Microbial Community Response to Granular Peroxide-Based Algaecide Treatment of a Cyanobacterial Harmful Algal Bloom in Lake Okeechobee, Florida (USA)

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
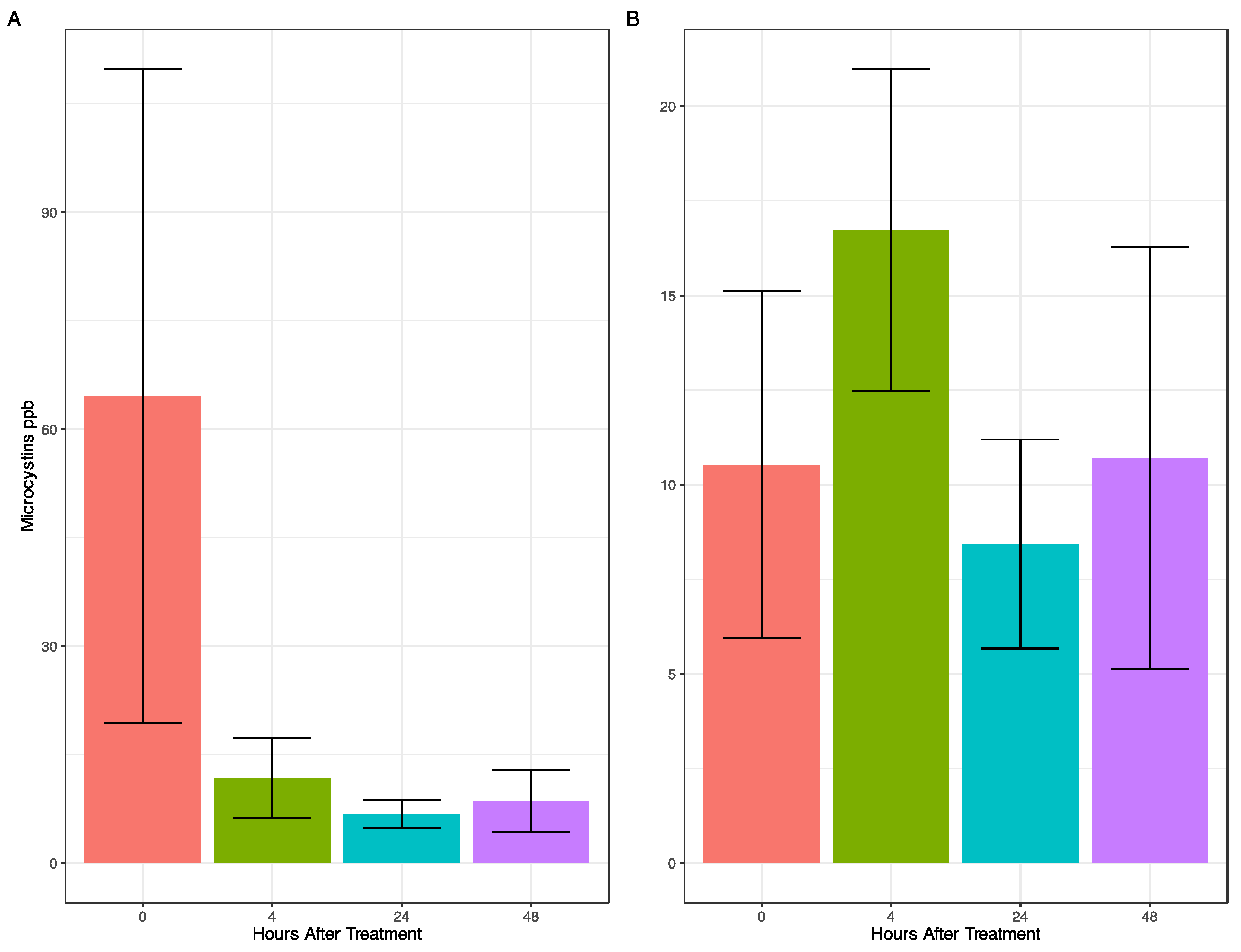
2.1. Effects of Peroxide-Based Algaecide on Cyanobacterial Abundance and Toxins
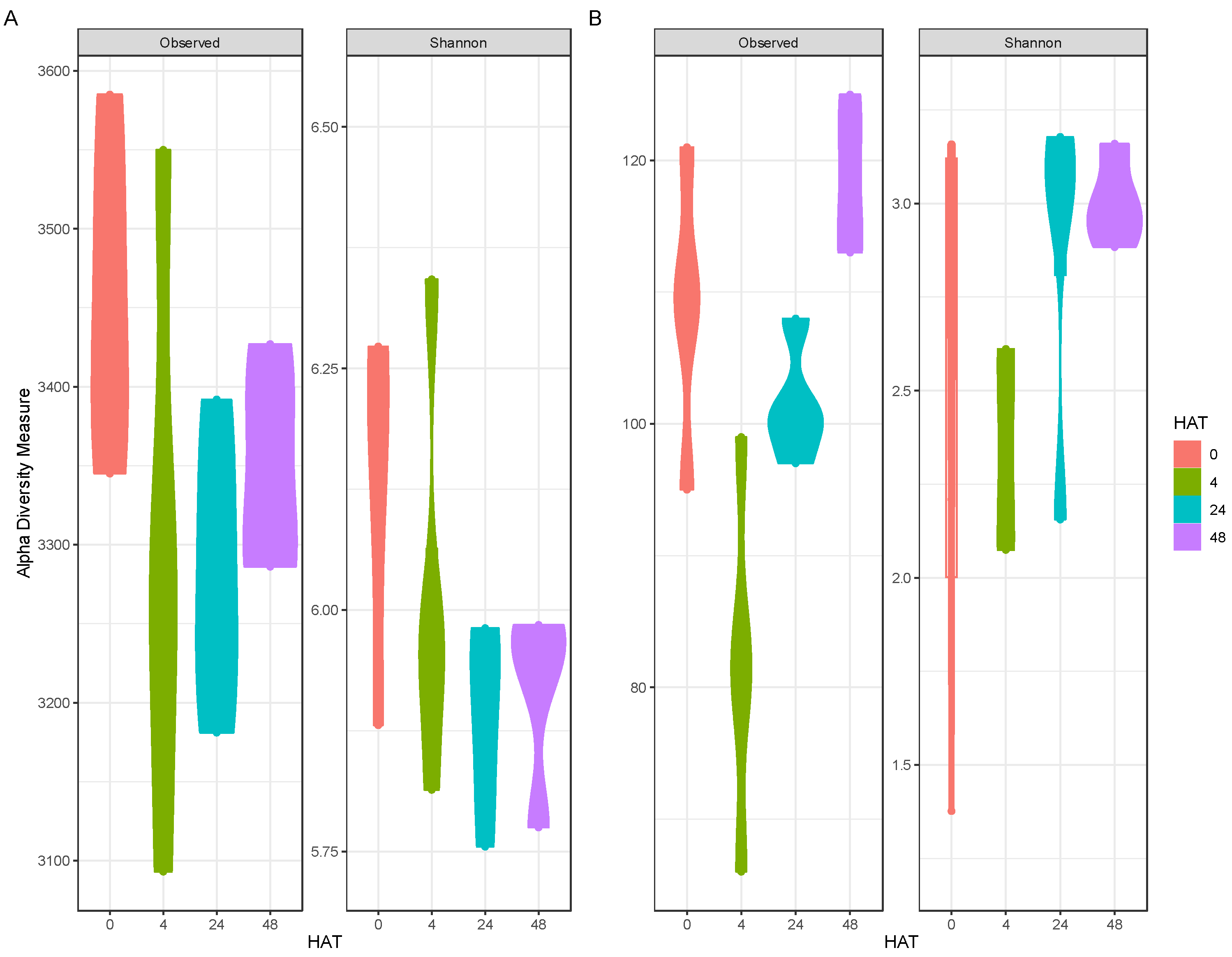
2.2. Community Composition
3. Conclusions
4. Materials and Methods
4.1. Study Site, Peroxide Treatment, Sampling Information
4.2. Microcystin Analysis
4.3. Pigment Analysis
4.4. Phytoplankton Counts
4.5. DNA Extraction, 16S rRNA and 18S rRNA Amplification and Sequencing
4.6. Sequencing Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeppesen, E.; Søndergaard, M.; Meerhoff, M.; Lauridsen, T.L.; Jensen, J.P. Shallow Lake Restoration by Nutrient Loading Reduction—Some Recent Findings and Challenges Ahead. Hydrobiologia 2007, 584, 239–252. [Google Scholar] [CrossRef]
- Huang, I.-S.; Zimba, P.V. Cyanobacterial Bioactive Metabolites—A Review of Their Chemistry and Biology. Harmful Algae 2019, 86, 139–209. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Berthold, D.E.; Yu, Z.; Silva-Sanchez, C.; Laughinghouse IV, H.D.; Denslow, N.D.; Han, S. Atmospheric Progression of Microcystin-LR from Cyanobacterial Aerosols. Environ. Sci. Technol. Lett. 2020, 7, 740–745. [Google Scholar] [CrossRef]
- Moretto, J.A.; Freitas, P.N.N.; Souza, J.P.; Oliveira, T.M.; Brites, I.; Pinto, E. Off-Flavors in Aquacultured Fish: Origins and Implications for Consumers. Fishes 2022, 7, 34. [Google Scholar] [CrossRef]
- Werner, V.R.; Laughinghouse, H.D.I. Bloom-Forming and Other Planktonic Anabaena (Cyanobacteria) Morphospecies with Twisted Trichomes from Rio Grande Do Sul State, Brazil. Nova Hedwig. 2009, 89, 17–47. [Google Scholar] [CrossRef]
- Almanza, V.; Pedreros, P.; Dail Laughinghouse, H.; Félez, J.; Parra, O.; Azócar, M.; Urrutia, R. Association between Trophic State, Watershed Use, and Blooms of Cyanobacteria in South-Central Chile. Limnologica 2019, 75, 30–41. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A Review of the Global Ecology, Genomics, and Biogeography of the Toxic Cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.C.; Michalak, A.M.; Pahlevan, N. Widespread Global Increase in Intense Lake Phytoplankton Blooms since the 1980s. Nature 2019, 574, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of Knowledge and Concerns on Cyanobacterial Blooms and Cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms Like It Hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef]
