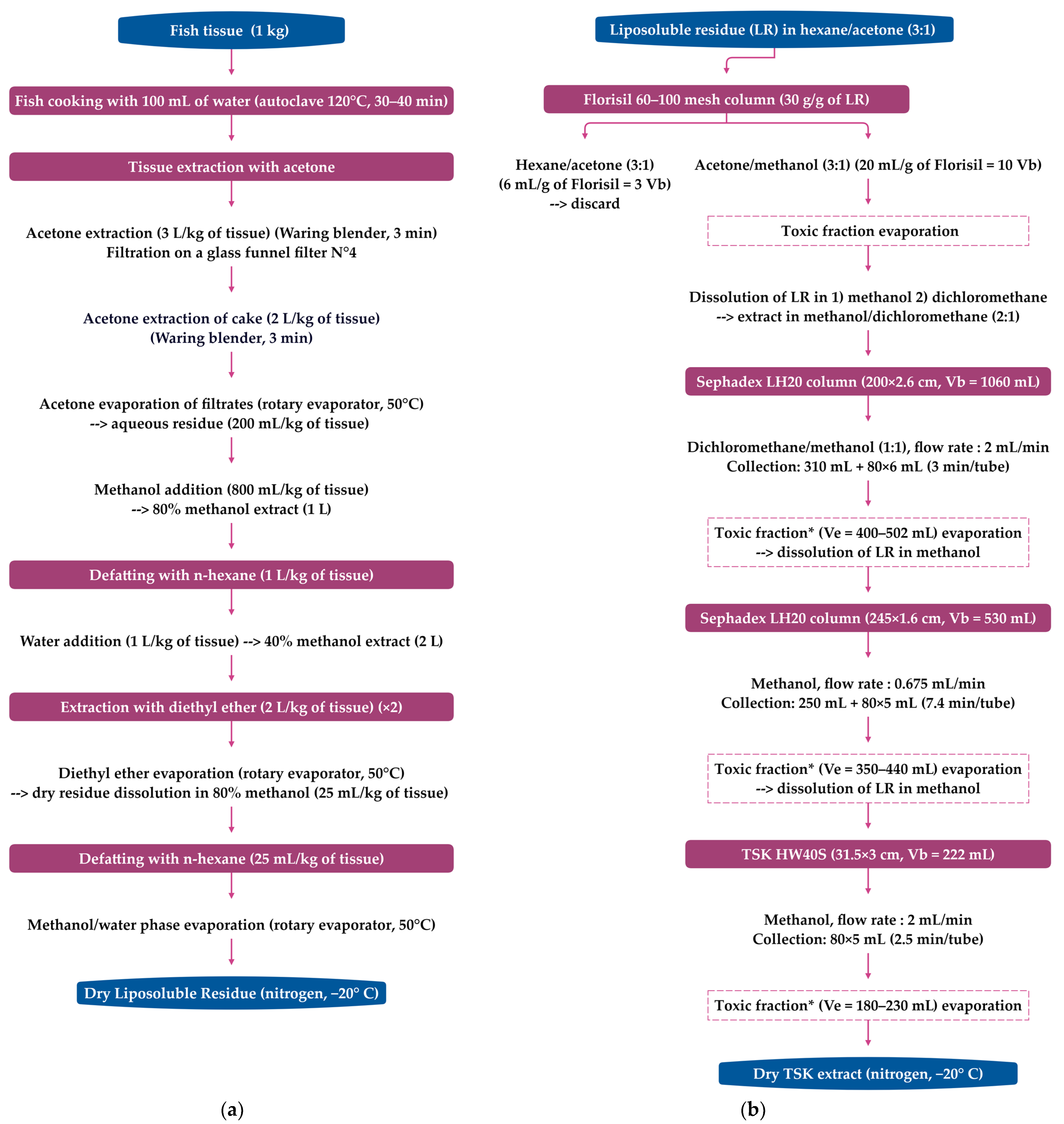
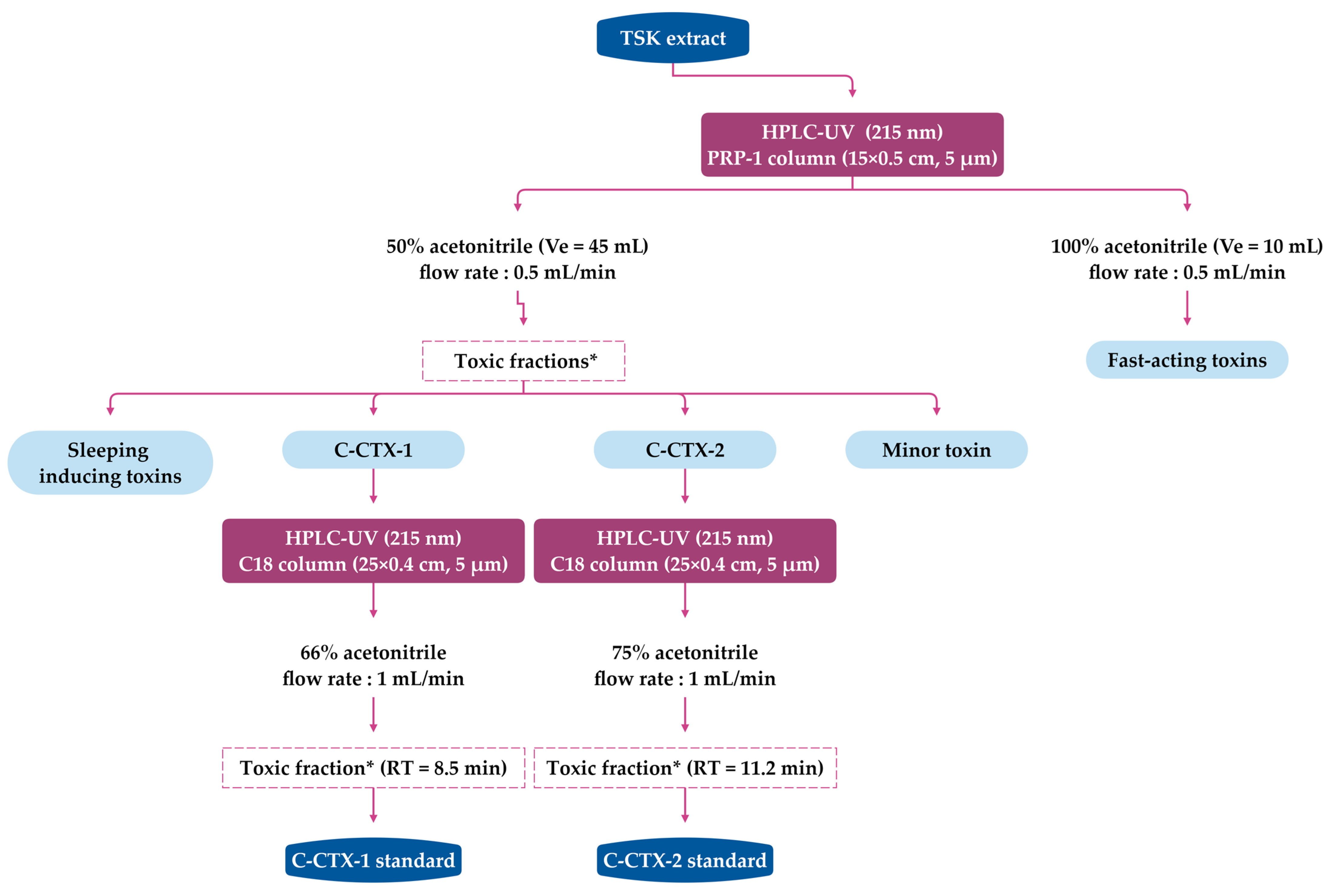
4. Extraction and Purification of C-CTXs
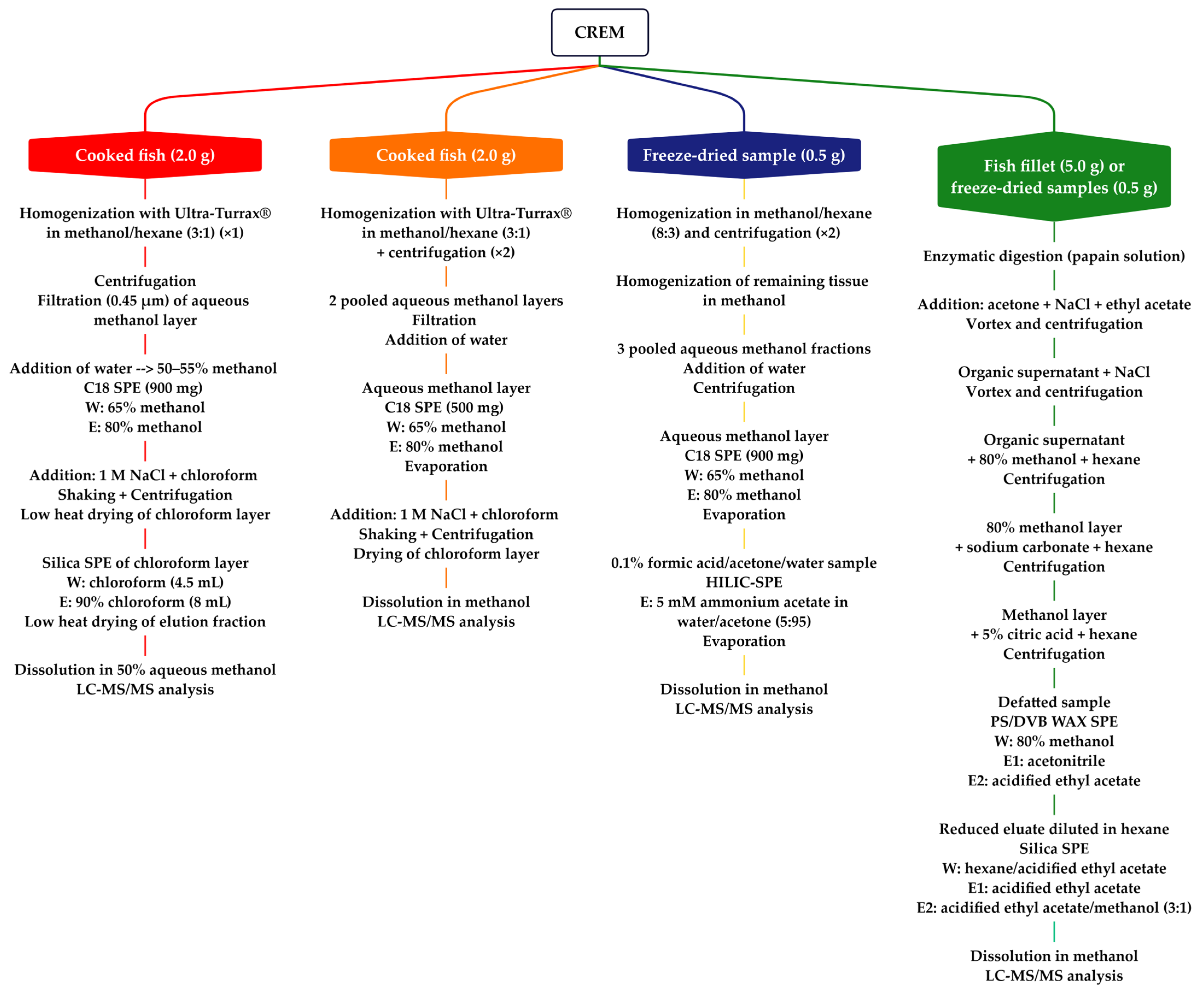
4.1. Extraction and Purification Methods for C-CTXs Isolation (>1 kg of Fish Flesh)
4.2. Rapid Extraction Methods for CTXs (<5 g of Fish Flesh)
- Reduce the quantity of fish flesh extracted (≤2 g).
- Replace the initial acetone extraction and filtration step with a one-step extraction and hexane cleanup.
- Reduce the number transfer and drying steps.
- Use centrifugation to speed the separation of phases.
- Consolidate LLE to a single step.
- Incorporate two orthogonal SPE cleanup steps on C18 and silica.
