Tetrodotoxins in Tissues and Cells of Different Body Regions of Ribbon Worms Kulikovia alborostrata and K. manchenkoi from Spokoynaya Bay, Sea of Japan


Abstract
:1. Introduction
2. Results
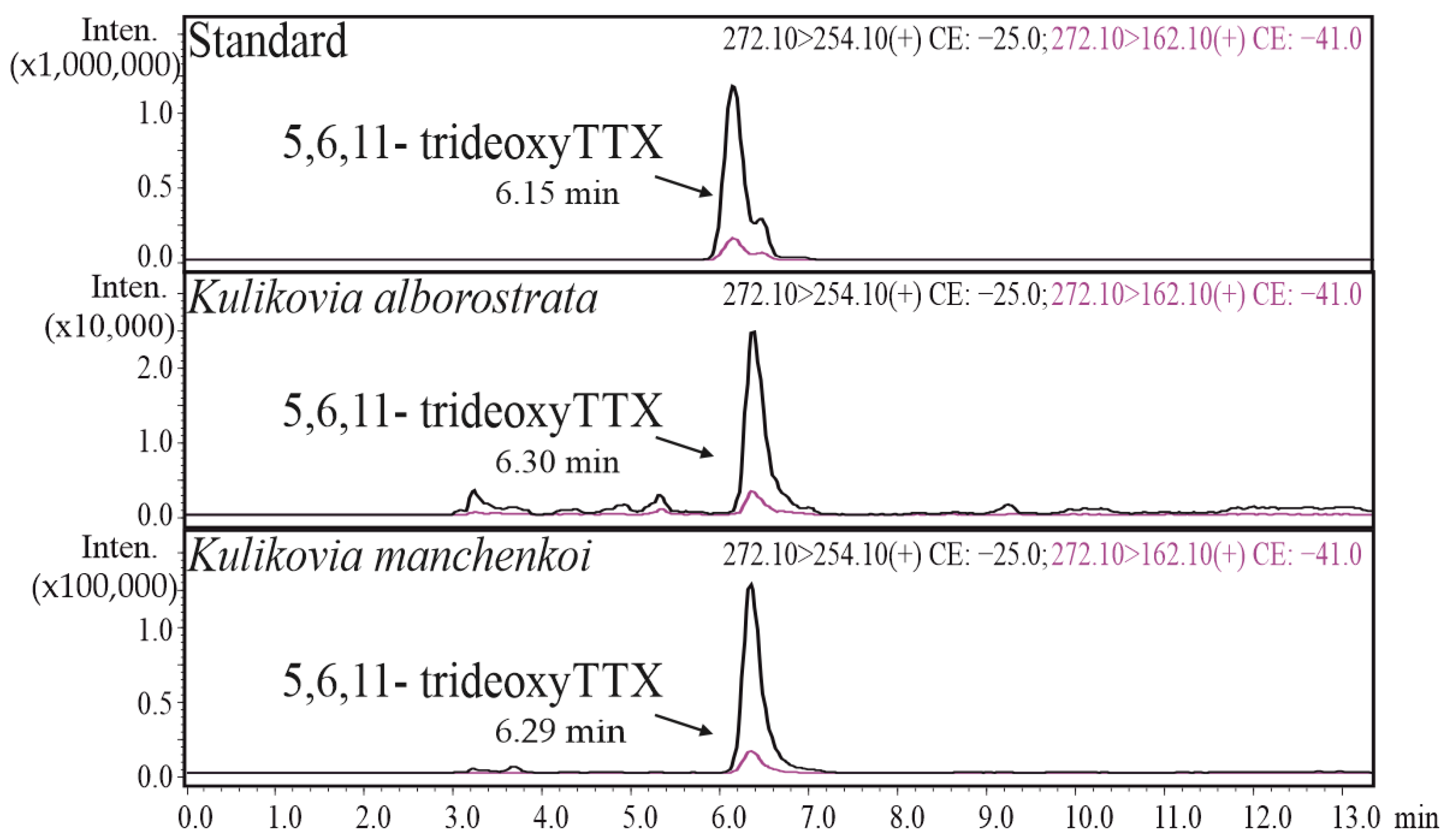
2.1. HPLC-MS/MS
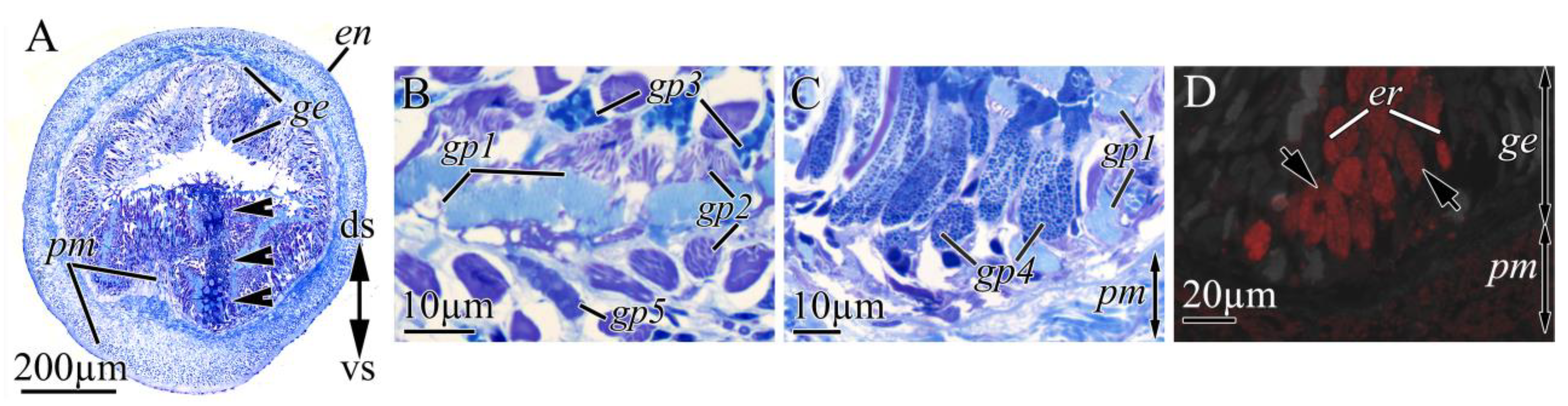
2.2. Morphological and Immunohistochemical Studies
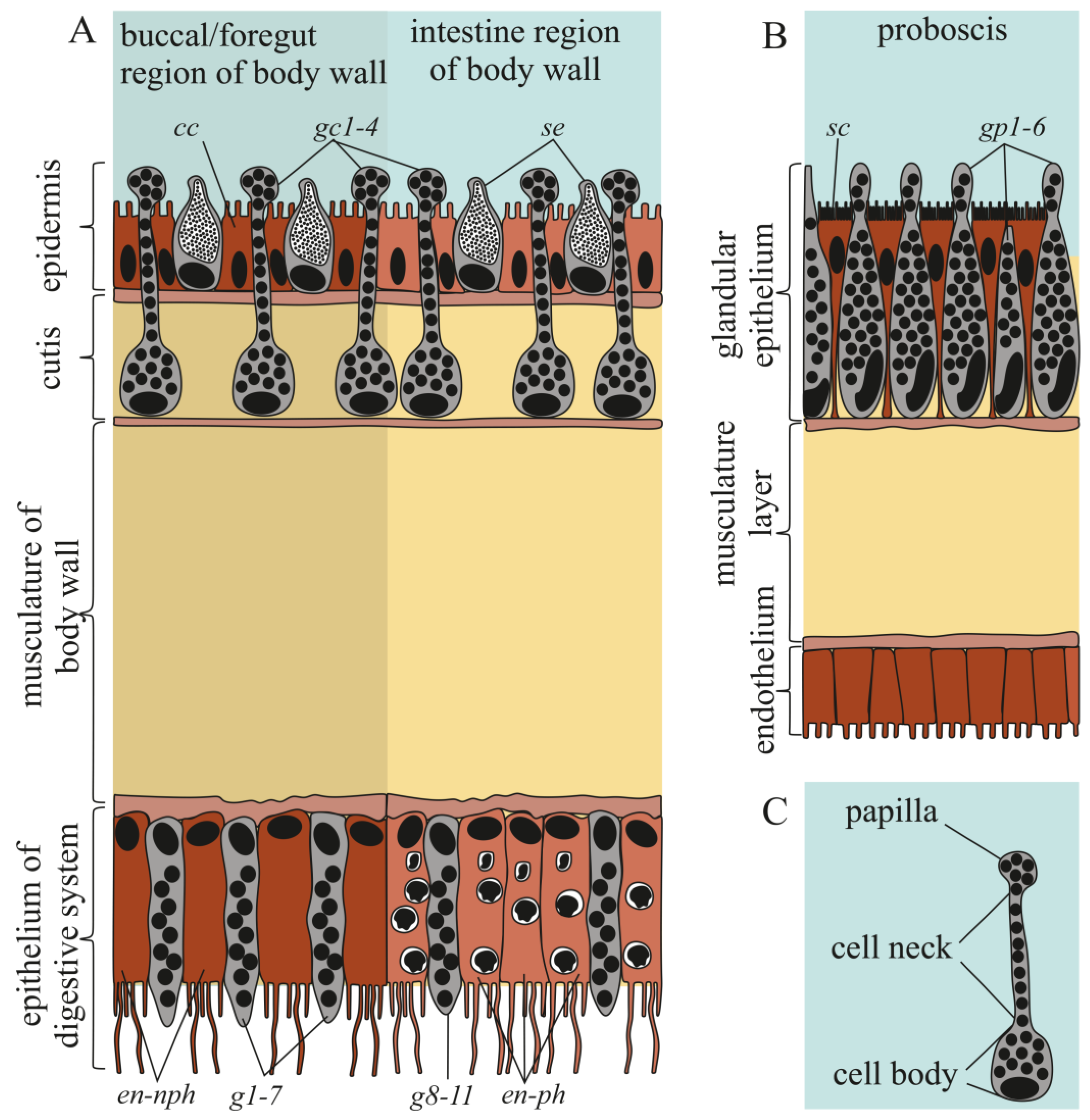
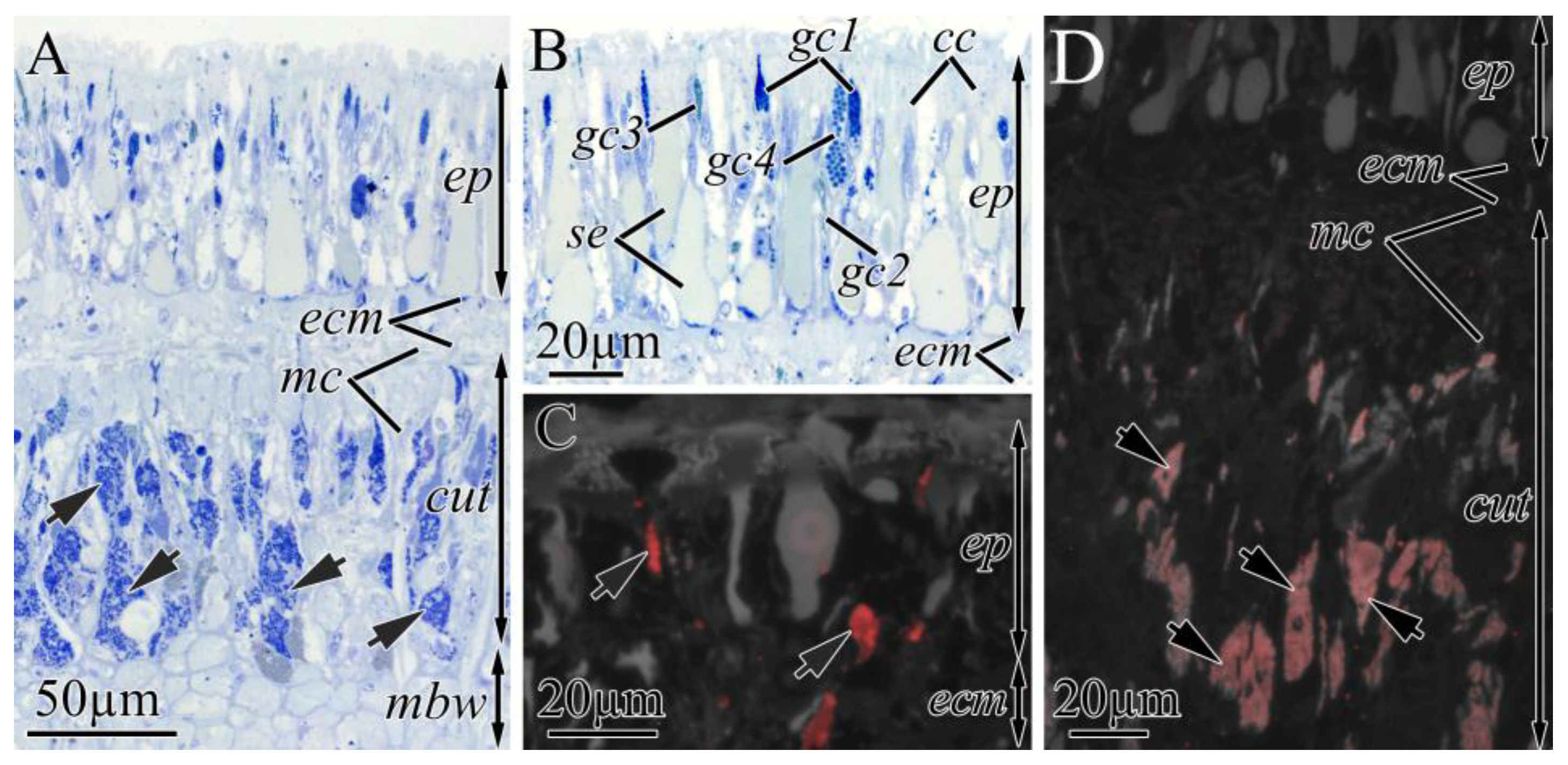
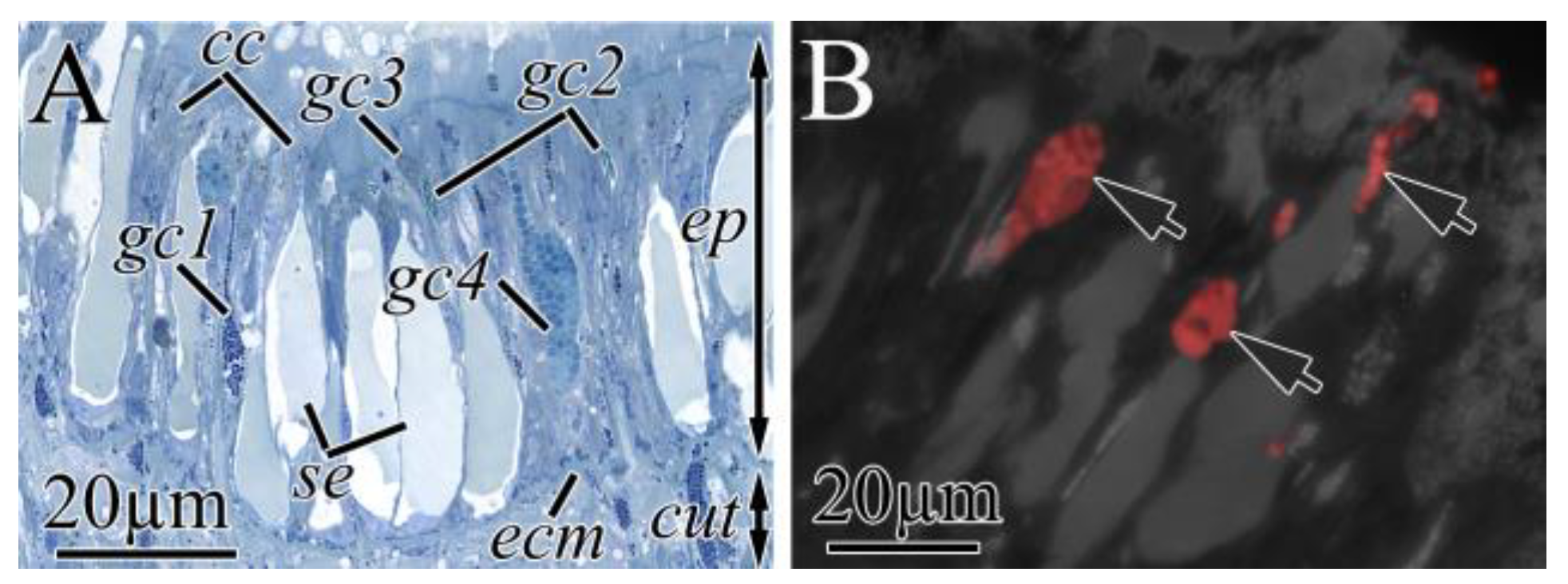
2.2.1. Integument
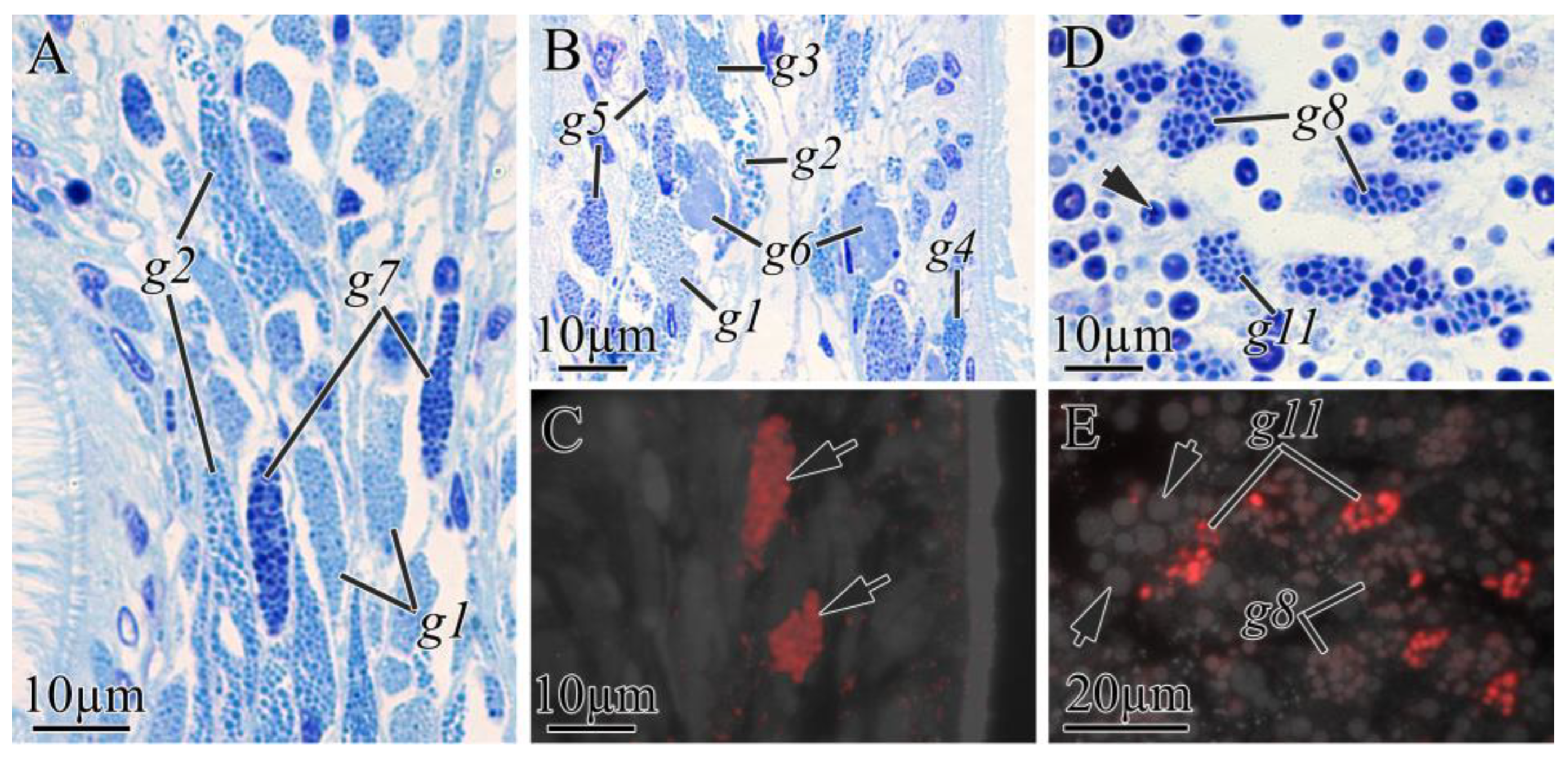
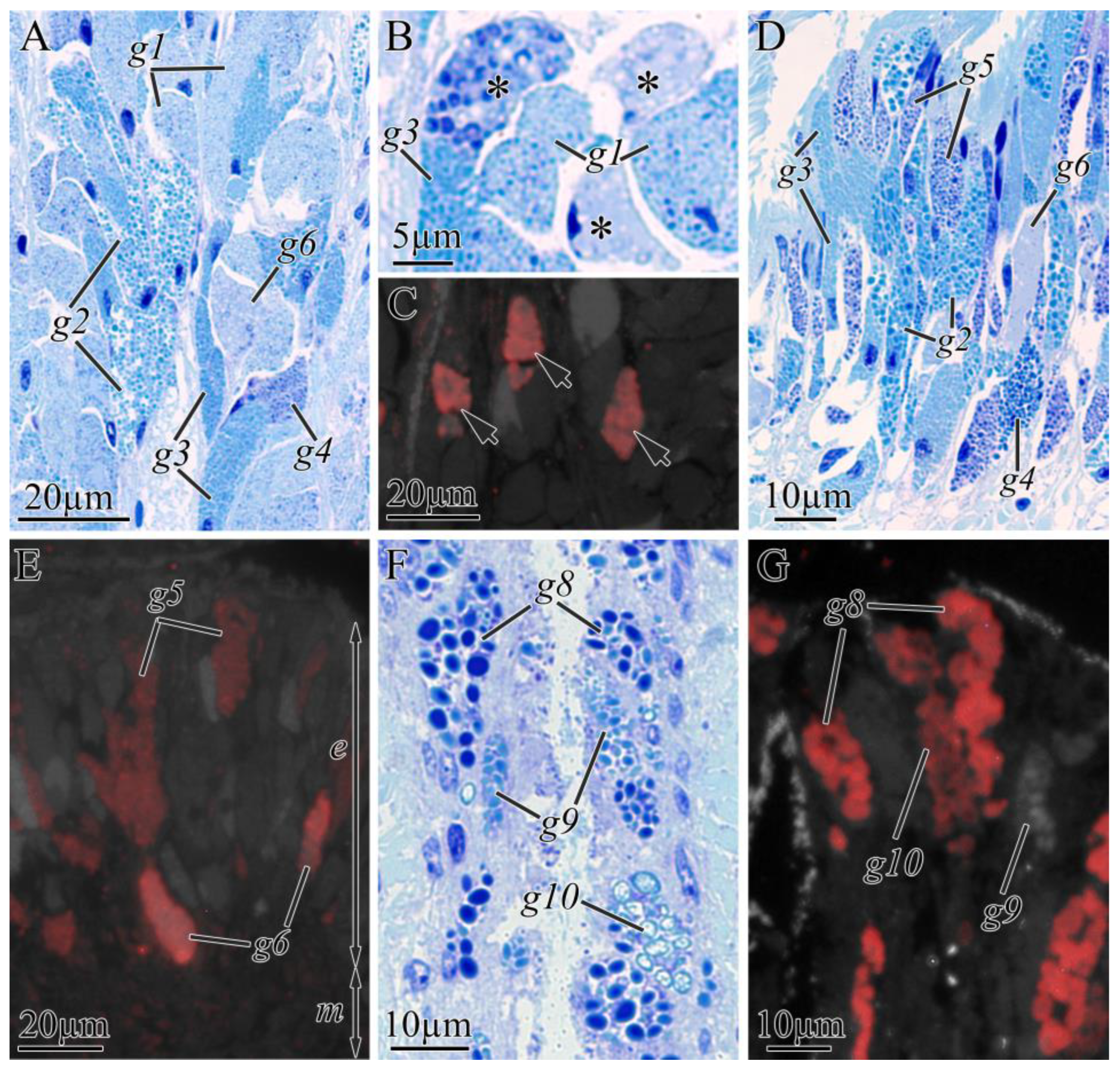
2.2.2. Digestive System
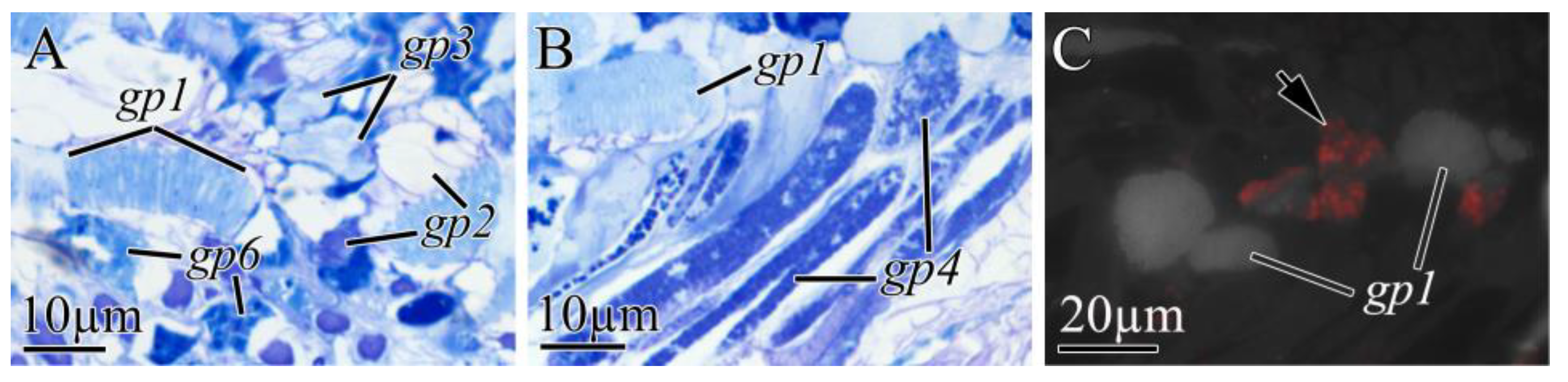
2.2.3. Proboscis
3. Discussion
4. Conclusions
5. Materials and Methods
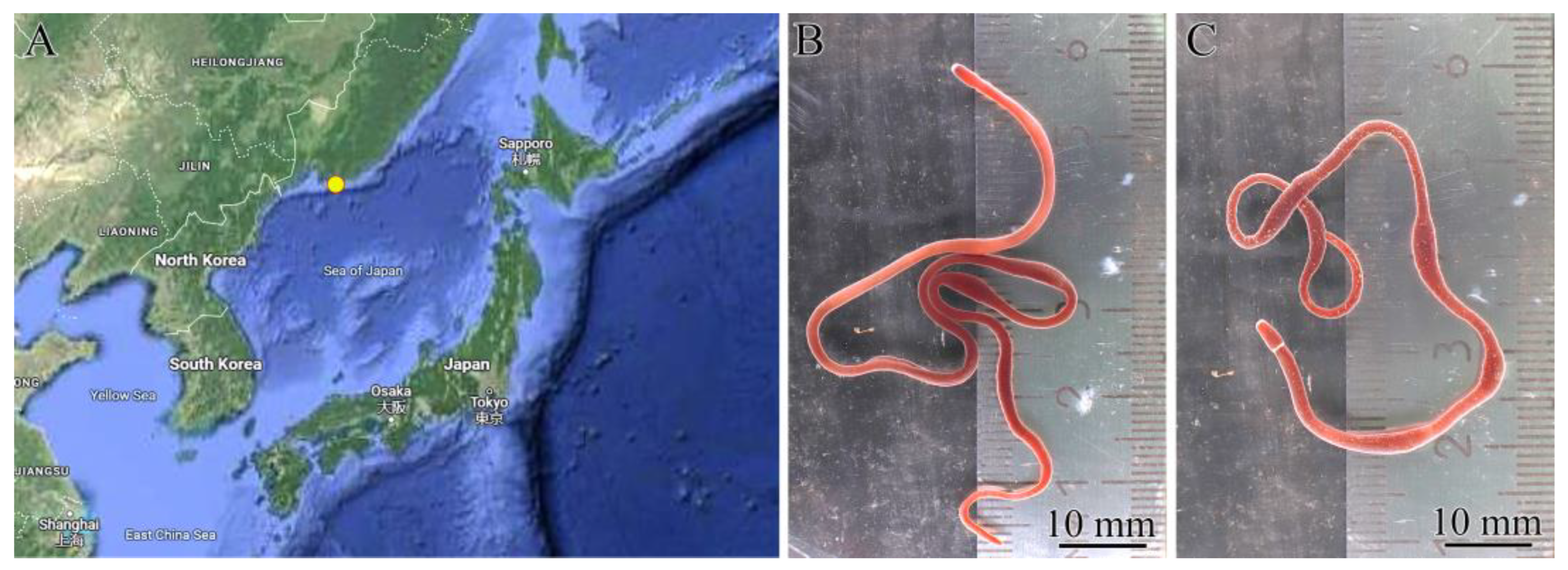
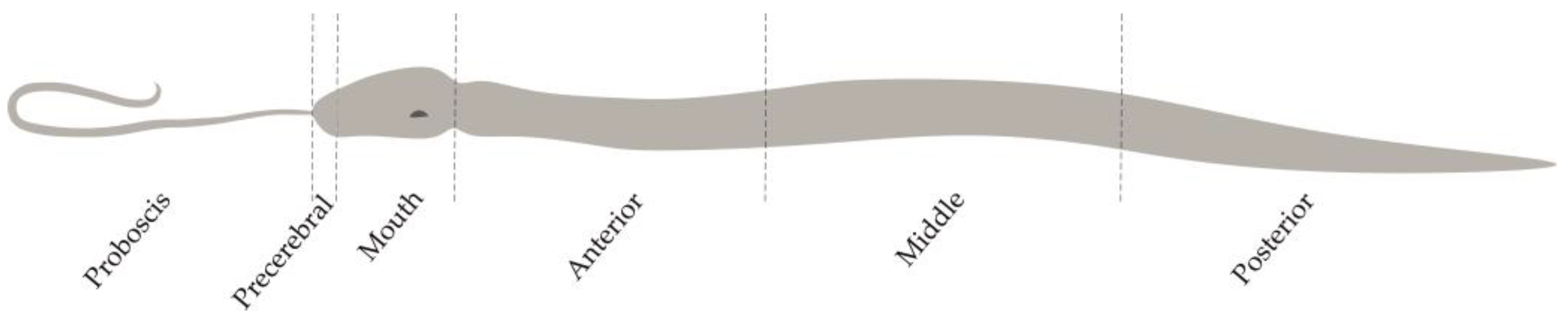
5.1. Sample Collection and Preparation
5.2. Species Identification
5.3. Extraction and HPLC–MS/MS Analysis of TTX and Its Analogues
5.4. Immunohistochemical Studies
5.5. Morphological Studies
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chernyshev, A.V. An updated classification of the phylum Nemertea. Invertebr. Zool. 2021, 18, 188–196. [Google Scholar] [CrossRef]
- Kajihara, H.; Chernyshev, A.V.; Sun, S.; Sundberg, P.; Crandall, F.B. Checklist of nemertean genera and species published between 1995 and 2007. Species Divers. 2008, 13, 245–274. [Google Scholar] [CrossRef]
- Kem, W.R. Purification and characterization of a new family of polypeptide neurotoxins from the heteronemertine Cerebratulus lacteus (Leidy). J. Biol. Chem. 1976, 251, 4184–4192. [Google Scholar] [CrossRef] [PubMed]
- Kem, W.R. Structure and action of nemertine toxins. Integr. Comp. Biol. 1985, 25, 99–111. [Google Scholar] [CrossRef]
- Göransson, U.; Jacobsson, E.; Strand, M.; Andersson, H.S. The toxins of nemertean worms. Toxins 2019, 11, 120. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, M.; Ito, K.; Kajihara, H. Highly toxic ribbon worm Cephalothrix simula containing tetrodotoxin in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxins 2013, 5, 376–395. [Google Scholar] [CrossRef] [PubMed]