- Lefler, F.W.; Barbosa, M.; Zimba, P.V.; Smyth, A.R.; Berthold, D.E.; Laughinghouse, H.D. Spatiotemporal Diversity and Community Structure of Cyanobacteria and Associated Bacteria in the Large Shallow Subtropical Lake Okeechobee (Florida, United States). Front. Microbiol. 2023, 14, 1219261. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.; Newman, R.S.; Osborne, T.Z.; White, J.R.; Fitz, H.C. Phosphorous Cycling in the Greater Everglades Ecosystem: Legacy Phosphorous Implications for Management and Restoration. Crit. Rev. Environ. Sci. Technol. 2011, 41, 149–186. [Google Scholar] [CrossRef]
- Jarvie, H.P.; Sharpley, A.N.; Spears, B.; Buda, A.R.; May, L.; Kleinman, P.J. Water quality remediation faces unprecedented challenges from “legacy phosphorus”. Environ. Sci. Technol. 2013, 47, 8997–8998. [Google Scholar] [CrossRef]
- Matthijs, H.C.P.; Visser, P.M.; Reeze, B.; Meeuse, J.; Slot, P.C.; Wijn, G.; Talens, R.; Huisman, J. Selective Suppression of Harmful Cyanobacteria in an Entire Lake with Hydrogen Peroxide. Water Res. 2012, 46, 1460–1472. [Google Scholar] [CrossRef] [PubMed]
- Kinley-Baird, C.; Calomeni, A.; Berthold, D.E.; Lefler, F.W.; Barbosa, M.; Rodgers, J.H.; Laughinghouse, H.D. Laboratory-Scale Evaluation of Algaecide Effectiveness for Control of Microcystin-Producing Cyanobacteria from Lake Okeechobee, Florida (USA). Ecotoxicol. Environ. Saf. 2021, 207, 111233. [Google Scholar] [CrossRef] [PubMed]
- Pokrzywinski, K.L.; Bishop, W.M.; Grasso, C.R.; Fernando, B.M.; Sperry, B.P.; Berthold, D.E.; Laughinghouse, H.D.; Van Goethem, E.M.; Volk, K.; Heilman, M.; et al. Evaluation of a Peroxide-Based Algaecide for Cyanobacteria Control: A Mesocosm Trial in Lake Okeechobee, FL, USA. Water 2022, 14, 169. [Google Scholar] [CrossRef]
- Santos, A.A.; Guedes, D.O.; Barros, M.U.G.; Oliveira, S.; Pacheco, A.B.F.; Azevedo, S.M.F.O.; Magalhães, V.F.; Pestana, C.J.; Edwards, C.; Lawton, L.A.; et al. Effect of Hydrogen Peroxide on Natural Phytoplankton and Bacterioplankton in a Drinking Water Reservoir: Mesocosm-Scale Study. Water Res. 2021, 197, 117069. [Google Scholar] [CrossRef] [PubMed]
- Lefler, F.W.; Berthold, D.E.; Barbosa, M.; Laughinghouse, H.D. The Effects of Algaecides and Herbicides on a Nuisance Microcystis wesenbergii-Dominated Bloom. Water 2022, 14, 1739. [Google Scholar] [CrossRef]