5. Analytical Methods for Detection, Identification, and Quantification of C-CTXs
5.1. Analysis of C-CTXs in Caribbean Fish Using HPLC-MS (Low-Resolution)
5.1.1. First Characterisation of C-CTX-1 and C-CTX-2 in Caribbean Fish
5.1.2. Combined Analysis of Toxin Profiles in Caribbean Fish Specimens (Horse-Eye Jacks, Grey Snapper, Grouper, Black Jack, and Barracuda) Using MBA, HPLC-MS, and RLB
5.2. Analysis of C-CTXs in Fish by HPLC-MS/MS (Low-Resolution)
5.2.1. First Quantification of C-CTX-1 in Caribbean Fish Using HPLC-MS/MS
5.2.2. Identification and Quantification of C-CTX-1 in Fish from Macaronesia Using HPLC-MS/MS with Water/Methanol LC Gradients and SRM on Precursor Ions [M+Na]+
5.2.3. Identification of C-CTX-1 and Identification of C-CTX Congeners in Fish Using HPLC-MS/MS with Water/Acetonitrile LC Gradients
- Identification of C-CTX-1 and C-CTXs congeners in Caribbean Fish
- Confirmation of C-CTX-1 and identification of C-CTX congeners in Fish from the Eastern Atlantic
5.2.4. Multianalytes Screening (P-CTXs, I-CTXs, and C-CTXs) Using HPLC-MS/MS with MRM on Precursor Ions [M+Na]+ and Confirmation Using MRM on Precursor Ions [M+NH4]+ (Methanol/Acetonitrile Elution)
5.3. Contribution of High-Resolution Mass Spectrometry for Identification and Chemical Characterisation of C-CTXs
5.3.1. Contribution of HRMS to Study Fish C-CTXs
- 17-hydroxy-C-CTX-1 ([M+H]+ m/z 1157.6255) with a possible location of the -OH group in the E-ring,
- two N-seco- forms of C-CTX-1 ([M+H]+ m/z 1143.6462) corresponding to C-CTX-1-reduced forms, previously known as C-CTX-3/-4 [120],
- 50,51-didehydro-C-CTX-3 ([M+H]+ m/z 1141.6306),
- 17-hydroxy-50,51-didehydro-C-CTX-3 ([M+H]+ m/z 1157.6255).
5.3.2. Contribution of HRMS to Study Gambierdiscus spp. Toxins
6. Discussion and Perspectives
6.1. Bioassays
6.2. Considerations for Analytical Approaches for C-CTX Detection and Quantification
6.3. Multiplicity of Caribbean Ciguatoxins
- Compounds with [M+H]+ at m/z 1143.6: isomers previously known as C-CTX-1143 showed no significant activity in radio-ligand binding assays [23,41]. However, RBA may not be sensitive enough to detect low amounts in fish, and another study identified an analogue, C-CTX-1143, using LC-MS/MS in a positive fraction by CBA-N2a [108]. Two anomers, C-CTX-3 and C-CTX-4 (C62H94O19), have been characterised using HRMS as the reduced forms of C-CTX-1 [120,131]. Their toxic potential and their role in fish total toxicity are yet to be clearly demonstrated due to their unstable hemiketal structure.
- Compounds with [M+H]+ at m/z 1157.6: two compounds with [M+H]+ at m/z 1157.6 were recently identified using LC-HRMS, 17-hydroxy-C-CTX-1 ([M+H]+ m/z 1157.6255) with a possible location of the -OH group in the E-ring, and putative 17-hydroxy-50,51-didehydro-C-CTX-3 ([M+H]+ m/z 1157.6255) [131]. An analogue C-CTX-1157 was also reported in a CTX-positive fraction analysed using CBA-N2a. [108] and in a positive fraction analysed using RBA (unpublished data from [40]).
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lewis, R.J.; Holmes, M.J. Origin and Transfer of Toxins Involved in Ciguatera. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1993, 106, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An Updated Review of Ciguatera Fish Poisoning: Clinical, Epidemiological, Environmental, and Public Health Management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pottier, I.; Vernoux, J.-P.; Lewis, R.J. Ciguatera Fish Poisoning in the Caribbean Islands and Western Atlantic. Rev. Environ. Contam. Toxicol. Contin. Residue Rev. 2001, 99–141. [Google Scholar] [CrossRef]
- Soliño, L.; Costa, P.R. Global Impact of Ciguatoxins and Ciguatera Fish Poisoning on Fish, Fisheries and Consumers. Environ. Res. 2020, 182, 109111. [Google Scholar] [CrossRef] [PubMed]
- Chinain, M.; Gatti, C.M.I.; Darius, H.T.; Quod, J.-P.; Tester, P.A. Ciguatera Poisonings: A Global Review of Occurrences and Trends. Harmful Algae 2021, 102, 101873. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, C.R.; Tartaglione, L.; Friedemann, M.; Spielmeyer, A.; Kappenstein, O.; Bodi, D. Ciguatera Mini Review: 21st Century Environmental Challenges and the Interdisciplinary Research Efforts Rising to Meet Them. Int. J. Environ. Res. Public. Health 2021, 18, 3027. [Google Scholar] [CrossRef] [PubMed]
- Dickey, R.W.; Plakas, S.M. Ciguatera: A Public Health Perspective. Toxicon 2010, 56, 123–136. [Google Scholar] [CrossRef]
- Darius, H.T.; Roué, M.; Sibat, M.; Viallon, J.; Gatti, C.M.I.; Vandersea, M.W.; Tester, P.A.; Litaker, R.W.; Amzil, Z.; Hess, P.; et al. Toxicological Investigations on the Sea Urchin Tripneustes Gratilla (Toxopneustidae, Echinoid) from Anaho Bay (Nuku Hiva, French Polynesia): Evidence for the Presence of Pacific Ciguatoxins. Mar. Drugs 2018, 16, 122. [Google Scholar] [CrossRef] [Green Version]
- Mak, Y.L.; Wai, T.-C.; Murphy, M.B.; Chan, W.H.; Wu, J.J.; Lam, J.C.W.; Chan, L.L.; Lam, P.K.S. Pacific Ciguatoxins in Food Web Components of Coral Reef Systems in the Republic of Kiribati. Environ. Sci. Technol. 2013, 47, 14070–14079. [Google Scholar] [CrossRef]
- Rongo, T.; van Woesik, R. Ciguatera Poisoning in Rarotonga, Southern Cook Islands. Harmful Algae 2011, 10, 345–355. [Google Scholar] [CrossRef]
- Roué, M.; Darius, H.T.; Picot, S.; Ung, A.; Viallon, J.; Gaertner-Mazouni, N.; Sibat, M.; Amzil, Z.; Chinain, M. Evidence of the Bioaccumulation of Ciguatoxins in Giant Clams (Tridacna Maxima) Exposed to Gambierdiscus spp. Cells. Harmful Algae 2016, 57, 78–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.; Rodriguez, I.; Barreiro, A.; Kaufmann, M.; Neto, A.I.; Hassouani, M.; Sabour, B.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. First Report of Ciguatoxins in Two Starfish Species: Ophidiaster Ophidianus and Marthasterias Glacialis. Toxins 2015, 7, 3740–3757. [Google Scholar] [CrossRef] [Green Version]
- Bagnis, R.; Mazelier, P.; Bennett, J.; Christian, E. Poissons de Polynésie, 3rd ed.; Les Editions du Pacifique: Papeete, Tahiti, 1973. [Google Scholar]
- Randall, J.E. Caribbean Reef Fishes, 3rd ed.; T.F.H. Publications, Inc. Ltd.: Hong Kong, China, 1996. [Google Scholar]
- Vernoux, J.P.; Magras, M.; Magras, P. Poissons Coralliens Des Antilles, 1st ed.; Faune & flore des Tropiques; Les Edition du Latanier: Gustavia, Saint Barthelémy, 1988; ISBN 2-9502284-2-9. Available online: https://www.researchgate.net/publication/301295899_''Poissons_coralliens_des_AntillesCoral_fishes_of_the_West_Indies_128_pages (accessed on 22 June 2023).
- Lehane, L.; Lewis, R.J. Ciguatera: Recent Advances but the Risk Remains. Int. J. Food Microbiol. 2000, 61, 91–125. [Google Scholar] [CrossRef]
- Mattei, C.; Vetter, I.; Eisenblätter, A.; Krock, B.; Ebbecke, M.; Desel, H.; Zimmermann, K. Ciguatera Fish Poisoning: A First Epidemic in Germany Highlights an Increasing Risk for European Countries. Toxicon 2014, 91, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis Malardé Institute French Polynesia Ciguawatch Initiative. Available online: https://ciguawatch.ilm.pf/fr/pacific-ciguatera-risk-map-and-declaration-system/ (accessed on 27 June 2023).
- FAO and WHO. Report of the Expert Meeting on Ciguatera Poisoning Rome, 19–23 November 2018; Food Safety and Quality, FAO and WHO: Rome, Italy, 2020; pp. 1–134. [Google Scholar]
- Vetter, I.; Touska, F.; Hess, A.; Hinsbey, R.; Sattler, S.; Lampert, A.; Sergejeva, M.; Sharov, A.; Collins, L.S.; Eberhardt, M.; et al. Ciguatoxins Activate Specific Cold Pain Pathways to Elicit Burning Pain from Cooling. EMBO J. 2012, 31, 3795–3808. [Google Scholar] [CrossRef] [Green Version]
- Epelboin, L.; Pérignon, A.; Hossen, V.; Vincent, R.; Krys, S.; Caumes, E. Two Clusters of Ciguatera Fish Poisoning in Paris, France, Related to Tropical Fish Imported From the French Caribbean by Travelers. J. Travel Med. 2014, 21, 397–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossen, V.; Soliño, L.; Leroy, P.; David, E.; Velge, P.; Dragacci, S.; Krys, S.; Flores Quintana, H.; Diogène, J. Contribution to the Risk Characterization of Ciguatoxins: LOAEL Estimated from Eight Ciguatera Fish Poisoning Events in Guadeloupe (French West Indies). Environ. Res. 2015, 143, 100–108. [Google Scholar] [CrossRef]
- Pottier, I.; Vernoux, J.P.; Jones, A.; Lewis, R.J. Analysis of Toxin Profiles in Three Different Fish Species Causing Ciguatera Fish Poisoning in Guadeloupe, French West Indies. Food Addit. Contam. 2002, 19, 1034–1042. [Google Scholar] [CrossRef]
- Gatti, C.M.I.; Chung, K.; Oehler, E.; Pierce, T.J.; Gribble, M.O.; Chinain, M. Screening for Predictors of Chronic Ciguatera Poisoning: An Exploratory Analysis among Hospitalized Cases from French Polynesia. Toxins 2021, 13, 646. [Google Scholar] [CrossRef]
- Boucaud-Maitre, D.; Vernoux, J.-P.; Pelczar, S.; Daudens-Vaysse, E.; Aubert, L.; Boa, S.; Ferracci, S.; Garnier, R. Incidence and Clinical Characteristics of Ciguatera Fish Poisoning in Guadeloupe (French West Indies) between 2013 and 2016: A Retrospective Cases-Series. Sci. Rep. 2018, 8, 3095. [Google Scholar] [CrossRef] [Green Version]
- Résière, D.; Florentin, J.; Mehdaoui, H.; Mahi, Z.; Gueye, P.; Hommel, D.; Pujo, J.; NKontcho, F.; Portecop, P.; Nevière, R.; et al. Clinical Characteristics of Ciguatera Poisoning in Martinique, French West Indies—A Case Series. Toxins 2022, 14, 535. [Google Scholar] [CrossRef]