- Vlasenko, A.E.; Velansky, P.V.; Chernyshev, A.V.; Kuznetsov, V.G.; Magarlamov, T.Y. Tetrodotoxin and its analogues profile in nemertean species from the Sea of Japan. Toxicon 2018, 156, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O.; Takatani, T. TTX accumulation in pufferfish. Comp. Biochem. Physiol.—Part D Genom. Proteom. 2006, 1, 145–152. [Google Scholar] [CrossRef]
- Asakawa, M.; Toyoshima, T.; Shida, Y.; Noguchi, T.; Miyazawa, K. Paralytic toxins in a ribbon worm Cephalothrix species (Nemertean) adherent to cultured oysters in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxicon 2000, 38, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.; McEvoy, E.G.; Gibson, R. The production of tetrodotoxin-like substances by nemertean worms in conjunction with bacteria. J. Exp. Mar. Bio. Ecol. 2003, 288, 51–63. [Google Scholar] [CrossRef]
- Strand, M.; Hedström, M.; Seth, H.; McEvoy, E.G.; Jacobsson, E.; Göransson, U.; Andersson, H.S.; Sundberg, P. The bacterial (Vibrio alginolyticus) production of tetrodotoxin in the ribbon worm Lineus longissimus—Just a false positive? Mar. Drugs 2016, 14, 63. [Google Scholar] [CrossRef]
- Miyazawa, K.; Higashiyama, M.; Ito, K.; Noguchi, T.; Arakawa, O.; Shida, Y.; Hashimoto, K. Tetrodotoxin in two species of ribbon worm (Nemertini), Lineus fuscoviridis and Tubulanus punctatus. Toxicon 1988, 26, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.S.; Min, S.K.; Yeon, S.J.; Hwang, J.H.; Hong, J.S.; Shin, H.S. Assessment of neuronal cell-based cytotoxicity of neurotoxins from an estuarine nemertean in the han river estuary. J. Microbiol. Biotechnol. 2017, 27, 725–730. [Google Scholar] [CrossRef]
- Ali, A.E.; Arakawa, O.; Noguchi, T.; Miyazawa, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin and related substances in a ribbon worm Cephalothrix linearis (Nemertean). Toxicon 1990, 28, 1083–1093. [Google Scholar] [CrossRef]
- Vlasenko, A.E.; Magarlamov, T.Y. Tetrodotoxin and its analogues in Cephalothrix cf. simula (Nemertea: Palaeonemertea) from the Sea of Japan (Peter the Great Gulf): Intrabody distribution and secretions. Toxins 2020, 12, 745. [Google Scholar] [CrossRef] [PubMed]
