Effectiveness of the High Dose/Refuge Strategy for Managing Pest Resistance to Bacillus thuringiensis (Bt) Plants Expressing One or Two Toxins

Abstract
:1. Introduction
2. Materials and Methods
2.1. Parameters

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Default-Value | Tested-Value | ||
|---|---|---|---|---|
| Operational Parameters | Refuge zone proportion | v | 0.05 [37] | 0.05–0.10–0.20–0.30–0.40 |
| Selection of the toxin A | sBtA | 1 [37] | 1–0.93–0.50 | |
| Selection of the toxin B | sBtB | 1 [37] | 1–0.93–0.50 | |
| Field area (hectare) | 260 [38] | na | ||
| Plants/hectare | 67,000 [38] | na | ||
| Biological Parameters | Initial Ar frequency | ArFreq | 1.5 × 10−3 [39] | 1.5 × 10−2–1.5 × 10−1 |
| Initial Br frequency | BrFreq | 1.5 × 10−3 [39] | 1.5 × 10−2–1.5 × 10−1 | |
| Ar dominance | hAr | 0 [12,40] | 0–0.23–0.53 | |
| Br dominance | hBr | 0 [12,40] | 0–0.23–0.53 | |
| Fitness cost associated to Ar | fcostA | 0.15 [41] | na | |
| Fitness cost associated to Br | fcostB | 0.15 [41] | na | |
| Fitness cost dominance associated to Ar | hfcA | 0 [42,43,44] | na | |
| Fitness cost dominance associated to Br | hfcB | 0 [42,43,44] | na | |
| Initial individual number/ha | nzero | 50,000 [38] | na | |
| Intrinsic growth rate | r | 0.15 [45,46] | na | |
| Carrying capacity/plant | K | 22 [38] | na |
2.1.1. Resistance Alleles
2.1.2. Relative Size of the Refuge Zone (v)
2.1.3. Mortality Exerted by the Bt Plants on Susceptible Insects and Resistance Allele Dominance
2.1.4. Fitness Cost of Resistance and Fitness Cost Dominance
2.1.5. Fitness Estimation

2.1.6. Initial Individual Number (Nzero); Carrying Capacity (K); Population Intrinsic Growth Rate (r):
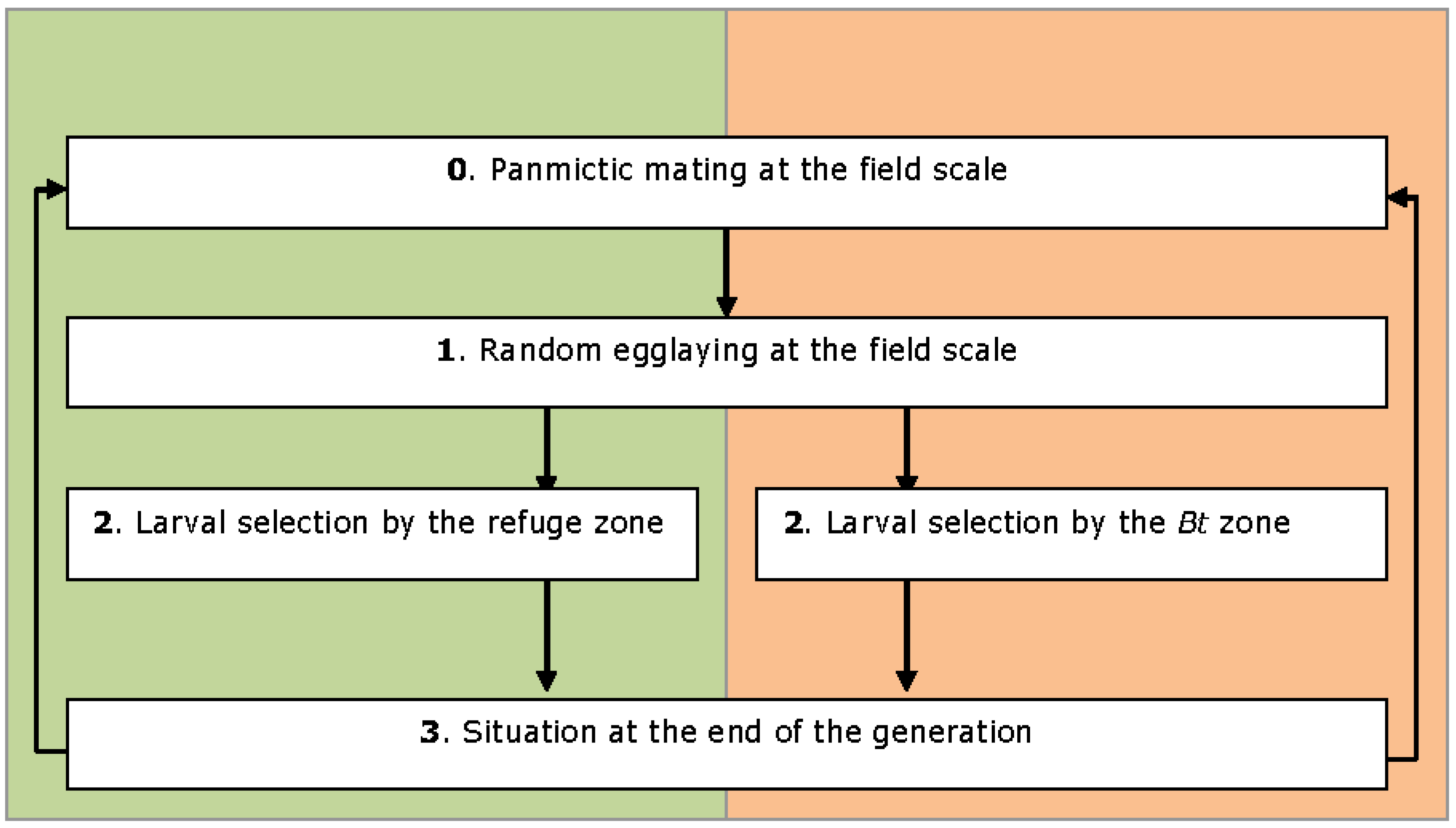
2.2. Model Description


2.3. Model Output
3. Results and Discussion
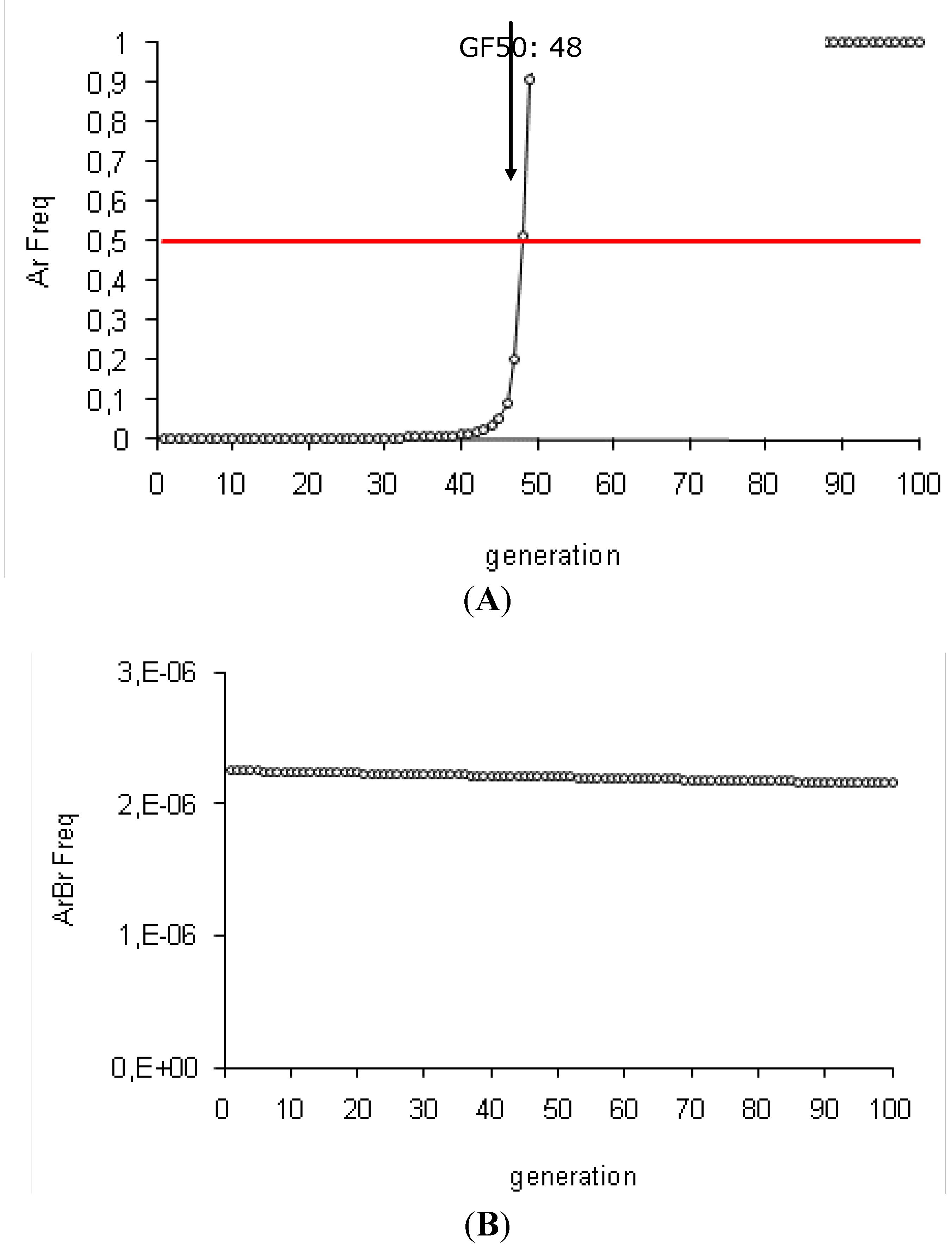
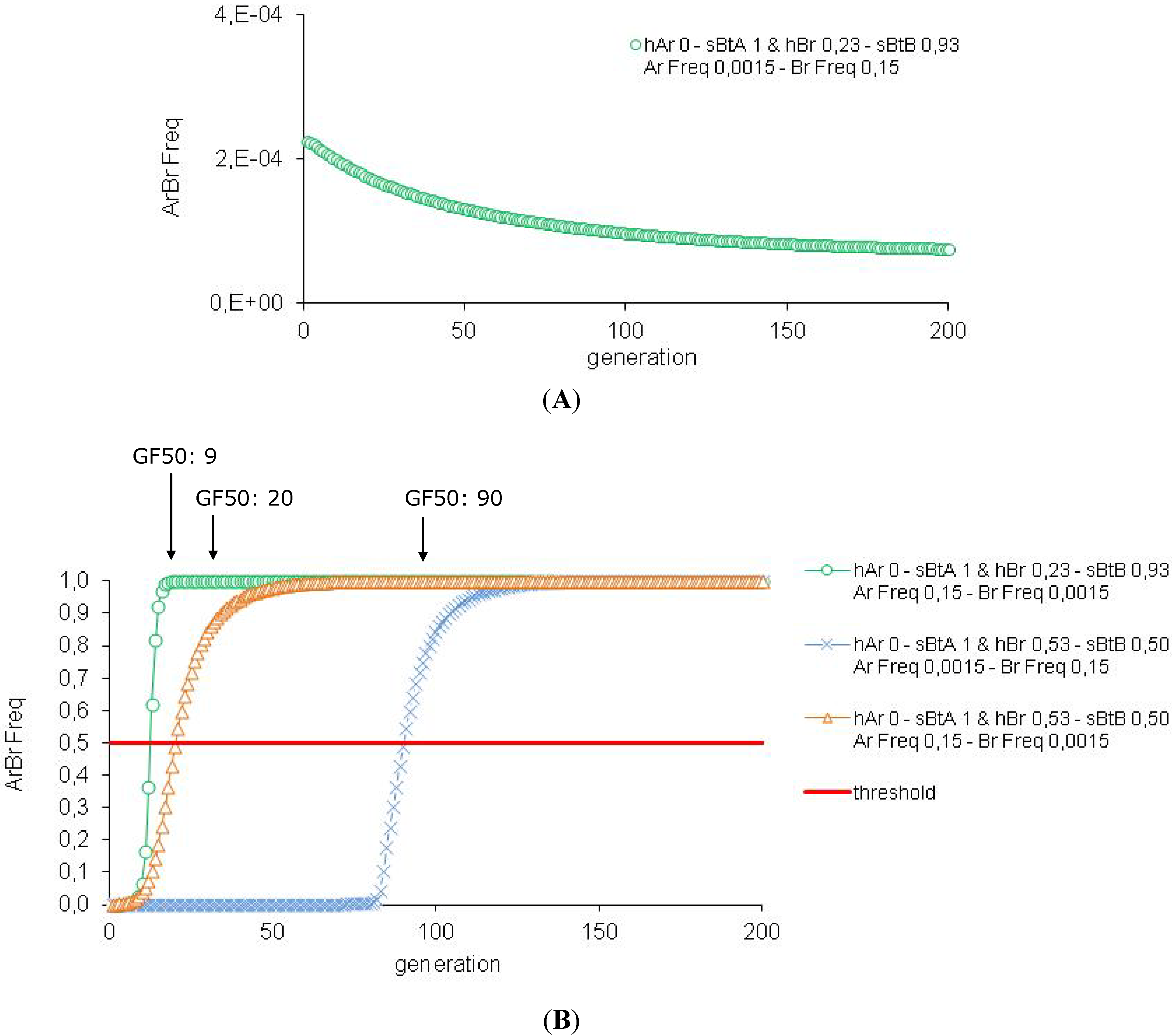