- Laughinghouse, H.D., IV; Berthold, D.E.; Bishop, W.B. Approaches to managing cyanobacterial blooms and altering water quality. Aquatics 2020, 42, 13–16. [Google Scholar]
- Reichwaldt, E.S.; Zheng, L.; Barrington, D.J.; Ghadouani, A. Acute Toxicological Response of Daphnia and Moina to Hydrogen Peroxide. J. Environ. Eng. 2012, 138, 607–611. [Google Scholar] [CrossRef]
- Piel, T.; Sandrini, G.; Weenink, E.F.J.; Qin, H.; Herk, M.J.V.; Morales-Grooters, M.L.; Schuurmans, J.M.; Slot, P.C.; Wijn, G.; Arntz, J.; et al. Shifts in Phytoplankton and Zooplankton Communities in Three Cyanobacteria-Dominated Lakes after Treatment with Hydrogen Peroxide. Harmful Algae 2024, 133, 102585. [Google Scholar] [CrossRef] [PubMed]
- Weenink, E.F.J.; Kraak, M.H.S.; Van Teulingen, C.; Kuijt, S.; Van Herk, M.J.; Sigon, C.A.M.; Piel, T.; Sandrini, G.; Leon-Grooters, M.; De Baat, M.L.; et al. Sensitivity of Phytoplankton, Zooplankton and Macroinvertebrates to Hydrogen Peroxide Treatments of Cyanobacterial Blooms. Water Res. 2022, 225, 119169. [Google Scholar] [CrossRef] [PubMed]
- Bauzá, L.; Aguilera, A.; Echenique, R.; Andrinolo, D.; Giannuzzi, L. Application of Hydrogen Peroxide to the Control of Eutrophic Lake Systems in Laboratory Assays. Toxins 2014, 6, 2657–2675. [Google Scholar] [CrossRef] [PubMed]
- Zilliges, Y.; Kehr, J.C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Börner, T.; Dittmann, E. The Cyanobacterial Hepatotoxin Microcystin Binds to Proteins and Increases the Fitness of Microcystis under Oxidative Stress Conditions. PLoS ONE 2011, 6, e17615. [Google Scholar] [CrossRef]
- Barrington, D.J.; Ghadouani, A.; Ivey, G.N. Cyanobacterial and Microcystins Dynamics Following the Application of Hydrogen Peroxide to Waste Stabilisation Ponds. Hydrol. Earth Syst. Sci. 2013, 17, 2097–2105. [Google Scholar] [CrossRef]
- Weenink, E.F.J.; Matthijs, H.C.P.; Schuurmans, J.M.; Piel, T.; Van Herk, M.J.; Sigon, C.A.M.; Visser, P.M.; Huisman, J. Interspecific Protection against Oxidative Stress: Green Algae Protect Harmful Cyanobacteria against Hydrogen Peroxide. Environ. Microbiol. 2021, 23, 2404–2419. [Google Scholar] [CrossRef]
- Lusty, M.W.; Gobler, C.J. The Efficacy of Hydrogen Peroxide in Mitigating Cyanobacterial Blooms and Altering Microbial Communities across Four Lakes in NY, USA. Toxins 2020, 12, 428. [Google Scholar] [CrossRef] [PubMed]