- Quod, J.P.; Turquet, J. Ciguatera in Réunion Island (SW Indian Ocean): Epidemiology and Clinical Patterns. Toxicon 1996, 34, 779–785. [Google Scholar] [CrossRef]
- Lawrence, D.N.; Enriquez, M.B.; Lumish, R.M.; Maceo, A. Ciguatera Fish Poisoning in Miami. JAMA 1980, 244, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Oshiro, N.; Yogi, K.; Asato, S.; Sasaki, T.; Tamanaha, K.; Hirama, M.; Yasumoto, T.; Inafuku, Y. Ciguatera Incidence and Fish Toxicity in Okinawa, Japan. Toxicon 2010, 56, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Yogi, K.; Oshiro, N.; Inafuku, Y.; Hirama, M.; Yasumoto, T. Detailed LC-MS/MS Analysis of Ciguatoxins Revealing Distinct Regional and Species Characteristics in Fish and Causative Alga from the Pacific. Anal. Chem. 2011, 83, 8886–8891. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.V.; Uesugi, A.; Uchida, H.; Ky, P.X.; Minh, D.Q.; Watanabe, R.; Matsushima, R.; Oikawa, H.; Nagai, S.; Iwataki, M.; et al. Identification of Causative Ciguatoxins in Red Snappers Lutjanus Bohar Implicated in Ciguatera Fish Poisonings in Vietnam. Toxins 2018, 10, E420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, R.J.; Holmes, M.J.; Alewood, P.F.; Jones, A. Lonspray Mass Spectrometry of Ciguatoxin-1, Maitotoxin-2 and -3, and Related Marine Polyether Toxins. Nat. Toxins 1994, 2, 56–63. [Google Scholar] [CrossRef]
- Lewis, R.J.; Jones, A. Characterization of Ciguatoxins and Ciguatoxin Congeners Present in Ciguateric Fish by Gradient Reverse-Phase High-Performance Liquid Chromatography/Mass Spectrometry. Toxicon 1997, 35, 159–168. [Google Scholar] [CrossRef]
- Spielmeyer, A.; Loeffler, C.R.; Kappenstein, O. Identical Ciguatoxin-3C Group Profiles in Lutjanus Bohar from the Pacific and Indian Oceans - Indicating the Need to Re-Evaluate Geographical CTX Classifications. Front. Mar. Sci. 2022, 9, 937438. [Google Scholar] [CrossRef]
- Lewis, R.J.; Vernoux, J.-P.; Brereton, I.M. Structure of Caribbean Ciguatoxin Isolated from Caranx Latus. J. Am. Chem. Soc. 1998, 120, 5914–5920. [Google Scholar] [CrossRef]
- Vernoux, J.-P. Aspects Épidémiologiques, Toxicologiques et Préventifs de La Ciguatera Dans l’île de Saint-Barthélémy. Ph.D. Thesis, Université Bordeaux I, Bordeaux, France, 1986. [Google Scholar]
- Loeffler, C.R.; Robertson, A.; Flores Quintana, H.A.; Silander, M.C.; Smith, T.B.; Olsen, D. Ciguatoxin Prevalence in 4 Commercial Fish Species along an Oceanic Exposure Gradient in the US Virgin Islands. Environ. Toxicol. Chem. 2018, 37, 1852–1863. [Google Scholar] [CrossRef] [PubMed]
- Vernoux, J.-P.; Lewis, R.J. Isolation and Characterisation of Caribbean Ciguatoxins from the Horse-Eye Jack (Caranx Latus). Toxicon 1997, 35, 889–900. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.; Garcia, A.C.; Quintana, H.A.F.; Smith, T.B.; Ii, B.F.C.; Reale-Munroe, K.; Gulli, J.A.; Olsen, D.A.; Hooe-Rollman, J.I.; Jester, E.L.E.; et al. Invasive Lionfish (Pterois Volitans): A Potential Human Health Threat for Ciguatera Fish Poisoning in Tropical Waters. Mar. Drugs 2014, 12, 88–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pottier, I. La Ciguatera Aux Antilles: Épidémiologie, Analyse de La C-CTX-1 et Étude de La Diversité Des Ciguatoxines Dans Les Poissons Toxicophores. Ph.D. Thesis, Université de Caen, Caen, France, 2002. [Google Scholar]
- Pottier, I.; Hamilton, B.; Jones, A.; Lewis, R.J.; Vernoux, J.P. Identification of Slow and Fast-Acting Toxins in a Highly Ciguatoxic Barracuda (Sphyraena Barracuda) by HPLC/MS and Radiolabelled Ligand Binding. Toxicon 2003, 42, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Pottier, I.; Vernoux, J.-P.; Jones, A.; Lewis, R.J. Characterisation of Multiple Caribbean Ciguatoxins and Congeners in Individual Specimens of Horse-Eye Jack (Caranx Latus) by High-Performance Liquid Chromatography/Mass Spectrometry. Toxicon 2002, 40, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Mudge, E.M.; Miles, C.O.; Ivanova, L.; Uhlig, S.; James, K.S.; Erdner, D.L.; Fæste, C.K.; McCarron, P.; Robertson, A. Algal Ciguatoxin Identified as Source of Ciguatera Poisoning in the Caribbean. Chemosphere 2023, 330, 138659. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Vernoux, J.-P.; Jones, A.; Lewis, R.J. Isolation and Characterisation of Indian Ocean Ciguatoxin. Toxicon 2002, 40, 685–693. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Jones, A.; Lewis, R.J. Multiple Ciguatoxins Present in Indian Ocean Reef Fish. Toxicon 2002, 40, 1347–1353. [Google Scholar] [CrossRef]
- Diogène, J.; Reverté, L.; Rambla-Alegre, M.; del Río, V.; de la Iglesia, P.; Campàs, M.; Palacios, O.; Flores, C.; Caixach, J.; Ralijaona, C.; et al. Identification of Ciguatoxins in a Shark Involved in a Fatal Food Poisoning in the Indian Ocean. Sci. Rep. 2017, 7, 8240. [Google Scholar] [CrossRef] [Green Version]
- Vernoux, J.P.; Chinain, M.; Diogène, J.; Frémy, J.-M.; Hess, P.; Soria, M.; Trotereau, S.; Turquet, J. ANSES Opinion Collective Expert Appraisal Report on Contamination of Sharks, Especially Tiger and Bull Sharks, by Ciguatoxins: Occurrence, Analytical Methods, Human Cases Reported and Ethological Information (81 Pages). Available online: https://www.anses.fr/fr/system/files/ERCA2013sa0198EN.pdf (accessed on 15 April 2023).
- Vignaud, H. De Orbe Novo: The Eight Decades of Peter Martyr d’Anghera. Translated from the Latin; G. P. Putnam’s Sons: New York, NY, USA; London, UK, 1913. [Google Scholar]
- Wheeler, J.F.G.; Ommanney, F.D. Report on the Mauritius-Seychelles Fisheries Survey, 1948–1949, Etc; Colonial Office, H.M. Stationery Office: London, UK, 1953.
- Lewis, R.J. Ciguatera: Australian Perspectives on a Global Problem. Toxicon 2006, 48, 799–809. [Google Scholar] [CrossRef]
- Banner, A.H.; Scheuer, P.J.; Sasaki, S.; Helfrich, P.; Alender, C.B. Observations on Ciguatera-Type Toxin in Fish. Ann. N. Y. Acad. Sci. 1960, 90, 770–787. [Google Scholar] [CrossRef] [PubMed]
- Banner, A.H.; Helfrich, P. The Distribution of Ciguatera in the Tropical Pacific; Hawaii Marine Laboratory, University of Hawaii: Honolulu, HI, USA, 1964; pp. 1–48. [Google Scholar]
- Hashimoto, Y.; Konosu, S.; Yasumoto, T.; Kamiya, H. Ciguatera in the Ryukyu and Amami Islands. Nippon. Suisan Gakkaishi 1969, 35, 316–326. [Google Scholar] [CrossRef] [Green Version]
- Yasumoto, T.; Scheuer, P.J. Marine Toxins of the Pacific. 8. Ciguatoxin from Moray Eel Livers. Toxicon Off. J. Int. Soc. Toxinol. 1969, 7, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Randall, J.E. A Review of Ciguatera, Tropical Fish Poisoning, with a Tentative Explanation of Its Cause. Bull. Mar. Sci. 1958, 8, 236–267. [Google Scholar]
- Randall, J.E. Food Habits of Reef Fishes of the West Indies. In Proceedings of the Studies in Tropical Oceanography, Miami Beach, FL, USA, 17–24 November 1965; University of Miami, Institute of Marine Sciences: Coral Gables, FL, USA, 1967; Volume 5, pp. 665–847. [Google Scholar]
- Yasumoto, T.; Hashimoto, Y.; Bagnis, R.; Randall, J.E.; Banner, A.H. Toxicity of the Surgeonfishes. Bull. Jpn. Soc. Sci. Fish. 1971, 37, 724–734. [Google Scholar] [CrossRef]
- Bagnis, R.; Vernoux, J.P. Ciguatoxin and edible reef fishes. Bull. Soc. Pathol. Exot. Filiales 1975, 68, 320–325. [Google Scholar]
- Vernoux, J.P.; Bagnis, R. Fractionnement d’extraits lipidiques ciguatoxiques en milieu alcalin. Biochimie 1976, 58, 479–484. [Google Scholar] [CrossRef]
- Chanteau, S.; Bagnis, R.; Yasumoto, T. Purification de la ciguatoxine de la loche Epinephelus microdon (Bleecker). Biochimie 1976, 58, 1149–1151. [Google Scholar] [CrossRef]
- Chungue, E.; Bagnis, R.; Fusetani, N.; Hashimoto, Y. Isolation of Two Toxins from a Parrotfish Scarus Gibbus. Toxicon 1977, 15, 89–93. [Google Scholar] [CrossRef]
- Vernoux, J.P. L’ichtyosarcotoxisme de Type Ciguatera Aux Antilles et En Polynésie Française: Test de Ciguatoxicité et Chaîne Trophique Ciguatérigène. Ph.D. Thesis, Univeristé Bordeaux I, Bordeaux, France, 1981. [Google Scholar]
- Bagnis, R.; Chanteau, S.; Yasumoto, T. Significance of the various toxins present on dead coral substrata in the origin of ciguatera. Bull. Soc. Pathol. Exot. Filiales 1977, 70, 320–324. [Google Scholar]
- Yasumoto, T.; Nakajima, I.; Bagnis, R.; Adachi, R. Finding of a Dinoflagellate as a Likely Culprit of Ciguatera. Bull. Jpn. Soc. Sci. Fish. 1977, 43, 1021–1026. [Google Scholar] [CrossRef] [Green Version]
- Adachi, R.; Fukuyo, Y. The Thecal Structure of a Marine Toxic Dinoflagellate Gambierdiscus Toxicus Gen. et Sp. Nov. Collected in a Ciguatera-Endemic Area. Bull. Jpn. Soc. Sci. Fish. 1979, 45, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Bagnis, R.; Chanteau, S.; Chungue, E.; Hurtel, J.M.; Yasumoto, T.; Inoue, A. Origins of Ciguatera Fish Poisoning: A New Dinoflagellate, Gambierdiscus Toxicus Adachi and Fukuyo, Definitively Involved as a Causal Agent. Toxicon 1980, 18, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Bagnis, R.; P.Vernoux, J. Toxicity of the Surgeonfishes-II. Bull. Jpn. Soc. Sci. Fish. 1976, 42, 359–365. [Google Scholar] [CrossRef]
- Vernoux, J.-P.; Lejeune, J. Ciguatera in the French West Indies. In Memoirs of the Queensland Museum; Queensland Museum: Brisbane, Australia, 1994; Volume 34, pp. 631–638. [Google Scholar]
- Murata, M.; Naoki, H.; Iwashita, T.; Matsunaga, S.; Sasaki, M.; Yokoyama, A.; Yasumoto, T. Structure of Maitotoxin. J. Am. Chem. Soc. 1993, 115, 2060–2062. [Google Scholar] [CrossRef]