3.1. Efficiency of the HD/R Strategy for Bt Plants Expressing High Dose of One or Two Toxins, with Initially Rare Resistance Alleles

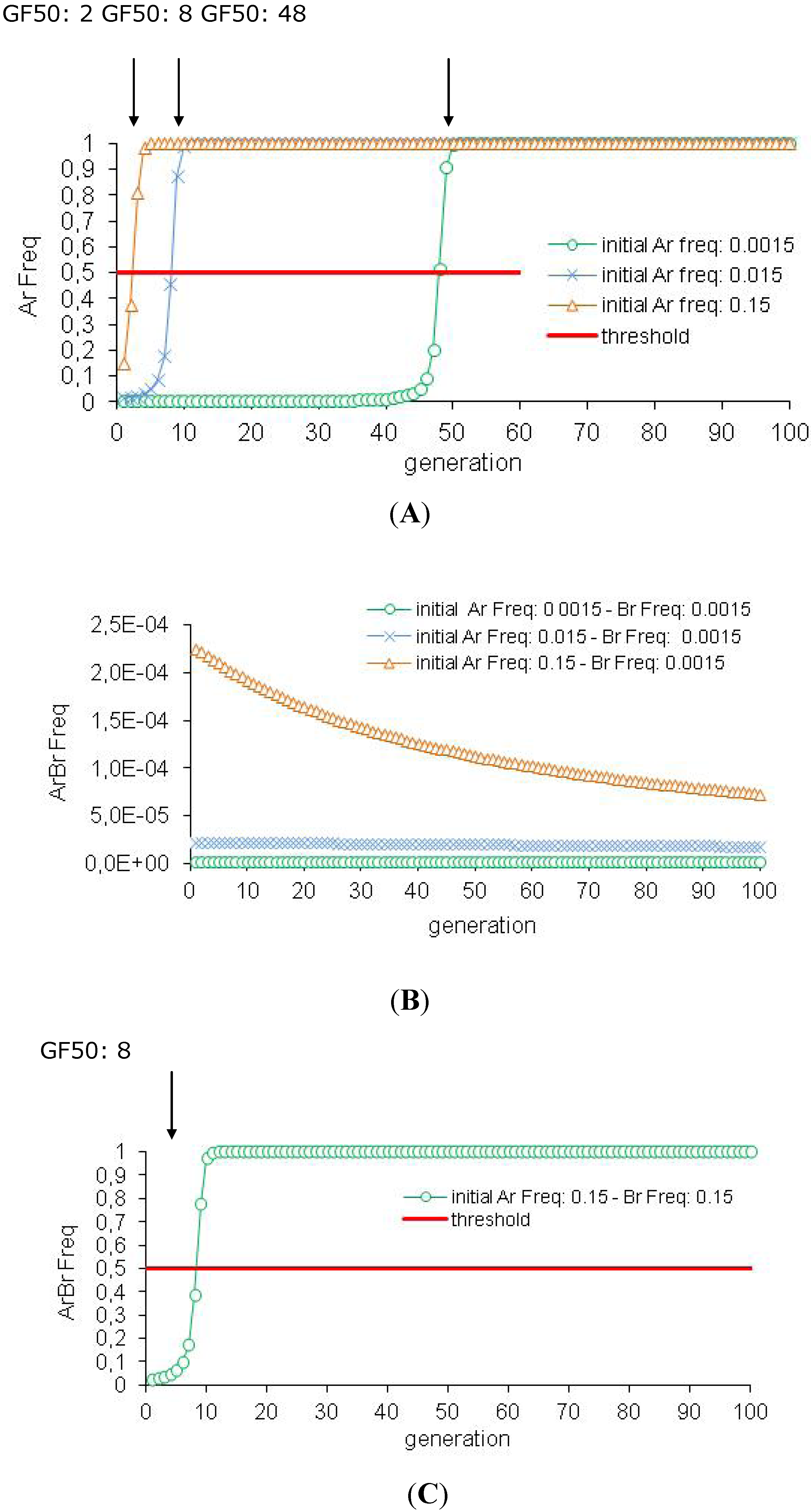
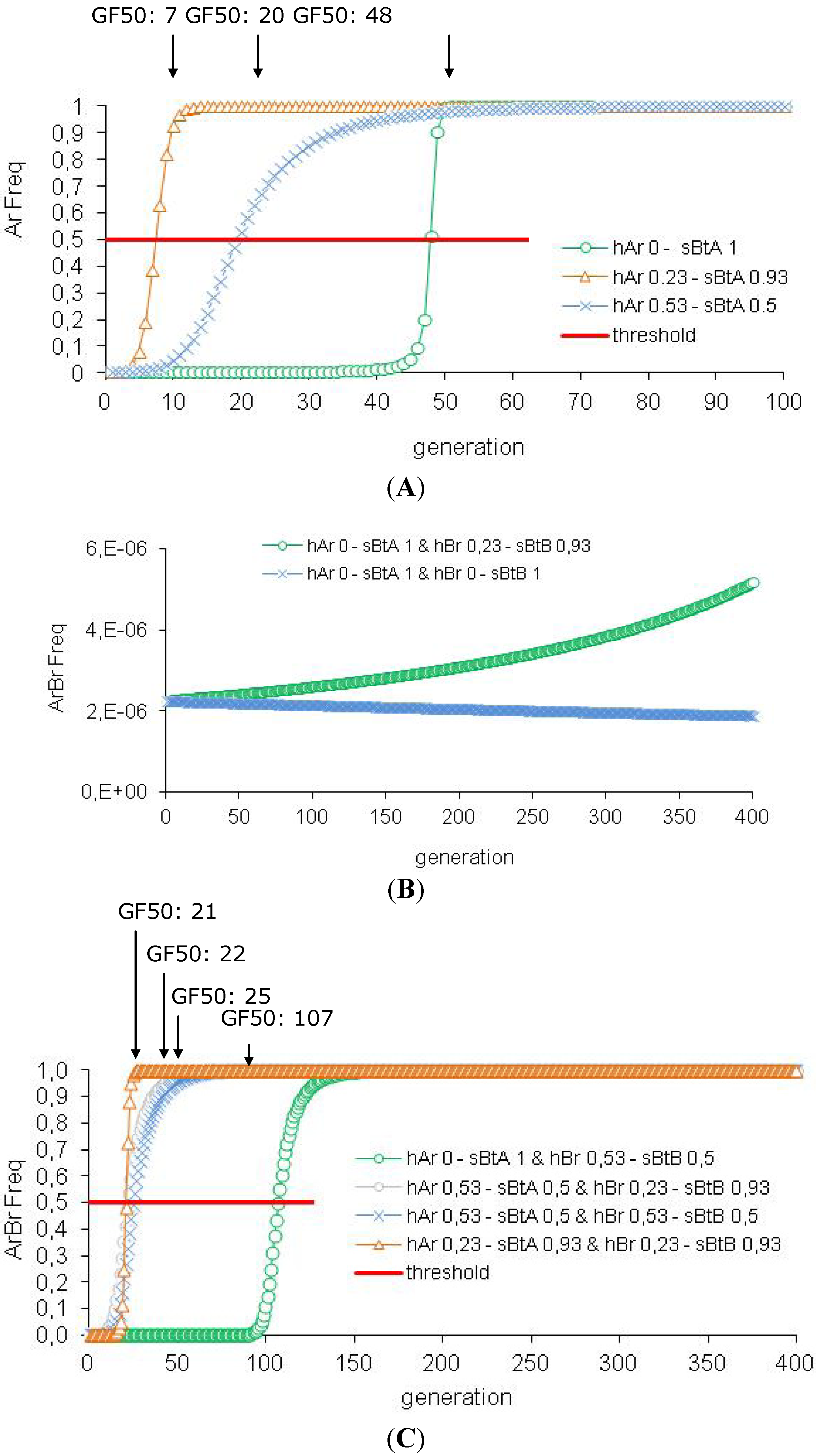
3.2. Efficiency of the HD/R Strategy if Resistance Is Initially Not Rare in the Population
| A. | Bt plants Synthesizing One Toxin | GF50 | % Pop. Decrease | |||
| Gen 2/Gen1 | Gen 10/Gen 1 | |||||
| 1 | Initial Conditions (sBtA = 1, hAr = 0, Ar Freq = 0.001, v = 5%) | 48 | 94 | 1 | ||
| 2 | Impact of the Ar Frequency | Ar Freq 0.0015 | 48 | 94 | 1 | |
| Ar Freq 0.015 | 8 | 94 | 99.9 | |||
| Ar Freq 0.15 | 2 | 92 | 99.1 | |||
| 3 | Impact of the Cry Concentration | hAr 0-sBtA 1 | 48 | 94 | 1 | |
| hAr 0.23-sBtA 0.93 | 7 | 86 | 99.9 | |||
| hAr 0.53-sBtA 0.50 | 20 | 39 | 98.7 | |||
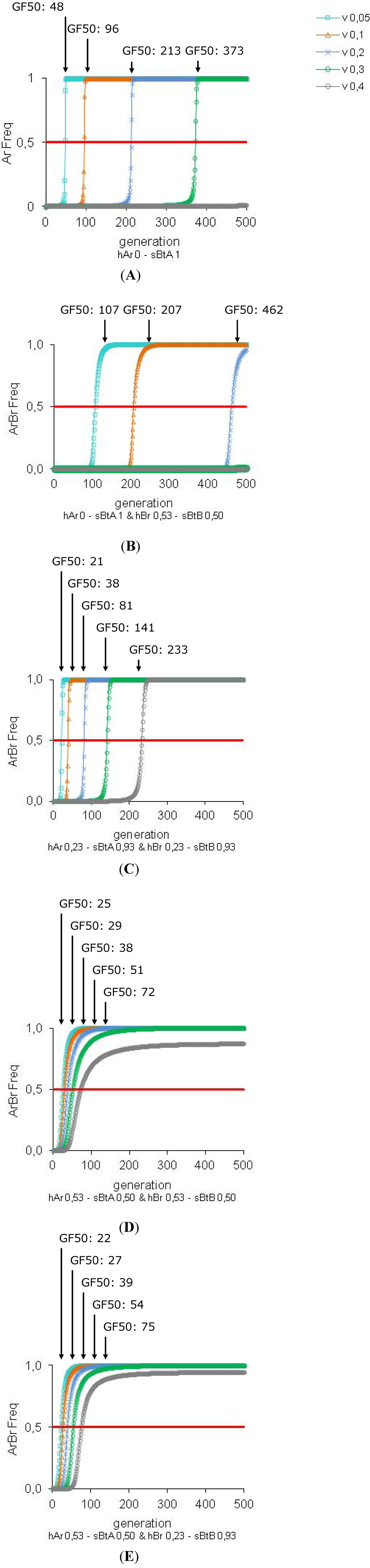
| 4 | Impact of the Refuge Zone Proportion | v 0.05 | 48 | 94 | 1 | |
| v 0.1 | 96 | 88 | 99.9 | |||
| v 0.2 | 213 | 77 | 99.9 | |||
| v 0.3 | 373 | 65 | 98.9 | |||
| v 0.4 | <500 | 54 | 99.8 | |||
| B. | Bt plants synthesizing two toxins | GF50 | % pop. decrease | |||
| Gen 2/Gen1 | Gen 10/Gen 1 | |||||
| 1 | Initial Conditions | >500 | 94 | 1 | ||
| 2 | Impact of the Ar Frequency(Br: 0.0015) | Ar Freq 0.0015 | >500 | 94 | 1 | |
| Ar Freq 0.015 | >500 | 94 | 1 | |||
| Ar Freq 0.15 | >500 | 94 | 1 | |||
| 3 | Impact of the Ar and Br Frequencies | Ar Freq 0.15-Br Freq 0.15 | 8 | 94 | 99.9 | |
| 4 | Impact of the Cryconcentration | hAr 0-sBtA 1 | hBr 0-sBtB 1 | >500 | 94 | 1 |
| hBr 0.23-sBtB 0.93 | >500 | 94 | 1 | |||
| hBr 0.53-sBtB 0.50 | 107 | 94 | 1 | |||
| hAr 0.53-sBtA 0.50 | hBr 0.23-sBtB 0.93 | 22 | 90 | 99.9 | ||
| hBr 0.53-sBtB 0.50 | 25 | 67 | 99.9 | |||
| hAr 0.23-sBtA 0.93 | hBr 0.23-sBtB 0.93 | 21 | 94 | 1 | ||