- Magarlamov, T.Y.; Shokur, O.A.; Chernyshev, A.V. Distribution of tetrodotoxin in the ribbon worm Lineus alborostratus. Toxicon 2016, 112, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Tanu, M.B.; Mahmud, Y.; Arakawa, O.; Takatani, T.; Kajihara, H.; Kawatsu, K.; Hamano, Y.; Asakawa, M.; Miyazawa, K.; Noguchi, T. Immunoenzymatic visualization of tetrodotoxin (TTX) in Cephalothrix species (Nemertea: Anopla: Palaeonemertea: Cephalotrichidae) and Planocera reticulata (Platyhelminthes: Turbellaria: Polycladida: Planoceridae). Toxicon 2004, 44, 515–520. [Google Scholar] [CrossRef]
- Campbell, M.E.; Schwartz, M. Immunohistological visualization of tetrodotoxin in Micrura verrili and Dushia atra (Phylum Nemertea ). In Proceedings of the National Conferences for Undergraduate Research (NCUR), Salisbury, MD, USA, 10–12 April 2008. [Google Scholar]
- Chen, H.; Strand, M.; Norenburg, J.L.; Sun, S.; Kajihara, H.; Chernyshev, A.V.; Maslakova, S.A.; Sundberg, P. Statistical parsimony networks and species assemblages in cephalotrichid nemerteans (Nemertea). PLoS ONE 2010, 5, e12885. [Google Scholar] [CrossRef]
- Sagorny, C.; Wesseler, C.; Krämer, D.; von Döhren, J. Assessing the diversity and distribution of Cephalothrix species (Nemertea: Palaeonemertea) in European waters by comparing different species delimitation methods. J. Zool. Syst. Evol. Res. 2019, 57, 497–519. [Google Scholar] [CrossRef]
- Kajihara, H.; Sun, S.C.; Chernyshev, A.V.; Chen, H.X.; Ito, K.; Asakawa, M.; Maslakova, S.A.; Norenburg, J.L.; Strand, M.; Sundberg, P.; et al. Taxonomic identity of a tetrodotoxin-accumulating ribbon-worm Cephalothrix simula (Nemertea: Palaeonemertea): A species artificially introduced from the pacific to Europe. Zoolog. Sci. 2013, 30, 985–997. [Google Scholar] [CrossRef]
- Kajihara, H. Resolving a 200-year-old taxonomic conundrum: Neotype designation for Cephalothrix linearis (nemertea: Palaeonemertea) based on a topotype from Bergen, Norway. Fauna Nor. 2019, 39, 39–76. [Google Scholar] [CrossRef]
- Asakawa, M.; Toyoshima, T.; Ito, K.; Bessho, K.; Yamaguchi, C.; Tsunetsugu, S.; Shida, Y.; Kajihara, H.; Mawatari, S.F.; Noguchi, T.; et al. Paralytic toxicity in the ribbon worm Cephalothrix species (Nemertea) in Hiroshima Bay, Hiroshima Prefecture, Japan and the isolation of tetrodotoxin as a main component of its toxins. Toxicon 2003, 41, 747–753. [Google Scholar] [CrossRef]