- Zheng, W.; DeMattei, J.A.; Wu, J.-P.; Duan, J.J.-W.; Cook, L.R.; Oinuma, H.; Kishi, Y. Complete Relative Stereochemistry of Maitotoxin. J. Am. Chem. Soc. 1996, 118, 7946–7968. [Google Scholar] [CrossRef]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Nau, A.W.; Holland, W.C.; Chinain, M.; Holmes, M.J.; Tester, P.A. Global Distribution of Ciguatera Causing Dinoflagellates in the Genus Gambierdiscus. Toxicon 2010, 56, 711–730. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J. Purification and Characterisation of Large and Small Maitotoxins from Cultured Gambierdiscus Toxicus. Nat. Toxins 1994, 2, 64–72. [Google Scholar] [CrossRef]
- Murray, J.S.; Finch, S.C.; Mudge, E.M.; Wilkins, A.L.; Puddick, J.; Harwood, D.T.; Rhodes, L.L.; van Ginkel, R.; Rise, F.; Prinsep, M.R. Structural Characterization of Maitotoxins Produced by Toxic Gambierdiscus Species. Mar. Drugs 2022, 20, 453. [Google Scholar] [CrossRef]
- Wang, D.-Z.; Xin, Y.-H.; Wang, M.-H. Gambierdiscus and Its Associated Toxins: A Minireview. Toxins 2022, 14, 485. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J. Origin of Ciguateric Fish: Quantitative Modelling of the Flow of Ciguatoxin through a Marine Food Chain. Toxins 2022, 14, 534. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J.; Gillespie, N.C. Toxicity of Australian and French Polynesian Strains of Gambierdiscus Toxicus (Dinophyceae) Grown in Culture: Characterization of a New Type of Maitotoxin. Toxicon Off. J. Int. Soc. Toxinol. 1990, 28, 1159–1172. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.J.; Lewis, R.J.; Poli, M.A.; Gillespie, N.C. Strain Dependent Production of Ciguatoxin Precursors (Gambiertoxins) by Gambierdiscus Toxicus (Dinophyceae) in Culture. Toxicon Off. J. Int. Soc. Toxinol. 1991, 29, 761–775. [Google Scholar] [CrossRef] [PubMed]
- Sakami, T.; Nakahara, H.; Chinain, M.; Ishida, Y. Effects of Epiphytic Bacteria on the Growth of the Toxic Dinoflagellate Gambierdiscus Toxicus (Dinophyceae). J. Exp. Mar. Biol. Ecol. 1999, 233, 231–246. [Google Scholar] [CrossRef]
- Chinain, M.; Darius, H.T.; Ung, A.; Cruchet, P.; Wang, Z.; Ponton, D.; Laurent, D.; Pauillac, S. Growth and Toxin Production in the Ciguatera-Causing Dinoflagellate Gambierdiscus Polynesiensis (Dinophyceae) in Culture. Toxicon 2010, 56, 739–750. [Google Scholar] [CrossRef]
- Gillespie, N.C.; Lewis, R.J.; Burke, J.; Holmes, M.J. The Significance of the Absence of Ciguatoxin in a Wild Population of a Gambierdiscus Toxicus. In Proceedings of the Fifth International Coral Reef Congress, Tahiti, French Polynesia, 27 May–1 June 1985; Delsalle, B., Galzin, R., Salvat, B., Eds.; Antenne Museum-Ephe: Tahiti, French Polynesia, 1985; Volume 4, pp. 437–442. [Google Scholar]
- Lewis, R.J.; Inserra, M.; Vetter, I.; Holland, W.C.; Hardison, D.R.; Tester, P.A.; Litaker, R.W. Rapid Extraction and Identification of Maitotoxin and Ciguatoxin-Like Toxins from Caribbean and Pacific Gambierdiscus Using a New Functional Bioassay. PLoS ONE 2016, 11, e0160006. [Google Scholar] [CrossRef] [Green Version]
- Longo, S.; Sibat, M.; Viallon, J.; Darius, H.T.; Hess, P.; Chinain, M. Intraspecific Variability in the Toxin Production and Toxin Profiles of In Vitro Cultures of Gambierdiscus Polynesiensis (Dinophyceae) from French Polynesia. Toxins 2019, 11, 735. [Google Scholar] [CrossRef] [Green Version]
- Darius, H.T.; Revel, T.; Viallon, J.; Sibat, M.; Cruchet, P.; Longo, S.; Hardison, D.R.; Holland, W.C.; Tester, P.A.; Litaker, R.W.; et al. Comparative Study on the Performance of Three Detection Methods for the Quantification of Pacific Ciguatoxins in French Polynesian Strains of Gambierdiscus Polynesiensis. Mar. Drugs 2022, 20, 348. [Google Scholar] [CrossRef]
- Stuart, J.; Smith, K.F.; Rhodes, L.; Murray, J.S.; Viallon, J.; Henry, K.; Darius, H.T.; Murray, S.A.; De Azevedo, C.D.; Argyle, P.; et al. Geographical Distribution, Molecular and Toxin Diversity of the Dinoflagellate Species Gambierdiscus Honu in the Pacific Region. Harmful Algae 2022, 118, 102308. [Google Scholar] [CrossRef]
- Chinain, M.; Faust, M.A.; Pauillac, S. Morphology and Molecular Analyses of Three Toxic Species of Gambierdiscus (Dinophyceae): G. Pacificus, Sp. Nov., G. Australes, Sp. Nov., and G. Polynesiensis, Sp. Nov. J. Phycol. 1999, 35, 1282–1296. [Google Scholar] [CrossRef]
- Yon, T.; Sibat, M.; Réveillon, D.; Bertrand, S.; Chinain, M.; Hess, P. Deeper Insight into Gambierdiscus Polynesiensis Toxin Production Relies on Specific Optimization of High-Performance Liquid Chromatography-High Resolution Mass Spectrometry. Talanta 2021, 232, 122400. [Google Scholar] [CrossRef] [PubMed]
- Kibler, S.R.; Davenport, E.D.; Tester, P.A.; Hardison, D.R.; Holland, W.C.; Litaker, R.W. Gambierdiscus and Fukuyoa Species in the Greater Caribbean: Regional Growth Projections for Ciguatera-Associated Dinoflagellates. Ecol. Model. 2017, 360, 204–218. [Google Scholar] [CrossRef]
- Laza-Martínez, A.; David, H.; Riobó, P.; Miguel, I.; Orive, E. Characterization of a Strain of Fukuyoa Paulensis (Dinophyceae) from the Western Mediterranean Sea. J. Eukaryot. Microbiol. 2016, 63, 481–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, M.J.; Venables, B.; Lewis, R.J. Critical Review and Conceptual and Quantitative Models for the Transfer and Depuration of Ciguatoxins in Fishes. Toxins 2021, 13, 515. [Google Scholar] [CrossRef]
- Ikehara, T.; Kuniyoshi, K.; Oshiro, N.; Yasumoto, T. Biooxidation of Ciguatoxins Leads to Species-Specific Toxin Profiles. Toxins 2017, 9, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vernoux, J.P.; Lahlou, N.; Abbad el Andaloussi, S.; Riyeche, N.; Magras, L.P. A Study of the Distribution of Ciguatoxin in Individual Caribbean Fish. Acta Trop. 1985, 42, 225–233. [Google Scholar]
- Gillespie, N.C.; Lewis, R.J.; Pearn, J.H.; Bourke, A.T.; Holmes, M.J.; Bourke, J.B.; Shields, W.J. Ciguatera in Australia. Occurrence, Clinical Features, Pathophysiology and Management. Med. J. Aust. 1986, 145, 584–590. [Google Scholar] [CrossRef]
- Crump, J.A.; McLay, C.L.; Chambers, S.T. Ciguatera Fish Poisoning. Postgrad. Med. J. 1999, 75, 678–679. [Google Scholar] [CrossRef]
- Vernoux, J.-P. La Ciguatera Dans l’île de Saint-Barthélémy: Aspects Épidémiologiques, Toxicologiques et Préventifs. Oceanol. Acta 1988, 11, 37–46. [Google Scholar]
- Clausing, R.J.; Losen, B.; Oberhaensli, F.R.; Darius, H.T.; Sibat, M.; Hess, P.; Swarzenski, P.W.; Chinain, M.; Dechraoui Bottein, M.-Y. Experimental Evidence of Dietary Ciguatoxin Accumulation in an Herbivorous Coral Reef Fish. Aquat. Toxicol. 2018, 200, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Inserra, M.C.; Israel, M.R.; Caldwell, A.; Castro, J.; Deuis, J.R.; Harrington, A.M.; Keramidas, A.; Garcia-Caraballo, S.; Maddern, J.; Erickson, A.; et al. Multiple Sodium Channel Isoforms Mediate the Pathological Effects of Pacific Ciguatoxin-1. Sci. Rep. 2017, 7, 42810. [Google Scholar] [CrossRef] [Green Version]
- Inserra, M.; Lavrukhina, Y.; Jones, A.; Lewis, R.J.; Vetter, I. Ciguatoxin Detection Methods and High-Throughput Assays. In Analysis of Food Toxins and Toxicants; Wong, Y., Lewis, R.J., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 469–488. ISBN 978-1-118-99268-5. [Google Scholar]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Fukui, M.; Yasumoto, T. Structures and Configurations of Ciguatoxin from the Moray Eel Gymnothorax Javanicus and Its Likely Precursor from the Dinoflagellate Gambierdiscus Toxicus. J. Am. Chem. Soc. 1990, 112, 4380–4386. [Google Scholar] [CrossRef]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Yasumoto, T. Structures of Ciguatoxin and Its Congener. J. Am. Chem. Soc. 1989, 111, 8929–8931. [Google Scholar] [CrossRef]
- Satake, M.; Morohashi, A.; Oguri, H.; Oishi, T.; Hirama, M.; Harada, N.; Yasumoto, T. The Absolute Configuration of Ciguatoxin. J. Am. Chem. Soc. 1997, 119, 11325–11326. [Google Scholar] [CrossRef]
- Dickey, R.W.; Granade, H.R.; Bencsath, F.A.; Martin, G.E. Characterization of Polyether Biotoxins from Caribbean Barracuda (Sphyraena Barracuda) and Horse Eye Jack (Caranx Latus). In Proceedings of the International Symposium on Ciguatera and Marine Natural Product, Honolulu, HI, USA, 8–10 August 1994; Hokama, Y., Scheuer, P.J., Yasumoto, T., Eds.; Asian Pacific Research Foundation: Honolulu, HI, USA, 1995; p. 292. [Google Scholar]
- Crouch, R.C.; Martin, G.E.; Musser, S.M.; Ray Grenade, H.; Dickey, R.W. Improvements in the Sensitivity of Inverse-Detected Heteronuclear Correlation Spectra Using Micro Inverse Probes and Micro Cells: HMQC and HMBC Spectra of Caribbean Ciguatoxin — Preliminary Structural Inferences. Tetrahedron Lett. 1995, 36, 6827–6830. [Google Scholar] [CrossRef]
- Vernoux, J.-P.; Abbad el Andaloussi, S. Hétérogénéité des ciguatoxines extraites de poissons péchés aux Antilles françaises. Biochimie 1986, 68, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Vernoux, J.P.; Talha, F. Fractionation and Purification of Some Muscular and Visceral Ciguatoxins Extracted from Carribean Fish. Comp. Biochem. Physiol. B 1989, 94, 499–504. [Google Scholar] [CrossRef]
- Pérez-Arellano, J.-L.; Luzardo, O.P.; Brito, A.P.; Cabrera, M.H.; Zumbado, M.; Carranza, C.; Angel-Moreno, A.; Dickey, R.W.; Boada, L.D. Ciguatera Fish Poisoning, Canary Islands. Emerg. Infect. Dis. J. -CDC 2005, 11, 1981–1982. [Google Scholar] [CrossRef] [Green Version]