| 5 | Impact of the Cry Concentration and of the Resistance Alleles Frequency | hAr 0-sBtA 1 hBr 0.23-sBtB 0.93 | Ar Freq 0.0015-Br Freq 0.15 | >500 | 94 | 1 |
| Ar Freq0.15-Br Freq 0.0015 | 9 | 94 | q | |||
| hAr 0-sBtA 1 hBr 0.53-sBtB 0.50 | Ar Freq 0.0015-Br Freq 0.15 | 90 | 94 | 1 | ||
| Ar Freq0.15-Br Freq 0.0015 | 20 | 93 | 99.9 | |||
| 6 | Impact of the Toxin Concentrations and of the Refuge Zone Proportion | hAr 0-sBtA 1 hBr 0.53-sBtB 0.50 | v 0.05 | 107 | 94 | 1 |
| v 0.1 | 207 | 88 | 99.9 | |||
| v 0.2 | 462 | 77 | 99.9 | |||
| v 0.3 | >500 | 65 | 99.9 | |||
| v 0.4 | >500 | 54 | 99.8 | |||
| hAr 0.53-sBtA 0.50 hBr 0.53-sBtB 0.50 | v 0.05 | 25 | 67 | 99.9 | ||
| v 0.1 | 29 | 62 | 99.9 | |||
| v 0.2 | 38 | 53 | 99.9 | |||
| v 0.3 | 51 | 45 | 99.5 | |||
| v 0.4 | 72 | 36 | 98.2 | |||
| hAr 0.53-sBtA 0.50 hBr 0.23-sBtB 0.93 | v 0.05 | 22 | 90 | 99.9 | ||
| v 0.1 | 27 | 85 | 99.9 | |||
| v 0.2 | 39 | 73 | 99.9 | |||
| v 0.3 | 54 | 62 | 99.9 | |||
| v 0.4 | 75 | 51 | 99.8 | |||
| hAr 0.23-sBtA 0.93 hBr 0.23-sBtB 0.93 | v 0.05 | 21 | 94 | 1 | ||
| v 0.1 | 38 | 87 | 99.9 | |||
| v 0.2 | 81 | 76 | 99.9 | |||
| v 0.3 | 141 | 65 | 99.9 | |||
| v 0.4 | 233 | 53 | 99.8 | |||

3.3. Efficiency of the HD/R Strategy If Bt Plants Produce a Low Toxin Concentration (for One or Two Toxins)

3.4. Efficiency of the HD/R Strategy If Resistance Is not Rare in the Population and Two-Toxin Bt Plants Produce a Low Toxin Concentration

3.5. Efficiency of the HD/R Strategy in Relation to the Refuge Zone Proportion

4. Conclusions
Acknowledgments
Conflict of Interests
References
- Crickmore, N.; Zeigler, D.R.; Feitelson, J.; Schnepf, E.; Van Rie, J.; Lereclus, D.; Baum, J.; Dean, D.H. Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 807. [Google Scholar]
- Gill, S.S.; Cowles, E.A.; Pietrantonio, P.V. The Mode of Action of Bacillus thuringiensis Endotoxins. Annu. Rev. Entomol. 1992, 37, 615–634. [Google Scholar] [CrossRef]