- Turner, A.D.; Fenwick, D.; Powell, A.; Dhanji-Rapkova, M.; Ford, C.; Hatfield, R.G.; Santos, A.; Martinez-Urtaza, J.; Bean, T.P.; Baker-Austin, C.; et al. New invasive nemertean species (Cephalothrix simula) in England with high levels of tetrodotoxin and a microbiome linked to toxin metabolism. Mar. Drugs 2018, 16, 452. [Google Scholar] [CrossRef] [PubMed]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [PubMed]
- Malykin, G.V.; Chernyshev, A.V.; Magarlamov, T.Y. Intrabody tetrodotoxin distribution and possible hypothesis for its migration in ribbon worms Cephalothrix cf. simula (Palaeonemertea, Nemertea). Mar. Drugs 2021, 19, 494. [Google Scholar] [CrossRef] [PubMed]
- Chernyshev, A.V. Nemerteans from the Far Eastern Seas of Russia. Russ. J. Mar. Biol. 2020, 46, 141–153. [Google Scholar] [CrossRef]
- Chernyshev, A.V.; Polyakova, N.E.; Turanov, S.V.; Kajihara, H. Taxonomy and phylogeny of Lineus torquatus and allies (Nemertea, Lineidae) with descriptions of a new genus and a new cryptic species. Syst. Biodivers. 2018, 16, 55–68. [Google Scholar] [CrossRef]
- Jennings, J.B.; Gibson, R. Observations on the nutrition of seven species of rthynchocoelan worms. Biol. Bull. 1969, 136, 405–433. [Google Scholar] [CrossRef]
- Gibson, R. The physiology of digestion in Nemertean worms. Bol. Zool. Biol. Mar. 1972, 29, 55–64. [Google Scholar] [CrossRef]
- Gibson, R. Nemertea. In Synopsis and Classification of Living Organisms; Parker, S.P., Ed.; McGraw-Hill: New York, NY, USA, 1982; pp. 823–846. [Google Scholar]
- Chernyshev, A.V. Comparative Morphology, Systematics and Phylogeny of the Nemerteans; Dalnauka: Vladivostok, Russia, 2011. [Google Scholar]
- Williams, B.L. Behavioral and chemical ecology of marine organisms with respect to tetrodotoxin. Mar. Drugs 2010, 8, 381–398. [Google Scholar] [CrossRef]
- Vlasenko, A.E.; Kuznetsov, V.G.; Malykin, G.V.; Pereverzeva, A.O.; Velansky, P.V.; Yakovlev, K.V.; Magarlamov, T.Y. Tetrodotoxins secretion and voltage-gated sodium channel adaptation in the ribbon worm Kulikovia alborostrata (Takakura, 1898) (Nemertea). Toxins 2021, 13, 606. [Google Scholar] [CrossRef]
- Magarlamov, T.Y.; Turbeville, J.M.; Chernyshev, A.V. Pseudocnidae of ribbon worms (Nemertea): Ultrastructure, maturation, and functional morphology. PeerJ 2021, 9, e10912. [Google Scholar] [CrossRef]