- Boada, L.D.; Zumbado, M.; Luzardo, O.P.; Almeida-González, M.; Plakas, S.M.; Granade, H.R.; Abraham, A.; Jester, E.L.E.; Dickey, R.W. Ciguatera Fish Poisoning on the West Africa Coast: An Emerging Risk in the Canary Islands (Spain). Toxicon 2010, 56, 1516–1519. [Google Scholar] [CrossRef]
- Otero, P.; Pérez, S.; Alfonso, A.; Vale, C.; Rodríguez, P.; Gouveia, N.N.; Gouveia, N.; Delgado, J.; Vale, P.; Hirama, M.; et al. First Toxin Profile of Ciguateric Fish in Madeira Arquipelago (Europe). Anal. Chem. 2010, 82, 6032–6039. [Google Scholar] [CrossRef]
- Abraham, A.; Jester, E.L.E.; Granade, H.R.; Plakas, S.M.; Dickey, R.W. Caribbean Ciguatoxin Profile in Raw and Cooked Fish Implicated in Ciguatera. Food Chem. 2012, 131, 192–198. [Google Scholar] [CrossRef]
- Costa, P.R.; Estevez, P.; Castro, D.; Soliño, L.; Gouveia, N.; Santos, C.; Rodrigues, S.M.; Leao, J.M.; Gago-Martínez, A. New Insights into the Occurrence and Toxin Profile of Ciguatoxins in Selvagens Islands (Madeira, Portugal). Toxins 2018, 10, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estevez, P.; Castro, D.; Manuel Leao, J.; Yasumoto, T.; Dickey, R.; Gago-Martinez, A. Implementation of Liquid Chromatography Tandem Mass Spectrometry for the Analysis of Ciguatera Fish Poisoning in Contaminated Fish Samples from Atlantic Coasts. Food Chem. 2019, 280, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Henao, J.A.; García-Álvarez, N.; Fernández, A.; Saavedra, P.; Silva Sergent, F.; Padilla, D.; Acosta-Hernández, B.; Martel Suárez, M.; Diogène, J.; Real, F. Predictive Score and Probability of CTX-like Toxicity in Fish Samples from the Official Control of Ciguatera in the Canary Islands. Sci. Total Environ. 2019, 673, 576–584. [Google Scholar] [CrossRef]
- Ramos-Sosa, M.J.; García-Álvarez, N.; Sanchez-Henao, A.; Silva Sergent, F.; Padilla, D.; Estévez, P.; Caballero, M.J.; Martín-Barrasa, J.L.; Gago-Martínez, A.; Diogène, J.; et al. Ciguatoxin Detection in Flesh and Liver of Relevant Fish Species from the Canary Islands. Toxins 2022, 14, 46. [Google Scholar] [CrossRef] [PubMed]
- Tudó, À.; Rambla-Alegre, M.; Flores, C.; Sagristà, N.; Aguayo, P.; Reverté, L.; Campàs, M.; Gouveia, N.; Santos, C.; Andree, K.B.; et al. Identification of New CTX Analogues in Fish from the Madeira and Selvagens Archipelagos by Neuro-2a CBA and LC-HRMS. Mar. Drugs 2022, 20, 236. [Google Scholar] [CrossRef]
- Otero, P.; Silva, M. Emerging Marine Biotoxins in European Waters: Potential Risks and Analytical Challenges. Mar. Drugs 2022, 20, 199. [Google Scholar] [CrossRef]
- Estevez, P.; Castro, D.; Pequeño-Valtierra, A.; Giraldez, J.; Gago-Martinez, A. Emerging Marine Biotoxins in Seafood from European Coasts: Incidence and Analytical Challenges. Foods 2019, 8, 149. [Google Scholar] [CrossRef] [Green Version]
- Tester, P.A.; Feldman, R.L.; Nau, A.W.; Kibler, S.R.; Wayne Litaker, R. Ciguatera Fish Poisoning and Sea Surface Temperatures in the Caribbean Sea and the West Indies. Toxicon 2010, 56, 698–710. [Google Scholar] [CrossRef]
- Tester, P.A.; Litaker, R.W.; Berdalet, E. Climate Change and Harmful Benthic Microalgae. Harmful Algae 2020, 91, 101655. [Google Scholar] [CrossRef]
- Pasinszki, T.; Lako, J.; Dennis, T.E. Advances in Detecting Ciguatoxins in Fish. Toxins 2020, 12, 494. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Igarashi, T.; Legrand, A.-M.; Cruchet, P.; Chinain, M.; Fujita, T.; Naoki, H. Structural Elucidation of Ciguatoxin Congeners by Fast-Atom Bombardment Tandem Mass Spectroscopy. J. Am. Chem. Soc. 2000, 122, 4988–4989. [Google Scholar] [CrossRef]
- Kryuchkov, F.; Robertson, A.; Miles, C.O.; Mudge, E.M.; Uhlig, S. LC-HRMS and Chemical Derivatization Strategies for the Structure Elucidation of Caribbean Ciguatoxins: Identification of C-CTX-3 and -4. Mar. Drugs 2020, 18, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estevez, P.; Leao, J.M.; Yasumoto, T.; Dickey, R.W.; Gago-Martinez, A. Caribbean Ciguatoxin-1 Stability under Strongly Acidic Conditions: Characterisation of a New C-CTX1 Methoxy Congener. Food Addit. Contam. Part A 2020, 37, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Sellin, M.; Poli, M.A.; Norton, R.S.; MacLeod, J.K.; Sheil, M.M. Purification and Characterization of Ciguatoxins from Moray Eel (Lycodontis Javanicus, Muraenidae). Toxicon 1991, 29, 1115–1127. [Google Scholar] [CrossRef]
- Satake, M.; Murata, M.; Yasumoto, T. The Structure of CTX3C, a Ciguatoxin Congener Isolated from Cultured Gambierdiscus Toxicus. Tetrahedron Lett. 1993, 34, 1975–1978. [Google Scholar] [CrossRef]
- Lin, Y.-Y.; Risk, M.; Ray, S.M.; Van Engen, D.; Clardy, J.; Golik, J.; James, J.C.; Nakanishi, K. Isolation and Structure of Brevetoxin B from the “Red Tide” Dinoflagellate Ptychodiscus Brevis (Gymnodinium Breve). J. Am. Chem. Soc. 1981, 103, 6773–6775. [Google Scholar] [CrossRef]
- Lewis, R.J.; Norton, R.S.; Brereton, I.M.; Eccles, C.D. Ciguatoxin-2 Is a Diastereomer of Ciguatoxin-3. Toxicon 1993, 31, 637–643. [Google Scholar] [CrossRef]
- Satake, M.; Ishibashi, Y.; Legrand, A.-M.; Yasumoto, T. Isolation and Structure of Ciguatoxin-4A, a New Ciguatoxin Precursor, from Cultures of Dinoflagellate Gambierdiscus Toxicus and Parrotfish Scarus Gibbus. Biosci. Biotechnol. Biochem. 1997, 60, 2103–2105. [Google Scholar] [CrossRef] [Green Version]
- Legrand, A.M.; Litaudon, M.; Genthon, J.N.; Bagnis, R.; Yasumoto, T. Isolation and Some Properties of Ciguatoxin. J. Appl. Phycol. 1989, 1, 183–188. [Google Scholar] [CrossRef]
- Yasumoto, T. The Chemistry and Biological Function of Natural Marine Toxins. Chem. Rec. 2001, 1, 228–242. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Yasumoto, T. Quantification of Representative Ciguatoxins in the Pacific Using Quantitative Nuclear Magnetic Resonance Spectroscopy. Mar. Drugs 2017, 15, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, D.; Estévez, P.; Leao-Martins, J.M.; Dickey, R.W.; García-Álvarez, N.; Real, F.; Costa, P.R.; Gago-Martínez, A. Preparation of Ciguatoxin Reference Materials from Canary Islands (Spain) and Madeira Archipelago (Portugal) Fish. J. Mar. Sci. Eng. 2022, 10, 835. [Google Scholar] [CrossRef]
- Estevez, P.; Oses Prieto, J.; Burlingame, A.; Gago Martinez, A. Characterization of the Ciguatoxin Profile in Fish Samples from the Eastern Atlantic Ocean Using Capillary Liquid Chromatography-High Resolution Mass Spectrometry. Food Chem. 2023, 418, 135960. [Google Scholar] [CrossRef]
- Mudge, E.M.; Meija, J.; Uhlig, S.; Robertson, A.; McCarron, P.; Miles, C.O. Production and Stability of Oxygen-18 Labeled Caribbean Ciguatoxins and Gambierones. Toxicon 2022, 211, 11–20. [Google Scholar] [CrossRef]
- Hashimoto, Y. A Note on the Poison of a Barracuda, Sphyraena Picuda BLOCH & SCHNEIDER. Bull. Jpn. Soc. Sci. Fish. 1956, 21, 1153–1157. [Google Scholar]
- Bertuzzi, T.; Rastelli, S.; Mulazzi, A.; Pietri, A. Evaluation and Improvement of Extraction Methods for the Analysis of Aflatoxins B1, B2, G1 and G2 from Naturally Contaminated Maize. Food Anal. Methods 2012, 5, 512–519. [Google Scholar] [CrossRef]
- Vaz, A.; Cabral Silva, A.C.; Rodrigues, P.; Venâncio, A. Detection Methods for Aflatoxin M1 in Dairy Products. Microorganisms 2020, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Scheuer, P.J. Marine Toxins. Acc. Chem. Res. 1977, 10, 33–39. [Google Scholar] [CrossRef]
- Lewis, R.J.; Hoy, A.W.; Sellin, M. Ciguatera and Mannitol: In Vivo and in Vitro Assessment in Mice. Toxicon Off. J. Int. Soc. Toxinol. 1993, 31, 1039–1050. [Google Scholar] [CrossRef]
- Bidard, J.N.; Vijverberg, H.P.; Frelin, C.; Chungue, E.; Legrand, A.M.; Bagnis, R.; Lazdunski, M. Ciguatoxin Is a Novel Type of Na+ Channel Toxin. J. Biol. Chem. 1984, 259, 8353–8357. [Google Scholar] [CrossRef]
- Nicholson, G.M.; Lewis, R.J. Ciguatoxins: Cyclic Polyether Modulators of Voltage-Gated Iion Channel Function. Mar. Drugs 2006, 4, 82–118. [Google Scholar] [CrossRef] [Green Version]
- Sauviat, M.-P.; Marquais, M.; Vernoux, J.-P. Muscarinic Effects of the Caribbean Ciguatoxin C-CTX-1 on Frog Atrial Heart Muscle. Toxicon 2002, 40, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Sauviat, M.-P.; Boydron-Le Garrec, R.; Masson, J.-B.; Lewis, R.L.; Vernoux, J.-P.; Molgó, J.; Laurent, D.; Benoit, E. Mechanisms Involved in the Swelling of Erythrocytes Caused by Pacific and Caribbean Ciguatoxins. Blood Cells. Mol. Dis. 2006, 36, 1–9. [Google Scholar] [CrossRef]
- Mattei, C.; Marquais, M.; Schlumberger, S.; Molgó, J.; Vernoux, J.-P.; Lewis, R.J.; Benoit, E. Analysis of Caribbean Ciguatoxin-1 Effects on Frog Myelinated Axons and the Neuromuscular Junction. Toxicon 2010, 56, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Hokama, Y.; Dickey, R.W.; Granade, H.R.; Lewis, R.; Yasumoto, T.; Wekell, M.M. Detection of Sodium Channel Toxins: Directed Cytotoxicity Assays of Purified Ciguatoxins, Brevetoxins, Saxitoxins, and Seafood Extracts. J. AOAC Int. 1995, 78, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Dechraoui, M.-Y.; Naar, J.; Pauillac, S.; Legrand, A.-M. Ciguatoxins and Brevetoxins, Neurotoxic Polyether Compounds Active on Sodium Channels. Toxicon 1999, 37, 125–143. [Google Scholar] [CrossRef]
- Asthana, P.; Zhang, N.; Kumar, G.; Chine, V.B.; Singh, K.K.; Mak, Y.L.; Chan, L.L.; Lam, P.K.S.; Ma, C.H.E. Pacific Ciguatoxin Induces Excitotoxicity and Neurodegeneration in the Motor Cortex Via Caspase 3 Activation: Implication for Irreversible Motor Deficit. Mol. Neurobiol. 2018, 55, 6769–6787. [Google Scholar] [CrossRef]