- Hofte, H.; Whiteley, H.R. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Mol. Biol. Rev. 1989, 53, 242. [Google Scholar]
- Bauer, L.S. Resistance: A threat to the insecticidal crystal proteins of Bacillus thuringiensis. Florida Entomol. 1995, 78, 414–443. [Google Scholar] [CrossRef]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberón, M. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochem. Mol. Biol. 2011, 41, 423–431. [Google Scholar] [CrossRef]
- Barton, K.A.; Whiteley, H.R.; Yang, N.-S. Bacillus thuringiensis §-Endotoxin Expressed in Transgenic Nicotiana tabacum Provides Resistance to Lepidopteran Insects. Plant Physiol. 1987, 85, 1103–1109. [Google Scholar] [CrossRef]
- Vaeck, M.; Reynaerts, A.; Hofte, H.; Jansens, S.; De Beuckeleer, M.; Dean, C.; Zabeau, M.; Montagu, M.V.; Leemans, J. Transgenic plants protected from insect attack. Nature 1987, 328, 33–37. [Google Scholar] [CrossRef]
- James, C. Global Status of Commercialized Biotech/GM Crops: 2010; ISAAA Brief No. 42; ISAAA: Ithaca, NY, USA, 2010. [Google Scholar]
- Gould, F. Sustainability of transgenic insecticidal cultivars: Integrating pest genetics and ecology. Ann. Rev. Entomol. 1998, 43, 701–726. [Google Scholar] [CrossRef]
- Caprio, M.; Summerford, D.; Simms, S. Evaluating transgenic plants for suitability in pest and resistance management programs. In Field Manual of Techniques in Invertebrate Pathology: Application and Evaluation of Pathogens for Control of Insects and Other Invertebrate Pests; Springer: Amsterdam, The Netherlands, 2000; pp. 805–828. [Google Scholar]
- Gould, F.; Anderson, A.; Reynolds, A.; Bumgarner, L.; Moar, W. Selection and genetic analysis of a Heliothis virescens (Lepidoptera: Noctuidae) strain with high levels of resistance to Bacillus thuringiensis toxins. J. Econ. Entomol. 1995, 88, 1545–1559. [Google Scholar]