- Malykin, G.V.; Velansky, P.V.; Magarlamov, T.Y. Tetrodotoxin and Its Analogues (TTXs) in the Food-Capture and Defense Organs of the Palaeonemertean Cephalothrix cf. simula. Toxins 2024, 16, 43. [Google Scholar] [CrossRef] [PubMed]
- Bürger, O. Die Nemertinen des Golfes von Neapel und der Angrenzenden Meeres–Abschnitte. Fauna und Flora des Golfes von Neapel; Verlag von R. Friedländer & Sohn: Berlin, Germany, 1895; Volume 22. [Google Scholar]
- Martin, C.H. A note on the occurrence of nematocysts and similar structures in the various groups of the animal kingdom. Biol. Zentrablatt 1914, 34, 248–273. [Google Scholar]
- Gontcharoff, M. Étude des rhabdites de la trompe de Lineus ruber (Némertien) au microscope électronique. Compte Rendu Hebd. Des Séances L’académie Des Sci. Paris 1957, 244, 1539–1541. [Google Scholar]
- Hyman, L.H. The Invertebrates. Vol. V: Smaller Coelomate Groups. Chaetognatha, Hemichordata, Pogonophora, Phoronida, Ectoprocta, Brachiopoda, Sipunculida. The Coelomate Bilateria; McGraw-Hill: New York, NY, USA, 1959. [Google Scholar]
- Ling, E.A. The proboscis apparatus of the nemertine Lineus ruber. Philos. Trans. R. Soc. Lond. B 1971, 262, 1–22. [Google Scholar]
- Whitelaw, B.L.; Strugnell, J.M.; Faou, P.; Da Fonseca, R.R.; Hall, N.E.; Norman, M.; Finn, J.; Cooke, I.R. Combined transcriptomic and proteomic analysis of the posterior salivary gland from the southern blue-ringed octopus and the southern sand octopus. J. Proteome Res. 2016, 15, 3284–3297. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Sugimoto, A.; Takai, A.; Yasumoto, T. Effects of specific modifications of several hydroxyls of tetrodotoxin on its affinity to rat brain membrane. J. Pharmacol. Exp. Ther. 1999, 289, 1688–1696. [Google Scholar]
- Noguchi, Y.; Suzuki, T.; Matsutani, K.; Sakakibara, R.; Nakahigashi, R.; Adachi, M.; Nishikawa, T.; Abe, H. An almost nontoxic tetrodotoxin analog, 5,6,11-trideoxytetrodotoxin, as an odorant for the grass puffer. Sci. Rep. 2022, 12, 15087. [Google Scholar] [CrossRef]
- Puilingi, C.G.; Kudo, Y.; Cho, Y.; Konoki, K.; Yotsu-Yamashita, M. Tetrodotoxin and its analogues in the pufferfish Arothron hispidus and A. nigropunctatus from the Solomon Islands: A comparison of their toxin profiles with the same species from Okinawa, Japan. Toxins 2015, 7, 3436–3454. [Google Scholar] [CrossRef]