- L’Herondelle, K.; Talagas, M.; Mignen, O.; Misery, L.; Le Garrec, R. Neurological Disturbances of Ciguatera Poisoning: Clinical Features and Pathophysiological Basis. Cells 2020, 9, 2291. [Google Scholar] [CrossRef]
- Pearn, J. Neurology of Ciguatera. J. Neurol. Neurosurg. Psychiatry 2001, 70, 4–8. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M.A.; Arena, P.; Levin, B.; Fleming, L.; Fernandez, M.; Weisman, R.; Bernstein, J.; Schrank, K.; Blythe, D.; Backer, L.; et al. Neuropsychological Study of Ciguatera Fish Poisoning: A Longitudinal Case-Control Study. Arch. Clin. Neuropsychol. 2007, 22, 545–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raposo-García, S.; Castro, D.; Lence, E.; Estévez, P.; Leão, J.M.; González-Bello, C.; Gago-Martínez, A.; Louzao, M.C.; Vale, C.; Botana, L.M. In Silico Simulations and Functional Cell Studies Evidence Similar Potency and Distinct Binding of Pacific and Caribbean Ciguatoxins. Expo. Health 2022. [Google Scholar] [CrossRef]
- Lahlou, N. Distribution cellulaire et subcellulaire de la ciguatoxine chez le poisson et action toxicologique chez le poussin. Ph.D. Thesis, Bordeaux I, Bordeaux, France, 1985. [Google Scholar]
- Bennett, C.T.; Robertson, A. Depuration Kinetics and Growth Dilution of Caribbean Ciguatoxin in the Omnivore Lagodon Rhomboides: Implications for Trophic Transfer and Ciguatera Risk. Toxins 2021, 13, 774. [Google Scholar] [CrossRef]
- Leite, I.d.P.; Sdiri, K.; Taylor, A.; Viallon, J.; Gharbia, H.B.; Mafra Júnior, L.L.; Swarzenski, P.; Oberhaensli, F.; Darius, H.T.; Chinain, M.; et al. Experimental Evidence of Ciguatoxin Accumulation and Depuration in Carnivorous Lionfish. Toxins 2021, 13, 564. [Google Scholar] [CrossRef]
- Li, J.; Mak, Y.L.; Chang, Y.-H.; Xiao, C.; Chen, Y.-M.; Shen, J.; Wang, Q.; Ruan, Y.; Lam, P.K.S. Uptake and Depuration Kinetics of Pacific Ciguatoxins in Orange-Spotted Grouper (Epinephelus Coioides). Environ. Sci. Technol. 2020, 54, 4475–4483. [Google Scholar] [CrossRef]
- Oshiro, N.; Nagasawa, H.; Kuniyoshi, K.; Kobayashi, N.; Sugita-Konishi, Y.; Asakura, H.; Yasumoto, T. Characteristic Distribution of Ciguatoxins in the Edible Parts of a Grouper, Variola Louti. Toxins 2021, 13, 218. [Google Scholar] [CrossRef]
- Ramos-Sosa, M.J.; García-Álvarez, N.; Sanchez-Henao, A.; Padilla, D.; Sergent, F.S.; Gago-Martínez, A.; Diogène, J.; Caballero, M.J.; Fernández, A.; Real, F. Ciguatoxin-like Toxicity Distribution in Flesh of Amberjack (Seriola spp.) and Dusky Grouper (Epinephelus Marginatus). Environ. Res. 2023, 228, 115869. [Google Scholar] [CrossRef] [PubMed]
- Pottier, I.; Vernoux, J.P. Evaluation of Antilles fish ciguatoxicity by mouse and chick bioassays. Bull. Soc. Pathol. Exot. 1990 2003, 96, 24–28. [Google Scholar]
- Hahn, S.T.; Capra, M.F.; Walsh, T.P. Ciguatoxin-Protein Association in Skeletal Muscle of Spanish Mackerel (Scomberomorus Commersoni). Toxicon 1992, 30, 843–852. [Google Scholar] [CrossRef]
- Capra, M.F.; Cameron, J.; Flowers, A.E.; Purcell, C.E. Response of Vertebrate Nerves to Ciguatoxins. Mem. Qld. Mus. 1994, 34, 454. [Google Scholar]
- Ho, B.; Yeo, D.S.A.; Ding, J.L. A Tetrodotoxin Neutralizing System in the Haemolymph of the Horseshoe Crab, Carcinoscorpius Rotundicauda. Toxicon 1994, 32, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, K.; Yamaguchi, S.; Kikuchi, T.; Yamamori, K.; Matsui, T. Occurrence of Tetrodotoxin-Binding High Molecular Weight Substances in the Body Fluid of Shore Crab (Hemigrapsus Sanguineus). Toxicon Off. J. Int. Soc. Toxinol. 1992, 30, 1529–1537. [Google Scholar] [CrossRef]
- Mak, Y.L.; Wu, J.J.; Chan, W.H.; Murphy, M.B.; Lam, J.C.W.; Chan, L.L.; Lam, P.K.S. Simultaneous Quantification of Pacific Ciguatoxins in Fish Blood Using Liquid Chromatography–Tandem Mass Spectrometry. Anal. Bioanal. Chem. 2013, 405, 3331–3340. [Google Scholar] [CrossRef]
- Bottein Dechraoui, M.-Y.; Wang, Z.; Ramsdell, J.S. Optimization of Ciguatoxin Extraction Method from Blood for Pacific Ciguatoxin (P-CTX-1). Toxicon 2007, 49, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Bottein Dechraoui, M.-Y.; Rezvani, A.H.; Gordon, C.J.; Levin, E.D.; Ramsdell, J.S. Repeat Exposure to Ciguatoxin Leads to Enhanced and Sustained Thermoregulatory, Pain Threshold and Motor Activity Responses in Mice: Relationship to Blood Ciguatoxin Concentrations. Toxicology 2008, 246, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Matta, J.; Navas, J.; Milad, M.; Manger, R.; Hupka, A.; Frazer, T. A Pilot Study for the Detection of Acute Ciguatera Intoxication in Human Blood. J. Toxicol. Clin. Toxicol. 2002, 40, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Gwinn, J.K.; Uhlig, S.; Ivanova, L.; Fæste, C.K.; Kryuchkov, F.; Robertson, A. In Vitro Glucuronidation of Caribbean Ciguatoxins in Fish: First Report of Conjugative Ciguatoxin Metabolites. Chem. Res. Toxicol. 2021, 34, 1910–1925. [Google Scholar] [CrossRef]
- Vernoux, J.-P. The Mouse Ciguatoxin Bioassay: Directions for Use to Control Fish for Consumption. Mem. Qld. Mus. 1994, 34, 625–629. [Google Scholar]
- Nukina, M.; Koyanagi, L.M.; Scheuer, P.J. Two Interchangeable Forms of Ciguatoxin. Toxicon 1984, 22, 169–176. [Google Scholar] [CrossRef]
- Talha, F.T. Séparation et Purification de Quelques Ciguatoxines. Ph.D. Thesis, Université Bordeaux I, Bordeaux, France, 1987. [Google Scholar]
- Harwood, D.T.; Murray, S.; Boundy, M.J. Chapter Three–Sample Preparation Prior to Marine Toxin Analysis. In Comprehensive Analytical Chemistry; Diogène, J., Campàs, M., Eds.; Recent Advances in the Analysis of Marine Toxins; Elsevier: Amsterdam, The Netherlands, 2017; Volume 78, pp. 89–136. [Google Scholar]
- Caillaud, A.; de la Iglesia, P.; Darius, H.T.; Pauillac, S.; Aligizaki, K.; Fraga, S.; Chinain, M.; Diogène, J. Update on Methodologies Available for Ciguatoxin Determination: Perspectives to Confront the Onset of Ciguatera Fish Poisoning in Europe. Mar. Drugs 2010, 8, 1838–1907. [Google Scholar] [CrossRef]
- Lewis, R.J.; Sellin, M. Multiple Ciguatoxins in the Flesh of Fish. Toxicon 1992, 30, 915–919. [Google Scholar] [CrossRef] [PubMed]
- Poli, M.A.; Lewis, R.J.; Dickey, R.W.; Musser, S.M.; Buckner, C.A.; Carpenter, L.G. Identification of Caribbean Ciguatoxins as the Cause of an Outbreak of Fish Poisoning among U.S. Soldiers in Haiti. Toxicon 1997, 35, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Hardison, D.R.; Holland, W.C.; McCall, J.R.; Bourdelais, A.J.; Baden, D.G.; Darius, H.T.; Chinain, M.; Tester, P.A.; Shea, D.; Quintana, H.A.F.; et al. Fluorescent Receptor Binding Assay for Detecting Ciguatoxins in Fish. PLoS ONE 2016, 11, e0153348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliño, L.; Widgy, S.; Pautonnier, A.; Turquet, J.; Loeffler, C.R.; Flores Quintana, H.A.; Diogène, J. Prevalence of Ciguatoxins in Lionfish (Pterois spp.) from Guadeloupe, Saint Martin, and Saint Barthélmy Islands (Caribbean). Toxicon 2015, 102, 62–68. [Google Scholar] [CrossRef]
- Estevez, P.; Castro, D.; Pequeño-Valtierra, A.; Leao, J.M.; Vilariño, O.; Diogène, J.; Gago-Martínez, A. An Attempt to Characterize the Ciguatoxin Profile in Seriola Fasciata Causing Ciguatera Fish Poisoning in Macaronesia. Toxins 2019, 11, 221. [Google Scholar] [CrossRef] [Green Version]
- Costa, P.R.; Estévez, P.; Soliño, L.; Castro, D.; Rodrigues, S.M.; Timoteo, V.; Leao-Martins, J.M.; Santos, C.; Gouveia, N.; Diogène, J.; et al. An Update on Ciguatoxins and CTX-like Toxicity in Fish from Different Trophic Levels of the Selvagens Islands (NE Atlantic, Madeira, Portugal). Toxins 2021, 13, 580. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Yang, A.; Jones, A. Rapid Extraction Combined with LC-Tandem Mass Spectrometry (CREM-LC/MS/MS) for the Determination of Ciguatoxins in Ciguateric Fish Flesh. Toxicon 2009, 54, 62–66. [Google Scholar] [CrossRef]
- Stewart, I.; Eaglesham, G.K.; Poole, S.; Graham, G.; Paulo, C.; Wickramasinghe, W.; Sadler, R.; Shaw, G.R. Establishing a Public Health Analytical Service Based on Chemical Methods for Detecting and Quantifying Pacific Ciguatoxin in Fish Samples. Toxicon 2010, 56, 804–812. [Google Scholar] [CrossRef]
- Meyer, L.; Carter, S.; Capper, A. An Updated Ciguatoxin Extraction Method and Silica Cleanup for Use with HPLC-MS/MS for the Analysis of P-CTX-1, PCTX-2 and P-CTX-3. Toxicon 2015, 108, 249–256. [Google Scholar] [CrossRef]
- Spielmeyer, A.; Loeffler, C.R.; Bodi, D. Extraction and LC-MS/MS Analysis of Ciguatoxins: A Semi-Targeted Approach Designed for Fish of Unknown Origin. Toxins 2021, 13, 630. [Google Scholar] [CrossRef]
- Wu, J.J.; Mak, Y.L.; Murphy, M.B.; Lam, J.C.W.; Chan, W.H.; Wang, M.; Chan, L.L.; Lam, P.K.S. Validation of an Accelerated Solvent Extraction Liquid Chromatography–Tandem Mass Spectrometry Method for Pacific Ciguatoxin-1 in Fish Flesh and Comparison with the Mouse Neuroblastoma Assay. Anal. Bioanal. Chem. 2011, 400, 3165–3175. [Google Scholar] [CrossRef] [PubMed]
- Lehane, L. Ciguatera Update. Med. J. Aust. 2000, 172, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J. Ciguatoxins Are Potent Ichthyotoxins. Toxicon 1992, 30, 207–211. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Health and Human Services Food and Drug Administration Fish and Fishery Products Hazards and Controls Guidance. Available online: https://www.fda.gov/media/80637/download (accessed on 29 December 2022).