- Tabashnik, B.E.; Patin, A.L.; Dennehy, T.J.; Liu, Y.B.; Carrière, Y.; Sims, M.A.; Antilla, L. Frequency of resistance to Bacillus thuringiensis in field populations of pink bollworm. Proc. Nat. Acad. Sci. USA 2000, 97, 12980. [Google Scholar]
- Liu, Y.-B.; Tabashnik, B.E.; Dennehy, T.J.; Patin, A.L.; Sims, M.A.; Meyer, S.K.; Carrière, Y. Effects of Bt Cotton and Cry1Ac Toxin on Survival and Development of Pink Bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2001, 94, 1237–1242. [Google Scholar] [CrossRef]
- Comins, H.N. The development of insecticide resistance in the presence of migration. J. Theor. Biol. 1977, 64, 177–197. [Google Scholar] [CrossRef]
- Comins, H.N. The management of pesticide resistance. J. Theor. Biol. 1977, 65, 399–420. [Google Scholar] [CrossRef]
- Tabashnik, B.E. Computer simulation as a tool for pesticide resistance management. In Pesticide Resistance: Strategies and Tactics for Management; The National Academies Press: Washington, DC, USA, 1986. [Google Scholar]
- Tabashnik, B.E. Delaying insect adaptation to transgenic plants: Seed mixtures and refugia reconsidered. Proc. Biol. Sci. 1994, 7–12. [Google Scholar] [CrossRef]
- Alstad, D.N.; Andow, D.A. Managing the Evolution of Insect Resistance to Transgenic Plants. Science 1995, 268, 1894–1896. [Google Scholar]
- Storer, N.P. A Spatially Explicit Model Simulating Western Corn Rootworm (Coleoptera: Chrysomelidae) Adaptation to Insect-Resistant Maize. J. Econ. Entomol. 2003, 96, 1530–1547. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Van Rensburg, J.B.J.; Carrière, Y. Field-Evolved Insect Resistance to Bt Crops: Definition, Theory, and Data. J. Econ. Entomol. 2009, 102, 2011–2025. [Google Scholar] [CrossRef]
- US EPA. Current & Previously Registered Section 3 PIP Registrations. Available online: http://www.epa.gov/oppbppd1/biopesticides/pips/pip_list.htm (accessed on 18 April 2012).