- Piel, T.; Sandrini, G.; Muyzer, G.; Brussaard, C.P.D.; Slot, P.C.; Van Herk, M.J.; Huisman, J.; Visser, P.M. Resilience of Microbial Communities after Hydrogen Peroxide Treatment of a Eutrophic Lake to Suppress Harmful Cyanobacterial Blooms. Microorganisms 2021, 9, 1495. [Google Scholar] [CrossRef]
- Smith, D.J.; Berry, M.A.; Cory, R.M.; Johengen, T.H.; Kling, G.W.; Davis, T.W.; Dick, G.J. Heterotrophic Bacteria Dominate Catalase Expression during Microcystis Blooms. Appl. Environ. Microbiol. 2022, 88, e02544-21. [Google Scholar] [CrossRef]
- Kim, M.; Shin, B.; Lee, J.; Park, H.Y.; Park, W. Culture-independent and culture-dependent analyses of the bacterial community in the phycosphere of cyanobloom-forming Microcystis aeruginosa. Sci. Rep. 2019, 9, 20416. [Google Scholar] [CrossRef]
- Schuurmans, J.M.; Brinkmann, B.W.; Makower, A.K.; Dittmann, E.; Huisman, J.; Matthijs, H.C.P. Microcystin interferes with defense against oxidative stress in harmful cyanobacteria. Harmful Algae 2018, 78, 47–55. [Google Scholar] [CrossRef]
- Rositano, J.; Nicholson, B.C.; Pieronne, P. Destruction of Cyanobacterial Toxins By Ozone. Ozone Sci. Eng. 1998, 20, 223–238. [Google Scholar] [CrossRef]
- Cornish, B.J.P.A.; Lawton, L.A.; Robertson, P.K.J. Hydrogen Peroxide Enhanced Photocatalytic Oxidation of Microcystin-LR Using Titanium Dioxide. Appl. Catal. B Environ. 2000, 25, 59–67. [Google Scholar] [CrossRef]
- Lürling, M.; Meng, D.; Faassen, E. Effects of Hydrogen Peroxide and Ultrasound on Biomass Reduction and Toxin Release in the Cyanobacterium, Microcystis Aeruginosa. Toxins 2014, 6, 3260–3280. [Google Scholar] [CrossRef]
- Spoof, L.; Jaakkola, S.; Važić, T.; Häggqvist, K.; Kirkkala, T.; Ventelä, A.M.; Kirkkala, T.; Svirčev, Z.; Meriluoto, J. Elimination of cyanobacteria and microcystins in irrigation water-effects of hydrogen peroxide treatment. Environ. Sci. Pollut. Res. Int. 2020, 27, 8638–8652. [Google Scholar] [CrossRef]
- Jones, G. Release and Degradation of Microcystin Following Algicide Treatment of a Microcystis Aeruginosa Bloom in a Recreational Lake, as Determined by HPLC and Protein Phosphatase Inhibition Assay. Water Res. 1994, 28, 871–876. [Google Scholar] [CrossRef]
- Cardman, Z.; Arnosti, C.; Durbin, A.; Ziervogel, K.; Cox, C.; Steen, A.D.; Teske, A. Verrucomicrobia Are Candidates for Polysaccharide-Degrading Bacterioplankton in an Arctic Fjord of Svalbard. Appl. Environ. Microbiol. 2014, 80, 3749–3756. [Google Scholar] [CrossRef]