- EFSA Panel on Contaminants in the Food Chain Scientific Opinion on Marine Biotoxins in Shellfish–Emerging Toxins: Ciguatoxin Group. EFSA J. 2010, 8, 1627. [CrossRef]
- Hoffman, P.A.; Granade, H.R.; McMillan, J.P. The Mouse Ciguatoxin Bioassay: A Dose - Response Curve and Symptomatology Analysis. Toxicon 1983, 21, 363–369. [Google Scholar] [CrossRef]
- Manger, R.; Woodle, D.; Berger, A.; Dickey, R.W.; Jester, E.; Yasumoto, T.; Lewis, R.; Hawryluk, T.; Hungerford, J. Flow Cytometric-Membrane Potential Detection of Sodium Channel Active Marine Toxins: Application to Ciguatoxins in Fish Muscle and Feasibility of Automating Saxitoxin Detection. J. AOAC Int. 2014, 97, 299–306. [Google Scholar] [CrossRef]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Wekell, M.M. Tetrazolium-Based Cell Bioassay for Neurotoxins Active on Voltage-Sensitive Sodium Channels: Semiautomated Assay for Saxitoxins, Brevetoxins, and Ciguatoxins. Anal. Biochem. 1993, 214, 190–194. [Google Scholar] [CrossRef]
- Viallon, J.; Chinain, M.; Darius, H.T. Revisiting the Neuroblastoma Cell-Based Assay (CBA-N2a) for the Improved Detection of Marine Toxins Active on Voltage Gated Sodium Channels (VGSCs). Toxins 2020, 12, 281. [Google Scholar] [CrossRef]
- Darius, H.T.; Ponton, D.; Revel, T.; Cruchet, P.; Ung, A.; Tchou Fouc, M.; Chinain, M. Ciguatera Risk Assessment in Two Toxic Sites of French Polynesia Using the Receptor-Binding Assay. Toxicon 2007, 50, 612–626. [Google Scholar] [CrossRef]
- Díaz-Asencio, L.; Clausing, R.J.; Vandersea, M.; Chamero-Lago, D.; Gómez-Batista, M.; Hernández-Albernas, J.I.; Chomérat, N.; Rojas-Abrahantes, G.; Litaker, R.W.; Tester, P.; et al. Ciguatoxin Occurrence in Food-Web Components of a Cuban Coral Reef Ecosystem: Risk-Assessment Implications. Toxins 2019, 11, 722. [Google Scholar] [CrossRef] [Green Version]
- McCall, J.R.; Jacocks, H.M.; Niven, S.C.; Poli, M.A.; Baden, D.G.; Bourdelais, A.J. Development and Utilization of a Fluorescence-Based Receptor-Binding Assay for the Site 5 Voltage-Sensitive Sodium Channel Ligands Brevetoxin and Ciguatoxin. J. AOAC Int. 2014, 97, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Yasumoto, T. Chemiluminescent Receptor Binding Assay for Ciguatoxins and Brevetoxins Using Acridinium Brevetoxin-B2. Toxins 2019, 11, 580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dechraoui, M.-Y.B.; Tiedeken, J.A.; Persad, R.; Wang, Z.; Granade, H.R.; Dickey, R.W.; Ramsdell, J.S. Use of Two Detection Methods to Discriminate Ciguatoxins from Brevetoxins: Application to Great Barracuda from Florida Keys. Toxicon 2005, 46, 261–270. [Google Scholar] [CrossRef]
- Lewis, R.J. Detection of Toxins Associated with Ciguatera Fish Poisoning. In Manual on Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, R.A., Cembella, A.D., Eds.; Monographs on oceanographic methodology; Unesco: Paris, France, 2004; Volume 11, pp. 267–277. [Google Scholar]
- Lewis, R.J. Detection of Ciguatoxins and Related Benthic Dinoflagellate Toxins: In Vivo and in Vitro Methods. In Manual on Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, R.A., Cembella, A.D., Eds.; IOC Manuals and Guides; Unesco: Paris, France, 1995; Volume 33, pp. 135–161. [Google Scholar]
- Suzuki, T.; Ha, D.V.; Uesugi, A.; Uchida, H. Analytical Challenges to Ciguatoxins. Curr. Opin. Food Sci. 2017, 18, 37–42. [Google Scholar] [CrossRef]
- Reverté, L.; Soliño, L.; Carnicer, O.; Diogène, J.; Campàs, M. Alternative Methods for the Detection of Emerging Marine Toxins: Biosensors, Biochemical Assays and Cell-Based Assays. Mar. Drugs 2014, 12, 5719–5763. [Google Scholar] [CrossRef] [Green Version]
- Tosteson, T.R.; Ballantine, D.L.; Durst, H.D. Seasonal Frequency of Ciguatoxic Barracuda in Southwest Puerto Rico. Toxicon Off. J. Int. Soc. Toxinol. 1988, 26, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Jones, A.; Vernoux, J.-P. HPLC/Tandem Electrospray Mass Spectrometry for the Determination of Sub-Ppb Levels of Pacific and Caribbean Ciguatoxins in Crude Extracts of Fish. Anal. Chem. 1999, 71, 247–250. [Google Scholar] [CrossRef]
- Moreiras, G.; Leão, J.M.; Gago-Martínez, A. Design of Experiments for the Optimization of Electrospray Ionization in the LC-MS/MS Analysis of Ciguatoxins. J. Mass Spectrom. 2018, 53, 1059–1069. [Google Scholar] [CrossRef]
- Sasaki, M.; Iwasaki, K.; Arai, K. Studies toward the Total Synthesis of Caribbean Ciguatoxin C-CTX-1: Synthesis of the LMN-Ring Fragment through Reductive Olefin Cross-Coupling. Org. Lett. 2018, 20, 7163–7166. [Google Scholar] [CrossRef]
- Sasaki, M.; Iwasaki, K.; Arai, K. Synthesis and Structural Implication of the JKLMN-Ring Fragment of Caribbean Ciguatoxin C-CTX-1. J. Org. Chem. 2021, 86, 4580–4597. [Google Scholar] [CrossRef]
- Estevez, P.; Sibat, M.; Leão-Martins, J.M.; Reis Costa, P.; Gago-Martínez, A.; Hess, P. Liquid Chromatography Coupled to High-Resolution Mass Spectrometry for the Confirmation of Caribbean Ciguatoxin-1 as the Main Toxin Responsible for Ciguatera Poisoning Caused by Fish from European Atlantic Coasts. Toxins 2020, 12, 267. [Google Scholar] [CrossRef] [Green Version]
- Kryuchkov, F.; Robertson, A.; Mudge, E.M.; Miles, C.O.; Van Gothem, S.; Uhlig, S. Reductive Amination for LC–MS Signal Enhancement and Confirmation of the Presence of Caribbean Ciguatoxin-1 in Fish. Toxins 2022, 14, 399. [Google Scholar] [CrossRef] [PubMed]
- Mudge, E.M.; Robertson, A.; Leynse, A.K.; McCarron, P.; Miles, C.O. Selective Extraction of Gambierone and Related Metabolites in Gambierdiscus Silvae Using m-Aminophenylboronic Acid–Agarose Gel and Liquid Chromatography–High-Resolution Mass Spectrometric Detection. J. Chromatogr. B 2022, 1188, 123014. [Google Scholar] [CrossRef] [PubMed]
- Estevez, P.; Sibat, M.; Leão-Martins, J.M.; Tudó, A.; Rambla-Alegre, M.; Aligizaki, K.; Diogène, J.; Gago-Martinez, A.; Hess, P. Use of Mass Spectrometry to Determine the Diversity of Toxins Produced by Gambierdiscus and Fukuyoa Species from Balearic Islands and Crete (Mediterranean Sea) and the Canary Islands (Northeast Atlantic). Toxins 2020, 12, 305. [Google Scholar] [CrossRef] [PubMed]
- Yon, T.; Sibat, M.; Robert, E.; Lhaute, K.; Holland, W.C.; Litaker, R.W.; Bertrand, S.; Hess, P.; Réveillon, D. Sulfo-Gambierones, Two New Analogs of Gambierone Produced by Gambierdiscus Excentricus. Mar. Drugs 2021, 19, 657. [Google Scholar] [CrossRef]
- Dickey, R.W. Ciguatera Toxins: Chemistry, Toxicology, and Detection; CRC Press: New York, NY, USA, 2008; pp. 497–518. ISBN 978-0-429-12850-9. [Google Scholar]
- Nagae, M.; Igarashi, T.; Mizukoshi, K.; Kuniyoshi, K.; Oshiro, N.; Yasumoto, T. Development and Validation of an LC-MS/MS Method for the Ultra-Trace Analysis of Pacific Ciguatoxins in Fish. J. AOAC Int. 2021, 104, 1272–1281. [Google Scholar] [CrossRef]
- Sibat, M.; Herrenknecht, C.; Darius, H.T.; Roué, M.; Chinain, M.; Hess, P. Detection of Pacific Ciguatoxins Using Liquid Chromatography Coupled to Either Low or High Resolution Mass Spectrometry (LC-MS/MS). J. Chromatogr. A 2018, 1571, 16–28. [Google Scholar] [CrossRef]
- Estevez, P.; Gago-Martinez, A. Contribution of Mass Spectrometry to the Advances in Risk Characterization of Marine Biotoxins: Towards the Characterization of Metabolites Implied in Human Intoxications. Toxins 2023, 15, 103. [Google Scholar] [CrossRef]
- AOAC Official Method 959.08. Paralytic Shellfish Poison. Biological Method. Final Action. In AOAC Official Methods for Analysis, Natural Toxins (Chapter 49); AOAC International: Gaithersburg, MD, USA, 2005; pp. 79–80.
- OECD. Test No. 425: Acute Oral Toxicity: Up-and-Down Procedure. In OECD Guidelines for the Testing of Chemicals, Section 4; OECD: Paris, France, 2022. [Google Scholar]
- Finch, S.C.; Boundy, M.J.; Webb, N.G.; Harwood, D.T. The Effect of Experimental Protocol on the Toxicity of Saxitoxin in Mice. Toxins 2023, 15, 290. [Google Scholar] [CrossRef]
- Turner, A.D.; Tarnovius, S.; Hatfield, R.G.; Teixeira-Alves, M.; Broadwater, M.; Dolah, F.V.; Garcia-Mendoza, E.; Medina, D.; Salhi, M.; Goya, A.B.; et al. Application of Six Detection Methods for Analysis of Paralytic Shellfish Toxins in Shellfish from Four Regions within Latin America. Mar. Drugs 2020, 18, 616. [Google Scholar] [CrossRef]
- Turner, A.D.; Hatfield, R.G.; Maskrey, B.H.; Algoet, M.; Lawrence, J.F. Evaluation of the New European Union Reference Method for Paralytic Shellfish Toxins in Shellfish: A Review of Twelve Years Regulatory Monitoring Using Pre-Column Oxidation LC-FLD. TrAC Trends Anal. Chem. 2019, 113, 124–139. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTX | Other Names | Molecular Formula | Molecular Mass (Da) | Known Source | Structural Characteristics |
|---|---|---|---|---|---|
| Type 1 (60 carbons, A to M rings, DEF rings size 7/7/9) | |||||
| P-CTX-1 | CTX-1B, CTX, CTX1 | C60H86O19 [98] | 1110.6 | Fish | |
| P-CTX-3 | 54-deoxy-CTX-1B | C60H86O18 [122] | 1094.6 | Fish and Gambierdiscus spp. | Two epimers without any -OH group at C54, compared to P-CTX-1 |