- Stewart, S.D.; Adamczyk, J.J., Jr.; Knighten, K.S.; Davis, F.M. Impact of Bt cottons expressing one or two insecticidal proteins of Bacillus thuringiensis Berliner on growth and survival of 8802616noctuid (Lepidoptera) larvae. J. Econ. Entomol. 2001, 94, 752–760. [Google Scholar] [CrossRef]
- Mani, G.S. Evolution of resistance in the presence of two insecticides. Genetics 1985, 109, 761–783. [Google Scholar]
- Tabashnik, B.E. Managing resistance with multiple pesticide tactics: Theory, evidence, and recommendations. J. Econ. Entomol. 1989, 82, 1263–1269. [Google Scholar]
- Roush, R.T. Managing pests and their resistance to Bacillus thuringiensis: Can transgenic crops be better than sprays? Biocontrol. Sci. Technol. 1994, 4, 501–516. [Google Scholar] [CrossRef]
- Livingston, M.J.; Carlson, G.A.; Fackler, P.L. Managing Resistance Evolution in Two Pests to Two Toxins with Refugia. Am. J. Agric. Econ. 2004, 86, 1–13. [Google Scholar] [CrossRef]
- Gould, F.; Cohen, M.B.; Bentur, J.S.; Kennedy, G.G.; Van Duyn, J. Impact of small fitness costs on pest adaptation to crop varieties with multiple toxins: a heuristic model. J. Econ. Entomol. 2006, 99, 2091–2099. [Google Scholar] [CrossRef]
- Georghiou, G.P.; Wirth, M.C. Influence of exposure to single versus multiple toxins of Bacillus thuringiensis subsp. israelensis on development of resistance in the mosquito Culex quinquefasciatus (Diptera: Culicidae). Appl. Environ. Microbiol. 1997, 63, 1095. [Google Scholar]
- Caprio, M.A. Evaluating Resistance Management Strategies for Multiple Toxins in the Presence of External Refuges. J. Econ. Entomol. 1998, 91, 1021–1031. [Google Scholar]
- Zhao, J.-Z.; Cao, J.; Collins, H.L.; Bates, S.L.; Roush, R.T.; Earle, E.D.; Shelton, A.M. Concurrent use of transgenic plants expressing a single and two Bacillus thuringiensis genes speeds insect adaptation to pyramided plants. Proc. Nat. Acad. Sci. USA 2005, 102, 8426–8430. [Google Scholar]
- Roush, R.T. Two-toxin strategies for management of insecticidal transgenic crops: Can pyramiding succeed where pesticide mixtures have not? Philos. Trans. R. Soc. Lond. B Biol. Sci. 1998, 353, 1777–1786. [Google Scholar] [CrossRef]
- Curtis, C.F. Theoretical models of the use of insecticide mixtures for the management of resistance. Bull. Entomol. Res. 1985, 75, 259–265. [Google Scholar] [CrossRef]
- Heckel, D.G.; Gahan, L.J.; Baxter, S.W.; Zhao, J.Z.; Shelton, A.M.; Gould, F.; Tabashnik, B.E. The diversity of Bt resistance genes in species of Lepidoptera. J. Invertebr. Pathol. 2007, 95, 192–197. [Google Scholar] [CrossRef]
- Monsanto website. Website online: http://monsanto.mediaroom.com/ (accessed on 15 October 2012).
- Downes, S.; Parker, T.; Mahon, R. Incipient Resistance of Helicoverpa punctigera to the Cry2Ab Bt Toxin in Bollgard II® Cotton. PLoS One 2010, 5, e12567. [Google Scholar]
- Georghiou, G.P.; Taylor, C.E. Genetic and biological influences in the evolution of insecticide resistance. J. Econ. Entomol. 1977, 70, 319–323. [Google Scholar]
- Introduction to Biotechnology Regulation for Pesticides< |Pesticides| US EPA. Website online: http://www.epa.gov/oppbppd1/biopesticides/regtools/biotech-reg-prod.htm#crops (accessed on 27 April 2012).