- Lusty, M.W.; Gobler, C.J. Repeated Hydrogen Peroxide Dosing Briefly Reduces Cyanobacterial Blooms and Microcystin While Increasing Fecal Bacteria Indicators in a Eutrophic Pond. J. Environ. Sci. 2023, 124, 522–543. [Google Scholar] [CrossRef]
- Chen, C.; Wang, Y.; Chen, K.; Shi, X.; Yang, G. Using Hydrogen Peroxide to Control Cyanobacterial Blooms: A Mesocosm Study Focused on the Effects of Algal Density in Lake Chaohu, China. Environ. Pollut. 2021, 272, 115923. [Google Scholar] [CrossRef] [PubMed]
- Yéprémian, C.; Catherine, A.; Bernard, C.; Congestri, R.; Elersek, T.; Pilkaityte, R. Chlorophyll a Extraction and Determination. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis, 1st ed.; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; Wiley: Chichester, UK, 2016; pp. 331–334. [Google Scholar]
- Yéprémian, C.; Catherine, A.; Bernard, C.; Congestri, R.; Elersek, T.; Pilkaityte, R. Phycocyanin extraction and determination. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis, 1st ed.; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; Wiley: Chichester, UK, 2016; pp. 335–338. [Google Scholar]
- Lund, J.W.G.; Kipling, C.; Le Cren, E.D. The inverted microscope method of estimating algal numbers and the statistical basis of estimations by counting. Hydrobiologia 1958, 11, 143–170. [Google Scholar] [CrossRef]
- Zhou, Q.; Chen, W.; Zhang, H.; Peng, L.; Liu, L.; Han, Z.; Wan, N.; Li, L.; Song, L. A flow cytometer based protocol for quantitative analysis of bloom-forming cyanobacteria (Microcystis) in lake sediments. J. Environ. Sci. 2012, 24, 1709–1716. [Google Scholar] [CrossRef] [PubMed]
- Utermöhl, H. Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar] [CrossRef]
- Pappas, J.L.; Stoermer, E.F. Quantitative method for determining a representative algal sample count1. J. Phycol. 1996, 32, 693–696. [Google Scholar] [CrossRef]
- Djurhuus, A.; Port, J.; Closek, C.J.; Yamahara, K.M.; Romero-Maraccini, O.; Walz, K.R.; Goldsmith, D.B.; Michisaki, R.; Breitbart, M.; Boehm, A.B.; et al. Evaluation of Filtration and DNA Extraction Methods for Environmental DNA Biodiversity Assessments across Multiple Trophic Levels. Front. Mar. Sci. 2017, 4, 314. [Google Scholar] [CrossRef]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every Base Matters: Assessing Small Subunit rRNA Primers for Marine Microbiomes with Mock Communities, Time Series and Global Field Samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Bower, S.M.; Carnegie, R.B.; Goh, B.; Jones, S.R.M.; Lowe, G.J.; Mak, M.W.S. Preferential PCR Amplification of Parasitic Protistan Small Subunit RDNA from Metazoan Tissues. J. Eukaryot. Microbiol. 2004, 51, 325–332. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 22 May 2022).