| P-CTX-2 | 52-epi-54-deoxy-CTX-1B | C60H86O18 [125] | 1094.6 | ||
| GT-4Ba | GTX-4B, CTX-4B, 52-epi-P-CTX-4A | C60H84O16 [98] | 1060.6 | Gambierdiscus/Fukuyoa spp. and herbivorous fish | Two epimers at C52 differing from P-CTX-1 by the absence of 2 -OH on A-ring and one -OH on M-ring |
| CTX-4A a | P-CTX-4A | C60H84O16 [126] | 1060.6 | ||
| Type 2 (57 carbons, A to M rings, DEF rings size 7/8/9) | |||||
| CTX-3Ca | P-CTX-3C | C57H82O16 [123] | 1022.6 | Gambierdiscus/Fukuyoa spp. | Two epimers at C49 |
| CTX-3B | P-CTX-3B, 49-epi-P-CTX-3C | C57H82O16 [119] | 1022.6 | ||
| Type 3 (62 carbons, A to N rings, DEF rings size 7/8/9) | |||||
| C-CTX-1 | C62H92O19 [35] | 1140.6 | Fish | Two epimers at C52 | |
| C-CTX-2 | C62H92O19 [35] | 1140.6 | Fish | ||
| C-CTX-3 | C-CTX-1143 | C62H94O19 [120] | 1142.6 | Fish | Two anomers of the hemiketal ring opened between C52 and C56, reduced forms of C-CTX-1/-2 |
| C-CTX-4 | C-CTX-1143 | C62H94O19 [120] | 1142.6 | Fish | |
| C-CTX-5 b | C62H90O19 [43] | 1138.6 | G. silvae, G. caribaeus | C-CTX-1 with the OH at C3 reduced to a ketone | |
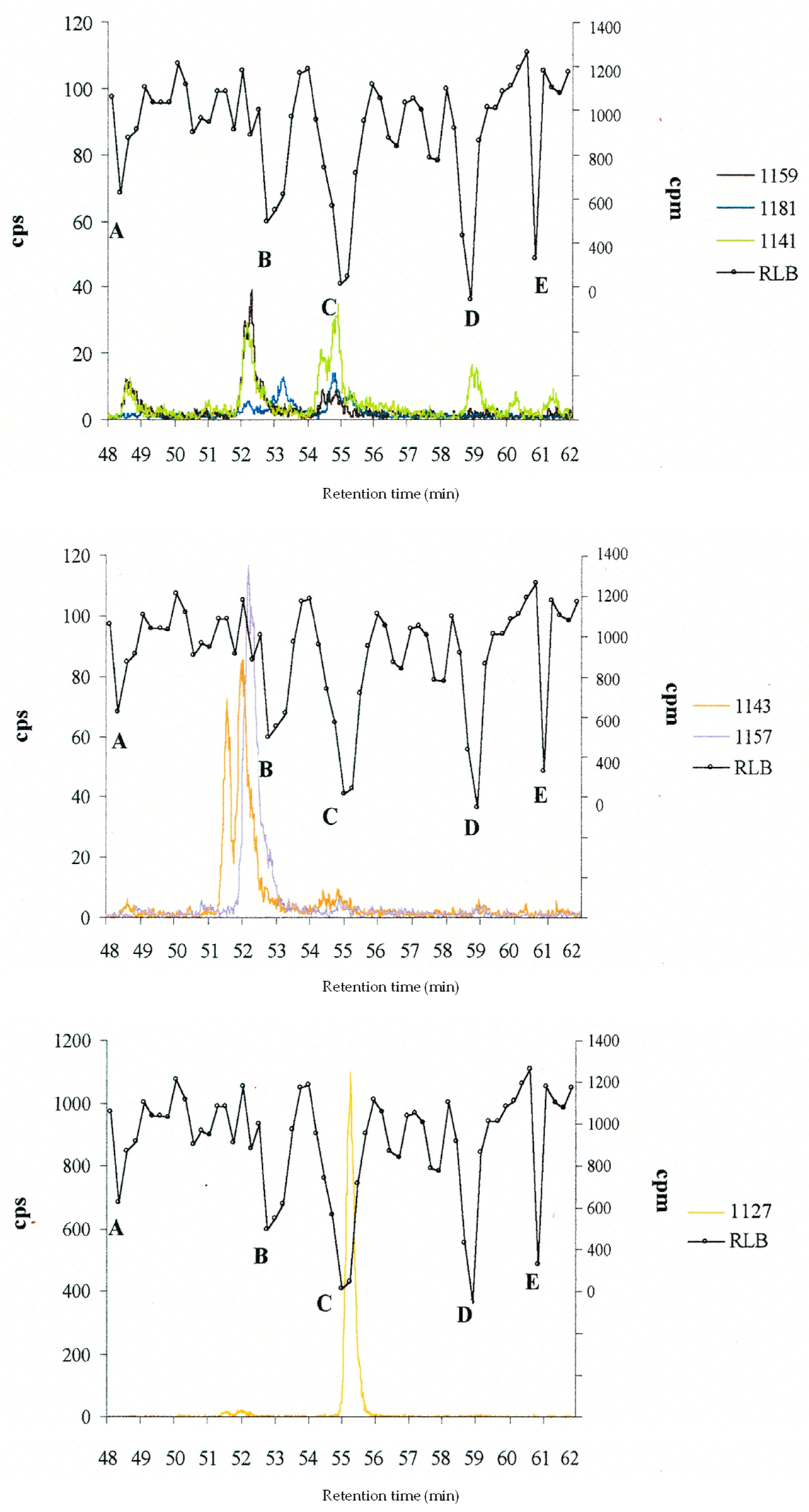
| Retention Time (min) | [M+H]+ m/z | C-CTX- | RLB Peaks (Relative Intensity) |
|---|---|---|---|
| 48.51 | 1159.58 859.42 * 811.47 ** | 1159 | A (+) |
| 51.55 | 1143.57 | 1143a | no peak |
| 52.03 | 1143.60 | 1143 | no peak |
| 52.17 | 1157.57 | 1157 | B (++) |
| 53.20 | 1181.60 | 1181 | |
| 54.47 | 1141.58 | 1141a | |
| 54.84 | 1141.58 | 1 | C (+++) |
| 55.26 | 1127.57 | 1127 | |
| 58.99 | 1141.61 | 2 | D (+++) |
| 60.32 | 1141.58 | 1141b | no peak |
| 61.38 | 1141.58 | 1141c | E (+++) |
| References | [109] | [110] | [175] | [176] | [112] |
| LC conditions | |||||
| LC column | Agilent Poroshell 120-EC C18 (50 × 3 mm, 2.7 µm) | ||||
| Mobile phases A and B | A: 5 mM ammonium formate and 0.1% formic acid in H2O; B: Methanol | ||||
| [M+Na]+ m/z for precursor and product ions in MRM for CTXs type 1 | |||||
| 1133.6 (P-CTX-1) | + # | + # | + # | + # | + # |
| 1117.6 (P-CTX-2/-3) | + # | + # | + # | + # | + # |
| 1083.6 (GT-4B/CTX-4A) | + # | + # | + # | + # | + # |
| 1101.6 (M-secoCTX-4B/-4A) | u.m. | + # | u.m. | u.m. | u.m. |
| [M+Na]+ m/z for precursor and product ions in MRM for CTXs type 2 | |||||
| 1045.6 (CTX-3C/-3B) | + # | + # | + # | + # | + # |
| 1061.6 (51-hydroxyCTX3C) | + # | + # | + # | + # | u.m. |
| 1063.6 (2-hydroxy-CTX3C) | u.m. | + # | + # | u.m. | u.m. |
| 1079.6 (2,3-dihydroxyCTX-3C) | + # | + # | + # | + # | u.m. |
| 1063.6 (M-seco-CTX3C) | u.m. | + # | u.m. | u.m. | u.m. |
| 1077.6 (M-secoCTX3C-methylacetal) | u.m. | + # | u.m. | u.m. | u.m. |
| [M+Na]+ m/z for precursor and product ions in MRM for C-CTXs type 3 | |||||
| 1163.7 (C-CTX-1) | + # | + # | + # | + # | + # |
| 1177.6 (C-CTX-1-Me) | u.m. | + | u.m. | + # | + # |
| Results for different fish | |||||
| Fish sampling (fishing spot) | 11 fish a (SI) 9 fish b (M) | 3 fish c (CI) 1 fish d (SI) | 1 fish e (SI) | 56 fish f (SI) | 109 fish g (CI) (flesh, liver) |
| Cytotoxicity levels by CBA-N2a (min–max) | not used | Positive CTX-like toxicity | 1.4 ppb * | 0.006–0.75 ppb ** | <LOQ–1.365 ppb ** (93 flesh) <LOQ–6.4390 ppb** (107 livers) |
| Fish C-CTX-1 levels by LC-MS/MS (min–max) | C-CTX-1 # (8 fish, SI) <LOQ–0.25 ppb * | C-CTX-1 # (4 fish) 0.12–0.76 ppb * | C-CTX-1 # (1 fish) 0.84 ppb * | C-CTX-1 # (20 fish) <LOQ–0.48 ppb * | C-CTX-1 # (30 fish/62 analysed) 0.018–0.270 ppb * |
| References | [108] | [39] | [174] | [37] |
| LC conditions | ||||
| LC column | Phenomenex Luna C8(2) (150 × 2.0 mm) | Phenomenex Luna C18 (100 × 2 mm, 3 μm) | Phenomenex Kinetex C8 (75 × 2.1 mm, 2.6 µm) | Phenomenex Kinetex C8 (75 × 2.1 mm, 2.6 µm) |
| Mobile phase A | H2O | H2O | H2O | H2O |
| Mobile phase B | 100% acetonitrile | 95% acetonitrile | 95% acetonitrile | 95% acetonitrile |
| Modifier (A and B) | 0.1% formic acid | 5 mM ammonium acetate | 5 mM ammonium formate | 0.1% formic acid |
| MRM transitions for C-CTX-1 detection | ||||
| [M+H–H2O]+ m/z precursor ion | 1123.6 | |||
| [M+H–nH2O]+ m/z product ions | 1105.6; 1087.6; 1069.6 | 1105.7; 1087.7; 1069.8 | 1087.6; 1069.7 | 1105.6; 1087.6; 1069.6 |
| Results for different fish | ||||
| Fish sampling (fishing spot) | 1 barracuda a (UO) | 153 lionfish b (VI) | 60 lionfish b 55 (SB), 5 (SM), 30 (G) | 77 fish c (VI) |
| Cytotoxicity levels by CBA-N2a | 1.6 ppb * (cooked) 2.1 ppb * (raw) | <0.1 ppb * (43 fish) >0.1 ppb * (19 fish) | <0.01 ppb ** (5 fish, SB) 0.01–0.1 ppb ** (14 fish, SB) >0.1 ppb ** (8 fish, SB) | ≥0.005 ppb * (29 fish) >0.1 ppb * (3 fish) |
| C-CTX-1 confirmation by LC-MS/MS | C-CTX-1 # (1 fish) | C-CTX-1 # (19 fish) | C-CTX-1 # (8 fish, SB) | C-CTX-1 # (13 fish) |
| C-CTX congeners detected by LC-MS/MS | C-CTX-1159, -1157 and -1143 | C-CTX-2 # | - | - |
| References → | [110] | [175] | [176] |
| LC conditions | |||
| LC column | Agilent Poroshell 120-EC C18 (100 × 2.1 mm, 2.7 µm) | ||
| Mobile phases | (A) 5 mM ammonium formate and 0.1% formic acid in H2O; (B) acetonitrile | ||
| Precursor and Product ions for confirmation of C-CTX-1 # and isomers | |||
| [M+H–H2O]+ m/z 1123.6 | C-CTX-1 [M+H–nH2O]+ m/z 1105.6; 1087.6; 1069.6 | ||
| specific fragments m/z 191.1; 108.9 | |||
| Precursor and Product ions for confirmation of C-CTX-1157 | |||
| [M+Na]+ m/z 1179.6 | u.m. | [M+Na]+ m/z 1179.6 | |
| [M+H–H2O]+ m/z 1139.6 | u.m. | [M+H–nH2O]+ m/z 1121.6; 1103.6; 1085.6 | [M+H–nH2O]+ m/z 1121.6; 1103.6; 1085.6 |
| u.m. | specific fragments m/z 191.1; 108.9 | specific fragments m/z 191.1; 108.9 | |
| Precursor and Product ions for confirmation of C-CTX-1127 | |||
| [M+Na]+ m/z 1149.6 | u.m. | C-CTX-1127 [M+Na]+ m/z 1149.6 | u.m. |
| [M+H]+ m/z 1127.6 | u.m. | [M+H–nH2O]+ m/z 1109.6; 1091.6; 1073.6; 1055.6 | u.m. |
| Precursor and Product ions for confirmation of C-CTX-1-Me | |||
| [M+H–CH3–H2O]+ m/z 1123.6 | u.m. | u.m. | [M+H–nH2O]+ m/z 1105.6; 1087.6; 1069.6 |
| u.m. | u.m. | specific fragments m/z 191.1; 108.9 | |
| Results for studied fish specimens | |||
| Fish sampling (fishing spot) | 3 fish a (CI) 1 fish b (SI) | 1 fish c (SI) | 56 fish d (SI) |
| Cytotoxicity levels by CBA-N2a (min–max) | Positive | 1.4 ppb * | 0.006–0.75 ppb ** |
| C-CTX-1 confirmation by LC-MS/MS | C-CTX-1 (4 fish) | C-CTX-1 (1 fish) | C-CTX-1 (20 fish) |
| C-CTX congeners detected by LC-MS/MS | C-CTX-1181 * | C-CTX-1157 *, C-CTX-1127 *, C-CTX-1 isomer * | C-CTX-1157 |
| Review Section | Organic Eluent | Eluent Modifier | Goal (MS Mode) | Precursor Ions | References |
|---|---|---|---|---|---|
| 5.2.2 | Methanol | Formic acid and ammonium formate | C-CTX-1 quantification (SRM) | [M+Na]+ | [109,110,112,155,175,176] |
| 5.2.3 | Acetonitrile | Formic acid or ammonium formate or ammonium acetate | C-CTX-1 confirmation (MRM) | [M+H–H2O]+ | [37,39,108,174] |
| C-CTXs analogues identification (MRM) | [M+Na]+; [M+H–H2O]; [M+H]+, [M+H–CH3–H2O]+ | [110,175,176] | |||
| 5.2.4 | Methanol and acetonitrile (3:1) | Formic acid and ammonium acetate | 4 CTX groups identification (MRM) | [M+Na]+ | [34,180] |
| CTXs confirmation (MRM) | [M+NH4]+ |
| Name | C-CTX-1159 | C-CTX-3/-4 [120] | C-CTX-1157 | C-CTX-1127 |
|---|---|---|---|---|
| [M+H]+ m/z | 1159.6 | 1143.6 | 1157.6 | 1127.6 |
| Polarity * | +  − − | |||
| Reported in Caribbean fish | [42,108] | [23,38,41,42,108,120] | [23,38,41,42,108] | [23,41,42] |
| Reported in fish from Macaronesia | [110] | [131] | [175,176] | [113,175] |
| Potential toxicity | [108,110] | [108] | [40,108] | [175] |
| Putative structure | Hydroxylated C-CTX-1 or oxidised C-CTX-1143 | reduced C-CTX-1/-2 hemiketals [120] | Oxidised C-CTX-1 | Loss of CH2 or demethylated C-CTX-1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pottier, I.; Lewis, R.J.; Vernoux, J.-P. Ciguatera Fish Poisoning in the Caribbean Sea and Atlantic Ocean: Reconciling the Multiplicity of Ciguatoxins and Analytical Chemistry Approach for Public Health Safety. Toxins 2023, 15, 453. https://doi.org/10.3390/toxins15070453
Pottier I, Lewis RJ, Vernoux J-P. Ciguatera Fish Poisoning in the Caribbean Sea and Atlantic Ocean: Reconciling the Multiplicity of Ciguatoxins and Analytical Chemistry Approach for Public Health Safety. Toxins. 2023; 15(7):453. https://doi.org/10.3390/toxins15070453
Chicago/Turabian StylePottier, Ivannah, Richard J. Lewis, and Jean-Paul Vernoux. 2023. "Ciguatera Fish Poisoning in the Caribbean Sea and Atlantic Ocean: Reconciling the Multiplicity of Ciguatoxins and Analytical Chemistry Approach for Public Health Safety" Toxins 15, no. 7: 453. https://doi.org/10.3390/toxins15070453
APA StylePottier, I., Lewis, R. J., & Vernoux, J. -P. (2023). Ciguatera Fish Poisoning in the Caribbean Sea and Atlantic Ocean: Reconciling the Multiplicity of Ciguatoxins and Analytical Chemistry Approach for Public Health Safety. Toxins, 15(7), 453. https://doi.org/10.3390/toxins15070453