- von Reumont, B.M.; Lüddecke, T.; Timm, T.; Lochnit, G.; Vilcinskas, A.; von Döhren, J.; Nilsson, M.A. Proteo-transcriptomic analysis identifies potential novel toxins secreted by the predatory, orey-piercing ribbon worm Amphiporus lactifloreus. Mar. Drugs 2020, 18, 407. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.T.J.; Calhoun, D.M.; Stokes, A.N.; Susbilla, C.B.; McDevitt-Galles, T.; Briggs, C.J.; Hoverman, J.T.; Tkach, V.V.; de Roode, J.C. Of poisons and parasites—The defensive role of tetrodotoxin against infections in newts. J. Anim. Ecol. 2018, 87, 1192–1204. [Google Scholar] [CrossRef]
- Chernyshev, A.V.; Polyakova, N.E. Nemerteans collected in the Bering Sea during the research cruises aboard the R/V Akademik M.A. Lavrentyev in 2016, 2018, and 2021 with an analysis of deep-sea heteronemertean and hoplonemertean species. Deep. Res. Part II Top. Stud. Oceanogr. 2022, 199, 105081. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Chen, X.W.; Liu, H.X.; Jin, Y.B.; Li, S.F.; Bi, X.; Chung, S.; Zhang, S.S.; Jiang, Y.Y. Separation, identification and quantification of tetrodotoxin and its analogs by LC-MS without calibration of individual analogs. Toxicon 2011, 57, 938–943. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Kawaura, R.; Togashi, K.; Mizusawa, N.; Yasumoto, K.; Takada, K.; Amano, M.; Watabe, S. De novo accumulation of tetrodotoxin and its analogs in pufferfish and newt and dosage-driven accumulation of toxins in newt: Tissue distribution and anatomical localization. J. Mar. Sci. Eng. 2021, 9, 1004. [Google Scholar] [CrossRef]
- Sato, S.; Takaishi, S.; Yasumoto, K.; Watabe, S. Novel polyclonal antibody raised against tetrodotoxin using its haptenic antigen prepared from 4,9-anhydrotetrodotoxin reacted with 1,2-Eethaneditiol and further reacted with keyhole limpet hemocyanin. Toxins 2019, 11, 551. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
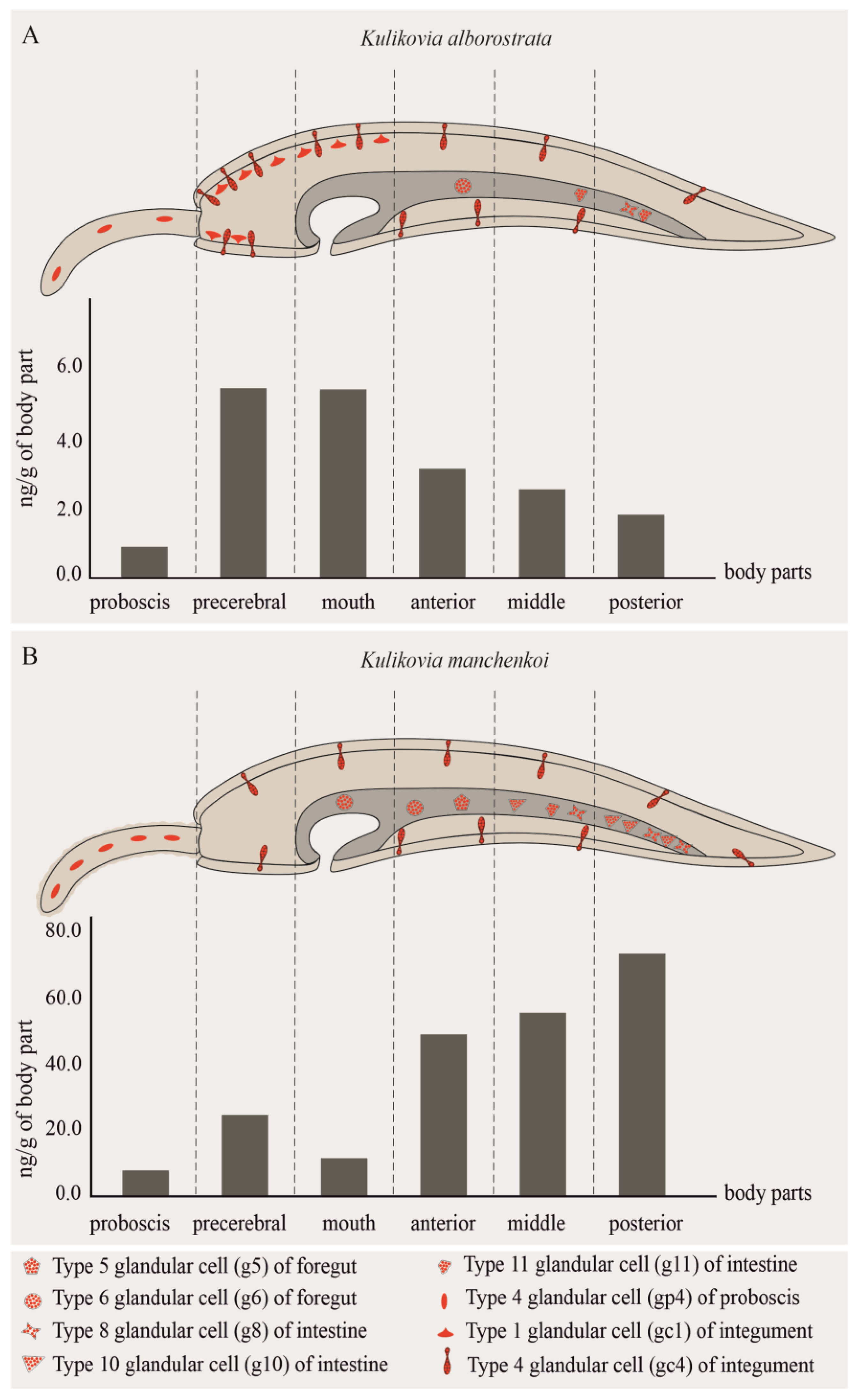
| Body Region | Weight, g | 5,6,11-TrideoxyTTX | TTX | 5-DeoxyTTX | ||
|---|---|---|---|---|---|---|
| ng/g | ng | % * | ||||
| Proboscis | 0.39 | 0.91 | 0.36 | 3.81 | − | − |