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Lefler, F.W.; Berthold, D.E.; Laughinghouse, H.D. CyanoSeq: A database of cyanobacterial 16S rRNA gene sequences with curated taxonomy. J. Phycol. 2023, 59, 470–480. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Guillou, L.; Bachar, D.; Audic, S.; Bass, D.; Berney, C.; Bittner, L.; Boutte, C.; Burgaud, G.; De Vargas, C.; Decelle, J.; et al. The Protist Ribosomal Reference Database (PR2): A Catalog of Unicellular Eukaryote Small Sub-Unit RRNA Sequences with Curated Taxonomy. Nucleic Acids Res. 2012, 41, D597–D604. [Google Scholar] [CrossRef]
- Schloss, P.D. Waste Not, Want Not: Revisiting the Analysis That Called into Question the Practice of Rarefaction. mSphere 2024, 9, e00355-23. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. R Package, version 2.5-6; Vegan: Community Ecology Package.
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lefler, F.W.; Barbosa, M.; Berthold, D.E.; Roten, R.; Bishop, W.M.; Laughinghouse, H.D., IV. Microbial Community Response to Granular Peroxide-Based Algaecide Treatment of a Cyanobacterial Harmful Algal Bloom in Lake Okeechobee, Florida (USA). Toxins 2024, 16, 206. https://doi.org/10.3390/toxins16050206
Lefler FW, Barbosa M, Berthold DE, Roten R, Bishop WM, Laughinghouse HD IV. Microbial Community Response to Granular Peroxide-Based Algaecide Treatment of a Cyanobacterial Harmful Algal Bloom in Lake Okeechobee, Florida (USA). Toxins. 2024; 16(5):206. https://doi.org/10.3390/toxins16050206
Chicago/Turabian StyleLefler, Forrest W., Maximiliano Barbosa, David E. Berthold, Rory Roten, West M. Bishop, and H. Dail Laughinghouse, IV. 2024. "Microbial Community Response to Granular Peroxide-Based Algaecide Treatment of a Cyanobacterial Harmful Algal Bloom in Lake Okeechobee, Florida (USA)" Toxins 16, no. 5: 206. https://doi.org/10.3390/toxins16050206
APA StyleLefler, F. W., Barbosa, M., Berthold, D. E., Roten, R., Bishop, W. M., & Laughinghouse, H. D., IV. (2024). Microbial Community Response to Granular Peroxide-Based Algaecide Treatment of a Cyanobacterial Harmful Algal Bloom in Lake Okeechobee, Florida (USA). Toxins, 16(5), 206. https://doi.org/10.3390/toxins16050206