- Guse, C.A.; Onstad, D.W.; Buschman, L.L.; Porter, P.; Higgins, R.A.; Sloderbeck, P.E.; Cronholm, G.B.; Peairs, F.B. Modeling the development of resistance by stalk-boring Lepidoptera (Crambidae) in areas with irrigated transgenic corn. Environ. Entomol. 2002, 31, 676–685. [Google Scholar] [CrossRef]
- Andow, D.A.; Olson, D.M.; Hellmich, R.L.; Alstad, D.N.; Hutchison, W.D. Frequency of resistance to Bacillus thuringiensis toxin Cry1Ab in an Iowa population of European corn borer (Lepidoptera: Crambidae). J. Econ. Entomol. 2000, 93, 26–30. [Google Scholar] [CrossRef]
- Liu, Y.B.; Tabashnik, B.E.; Meyer, S.K.; Carrière, Y.; Bartlett, A.C. Genetics of pink bollworm resistance to Bacillus thuringiensis toxin Cry1Ac. J. Econ. Entomol. 2001, 94, 248–252. [Google Scholar] [CrossRef]
- Vacher, C.; Bourguet, D.; Rousset, F.; Chevillon, C.; Hochberg, M.E. Modelling the spatial configuration of refuges for a sustainable control of pests: A case study of Bt cotton. J. Evol. Biol. 2003, 16, 378–387. [Google Scholar] [CrossRef]
- Tabashnik, B.; Gould, F.; Carriere, Y. Delaying evolution of insect resistance to transgenic crops by decreasing dominance and heritability. J. Evolution. Biol. 2004, 17, 904–912. [Google Scholar] [CrossRef]
- Carriére, Y.; Ellers-Kirk, C.; Liu, Y.B.; Sims, M.A.; Patin, A.L.; Dennehy, T.J.; Tabashnik, B.E. Fitness costs and maternal effects associated with resistance to transgenic cotton in the pink bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2001, 94, 1571–1576. [Google Scholar] [CrossRef]
- Carrière, Y.; Ellers-kirk, C.; Patin, A.L.; Sims, M.A.; Meyer, S.; Liu, Y.; Dennehy, T.J.; Tabashnik, B.E. Overwintering cost associated with resistance to transgenic cotton in the pink bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2001, 94, 935–941. [Google Scholar] [CrossRef]
- Sayyed, A.H.; Wright, D.J. Fitness costs and stability of resistance to Bacillus thuringiensis in a field population of the diamondback moth Plutella xylostella L. Ecol. Entomol. 2001, 26, 502–508. [Google Scholar] [CrossRef]
- Bird, L.J.; Akhurst, R.J. Relative fitness of Cry1A-resistant and -susceptible Helicoverpa armigera (Lepidoptera: Noctuidae) on conventional and transgenic cotton. J. Econ. Entomol. 2004, 97, 1699–1709. [Google Scholar] [CrossRef]
- Conner, J.K.; Hartl, D.L. A Primer of Ecological Genetics; Sinauer Associates: Massachussets, MA, USA, 2004. [Google Scholar]
- Gould, F.; Martinez-Ramirez, A.; Anderson, A.; Ferre, J.; Silva, F.J.; Moar, W.J. Broad-spectrum resistance to Bacillus thuringiensis toxins in Heliothis virescens. Proc. Nat. Acad. Sci. 1992, 89, 79–86. [Google Scholar]
- McGaughey, W.H.; Johnson, D.E. Influence of Crystal Protein-Composition of Bacillus-Thuringiensis Strains on Cross-Resistance in Indianmeal Moths (Lepidoptera, Pyralidae). J. Econ. Entomol. 1994, 87, 535–540. [Google Scholar]
- US EPA. Bt Plant-Pesticides Biopesticides Registration Action Document. Website online: http://www.epa.gov/scipoly/sap/meetings/2000/october/brad4_irm.pdf (accessed on 15 October 2012).
- Onstad, D.W.; Meinke, L.J. Modeling evolution of Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae) to transgenic corn with two insecticidal traits. J. Econ. Entomol. 2010, 103, 849–860. [Google Scholar] [CrossRef]
- Shelton, A.M.; Zhao, J.Z.; Roush, R.T. Economic, ecological, food safety, and social consequences of the deployment of Bt transgenic plants. Ann. Rev. Entomol. 2002, 47, 845–881. [Google Scholar] [CrossRef]
- Monsanto proposes natural refuge for Bollgard II cotton, Management content from Delta Farm Press. Website online: http://deltafarmpress.com/management/monsanto-proposes-natural-refuge-bollgard-ii-cotton (accessed on 1 June 2012).
- US EPA. EPA Approves Natural Refuge for Insect Resistance Management in Bollgard II Cotton. Website online: http://www.epa.gov/oppfead1/cb/csb_page/updates/2007/bollgard-cotton.htm (accessed on 1 June 2012).
- Tabashnik, B.E.; Liu, Y.; Malvar, T.; Heckel, D.G.; Masson, L.; Ferre, J. Insect resistance to Bacillus thuringiensis: Uniform or Diverse? Phil. Tran. R. Soc. Lond. Series B Biol. Sci. 1998, 353, 1751–1756. [Google Scholar] [CrossRef]
- Frutos, R.; Rang, C.; Royer, M. Managing insect resistance to plants producing Bacillus thuringiensis toxins. Crit. Rev. Biotechnol. 1999, 19, 227–276. [Google Scholar] [CrossRef]
- Rawlings, P.; Davidson, G.; Sakai, R.K.; Rathor, H.R.; Aslamkhan, M.; Curtis, C.F. Field measurement of the effective dominance of an insecticide resistance in anopheline mosquitos. Bull. World Health Organ. 1981, 59, 631–640. [Google Scholar]
- Sayyed, A.H.; Haward, R.; Herrero, S.; Ferre, J.; Wright, D.J. Genetic and Biochemical Approach for Characterization of Resistance to Bacillus thuringiensis Toxin Cry1Ac in a Field Population of the Diamondback Moth, Plutella xylostella. Appl. Environ. Microbiol. 2000, 66, 1509–1516. [Google Scholar] [CrossRef]
- Bourguet, D.; Prout, M.; Raymond, M. Dominance of insecticide resistance presents a plastic response. Genetics 1996, 143, 407. [Google Scholar]
- Raymond, B.; Sayyed, A.H.; Wright, D.J. Genes and Environment Interact to Determine the Fitness Costs of Resistance to Bacillus thuringiensis. Proc. R. Soc. B Biol. Sci. 2005, 272, 1519–1524. [Google Scholar] [CrossRef]
- Carriére, Y.; Tabashnik, B. Reversing insect adaptation to transgenic insecticidal plants. Proc. R. Soc. Lond. Series B Biol. Sci. 2001, 268, 1475. [Google Scholar] [CrossRef]
- Vacher, C.; Bourguet, D.; Rousset, F.; Chevillon, C.; Hochberg, M.E. High dose refuge strategies and genetically modified crops–reply to Tabashnik et al. J. Evol. Biol. 2004, 17, 913–918. [Google Scholar] [CrossRef]
- Lenormand, T.; Raymond, M. Resistance management: The stable zone strategy. Proc. Biol. Sci. 1998, 265, 1985–1990. [Google Scholar] [CrossRef]
- Orr, D.B.; Landis, D.A. Oviposition of European corn borer (Lepidoptera: Pyralidae) and impact of natural enemy populations in transgenic versus isogenic corn. J. Econ. Entomol. 1997, 90, 905–909. [Google Scholar]
- Ramachandran, S.; Buntin, G.D.; All, J.N.; Tabashnik, B.E.; Raymer, P.L.; Adang, M.J.; Pulliam, D.A.; Stewart, C.N., Jr. Survival, development, and oviposition of resistant diamondback moth (Lepidoptera: Plutellidae) on transgenic canola producing a Bacillus thuringiensis toxin. J. Econ. Entomol. 1998, 91, 1239–1244. [Google Scholar]
- Anonymous The R Project for Statistical Computing. Website online: http://www.r-project.org/ (accessed on 23 May 2012).