| Precerebral | 0.04 | 5.52 | 0.20 | 2.19 | − | − |
| Mouth | 0.26 | 5.51 | 1.43 | 15.33 | − | − |
| Anterior body | 0.98 | 3.19 | 3.13 | 33.46 | − | − |
| Middle body | 1.10 | 2.59 | 2.84 | 30.45 | + | − |
| Posterior body | 0.75 | 1.84 | 1.38 | 14.76 | − | − |
| Body Region | Weight, g | 5,6,11-TrideoxyTTX | TTX | 5-DeoxyTTX | ||
|---|---|---|---|---|---|---|
| ng/g | ng | % * | ||||
| Proboscis | 0.30 | 7.47 | 2.24 | 2.07 | − | − |
| Precerebral | 0.06 | 24.06 | 1.44 | 1.33 | − | − |
| Mouth | 0.39 | 11.27 | 4.39 | 4.06 | − | − |
| Anterior body | 0.76 | 48.16 | 36.60 | 33.82 | − | + |
| Middle body | 0.59 | 54.99 | 32.44 | 29.98 | − | + |
| Posterior body | 0.43 | 72.32 | 31.10 | 28.74 | − | + |
| Cell Type | Morphological Description | Methylene Blue Staining | Kulikovia alborostrata | Kulikovia manchenkoi | ||||
|---|---|---|---|---|---|---|---|---|
| Localization | Distribution | TTXs-Positive Immunoreactivity | Localization | Distribution | TTXs-Positive Immunoreactivity | |||
| se | single large secretory granule | light blue | epidermis | +++ | − | epidermis | +++ | − |
| gc1 | small rounded secretory granules | blue-purple | cutis | +++ | weak | cutis | +++ | − |
| gc2 | elongated secretory granules | blue-green | cutis | ++ | − | cutis | ++ | − |
| gc3 | small rounded secretory granules | blue-green | cutis | + | − | cutis | + | − |
| gc4 | large spherical secretory granules | blue | cutis | + | high | cutis | + | high |
| g1 | small rounded secretory granules | light blue | buccal cavity, foregut | +++ | − | buccal cavity | +++ | − |
| g2 | large rounded secretory granules | blue | buccal cavity, foregut | ++ | − | buccal cavity, foregut | ++ | − |
| g3 | rounded or oval secretory granules | blue | foregut | ++ | − | buccal cavity, foregut | ++ | − |
| g4 | rounded or oval secretory granules | purple-blue | foregut | + | − | buccal cavity, foregut | + | − |
| g5 | rounded secretory granules | purple | foregut | +++ | − | foregut | +++ | medium |
| g6 | large spherical secretory granules with heterogeneous contents | light purple | foregut | + | high | buccal cavity, foregut | + | high |
| g7 | large rounded secretory granules | dark blue | buccal cavity | + | − | − | − | − |
| g8 | large rounded secretory granules | blue-purple | intestine | +++ | weak | intestine | +++ | high |
| g9 | rounded or oval secretory granules | light blue | − | − | − | intestine | ++ | − |
| g10 | large rounded secretory granules with heterogeneous contents | light blue | − | − | − | intestine | ++ | high |
| g11 | rounded secretory granules | blue-purple | intestine | ++ | high | − | − | − |
| gp1 | pseudocnidae | light blue | proboscis | +++ | − | proboscis | +++ | − |
| gp2 | large bacillary secretory granules | purple or light purple | proboscis | +++ | − | proboscis | +++ | − |
| gp3 | large spherical secretory granules | blue or light blue | proboscis | ++ | − | proboscis | ++ | − |
| gp4 | small rounded secretory granules | dark blue | proboscis | + | high | proboscis | ++ | high |
| gp5 | large spherical secretory granules with heterogeneous content | purple | − | − | − | proboscis | ++ | − |
| gp6 | rounded secretory granules with heterogeneous content | blue | proboscis | ++ | − | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vlasenko, A.E.; Pereverzeva, A.O.; Velansky, P.V.; Magarlamov, T.Y. Tetrodotoxins in Tissues and Cells of Different Body Regions of Ribbon Worms Kulikovia alborostrata and K. manchenkoi from Spokoynaya Bay, Sea of Japan. Toxins 2024, 16, 186. https://doi.org/10.3390/toxins16040186
Vlasenko AE, Pereverzeva AO, Velansky PV, Magarlamov TY. Tetrodotoxins in Tissues and Cells of Different Body Regions of Ribbon Worms Kulikovia alborostrata and K. manchenkoi from Spokoynaya Bay, Sea of Japan. Toxins. 2024; 16(4):186. https://doi.org/10.3390/toxins16040186
Chicago/Turabian StyleVlasenko, Anna E., Alexandra O. Pereverzeva, Peter V. Velansky, and Timur Yu. Magarlamov. 2024. "Tetrodotoxins in Tissues and Cells of Different Body Regions of Ribbon Worms Kulikovia alborostrata and K. manchenkoi from Spokoynaya Bay, Sea of Japan" Toxins 16, no. 4: 186. https://doi.org/10.3390/toxins16040186
APA StyleVlasenko, A. E., Pereverzeva, A. O., Velansky, P. V., & Magarlamov, T. Y. (2024). Tetrodotoxins in Tissues and Cells of Different Body Regions of Ribbon Worms Kulikovia alborostrata and K. manchenkoi from Spokoynaya Bay, Sea of Japan. Toxins, 16(4), 186. https://doi.org/10.3390/toxins16040186