- Tabashnik, B.E.; Croft, B.A. Managing pesticide resistance in crop-arthropod complexes: Interactions between biological and operational factors. Environ. Entomol. 1982, 11, 1137–1144. [Google Scholar]
- Mallet, J.; Porter, P. Preventing insect adaptation to insect-resistant crops: Are seed mixtures or refugia the best strategy? Proc. Biol. Sci. 1992, 165–169. [Google Scholar] [CrossRef]
- Crow, J.F.; Kimura, M. An Introduction to Population Genetic Theory; Harper and Row: New York, NY, USA, 1970. [Google Scholar]
- Ricker, W.E. Stock and recruitment. J. Fish. Board Can. 1954, 11, 559–623. [Google Scholar] [CrossRef]
- Gould, F. Bt-resistance management-theory meets data. Nat. Biotechnol. 2003, 21, 1450–1451. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Gassmann, A.J.; Crowder, D.W.; Carrière, Y. Insect resistance to Bt crops: Evidence versus theory. Nat. Biotechnol. 2008, 26, 199–202. [Google Scholar] [CrossRef]
- Luttrell, R.G.; Ali, I.; Allen, K.C.; Young, S.Y., III; Szalanski, A.; Williams, K.; Lorenz, G.; Parker, C.D., Jr.; Blanco, C. Resistance to Bt in Arkansas populations of cotton bollworm. In Proceedings of 2004 Beltwide Cotton Conferences, San Antonio, TE, USA; 2004. [Google Scholar]
- Ali, M.I.; Luttrell, R.G.; Young, S.Y., III. Susceptibilities of Helicoverpa zea and Heliothis virescens (Lepidoptera: Noctuidae) populations to Cry1Ac insecticidal protein. J. Econ. Entomol. 2006, 99, 164–175. [Google Scholar] [CrossRef]
- Matten, S.R.; Head, G.P.; Quemada, H.D. How governmental regulation can help or hinder the integration of Bt crops within IPM programs. In Integration of Insect-Resistant Genetically Modified Crops withinIPM Programs; Romeis, J., Shelton, A.M., Kennedy, G.G., Eds.; Springer: New York, NY, USA, 2008; Volume 5, pp. 27–39. [Google Scholar]
- Van Rensburg, J.B.J. First report of field resistance by stem borer, Busseola fusca (Fuller) to Bt-transgenic maize. S. Afr. J. Plant Soil 2007, 24, 147–151. [Google Scholar]
- Carrière, Y.; Ellers-Kirk, C.; Sisterson, M.; Antilla, L.; Whitlow, M.; Dennehy, T.J.; Tabashnik, B.E. Long-term regional suppression of pink bollworm by Bacillus thuringiensis cotton. Proc. Nat. Acad. Sci. USA 2003, 100, 1519. [Google Scholar]
- Van Rie, J.; McGaughey, W.H.; Johnson, D.E.; Barnett, B.D.; Van Mellaert, H. Mechanism of insect resistance to the microbial insecticide Bacillus thuringiensis. Science 1990, 247, 72. [Google Scholar]
- Giles, K.L.; Hellmich, R.L.; Iverson, C.T.; Lewis, L.C. Effects of transgenic Bacillus thuringiensis maize grain on B. thuringiensis-susceptible Plodia interpunctella (Lepidoptera: Pyralidae). J. Econ. Entomol. 2000, 93, 1011–1016. [Google Scholar] [CrossRef]
- Sedlacek, J.D.; Komaravalli, S.R.; Hanley, A.M.; Price, B.D.; Davis, P.M. Life History Attributes of Indian Meal Moth (Lepidoptera: Pyralidae) and Angoumois Grain Moth (Lepidoptera: Gelechiidae) Reared on Transgenic Corn Kernels. J. Econ. Entomol. 2001, 94, 586–592. [Google Scholar] [CrossRef]
- Liu, Y.-B.; Tabashnik, B.E.; Dennehy, T.J.; Patin, A.L.; Bartlett, A.C. Development time and resistance to Bt crops. Nature 1999, 400, 519. [Google Scholar]
- Lopez, M.D.; Sumerford, D.V.; Lewis, L.C. Nosema pyrausta and Cry1Ab-incorporated diet led to decreased survival and developmental delays in European corn borer. Entomologia Experimentalis et Applicata 2010, 134, 146–153. [Google Scholar] [CrossRef]
- Gryspeirt, A.; Grégoire, J.-C. Effects of Two Varieties of Bacillus thuringiensis Maize on the Biology of Plodia Interpunctella. Toxins 2012, 4, 373–389. [Google Scholar] [CrossRef]
- Storer, N.P.; Van Duyn, J.W.; Kennedy, G.G. Life history traits of Helicoverpa zea (Lepidoptera: Noctuidae) on non-Bt and Bt transgenic corn hybrids in eastern North Carolina. J. Econ. Entomol. 2001, 94, 1268–1279. [Google Scholar] [CrossRef]
- Horner, T.A.; Dively, G.P.; Herbert, D.A. Effects of MON810 Bt field corn on adult emergence of Helicoverpa zea (Lepidoptera: Noctuidae). J. Econ. Entomol. 2003, 96, 925–930. [Google Scholar] [CrossRef]
- Peck, S.L.; Gould, F.; Ellner, S.P. Spread of resistance in spatially extended regions of transgenic cotton: implications for management of Heliothis virescens (Lepidoptera: Noctuidae). J. Econ. Entomol. 1999, 92, 1–16. [Google Scholar]
- Fabrick, J.A.; Forlow Jech, L.; Henneberry, T.J. Novel pink bollworm resistance to the Bt toxin Cry 1Ac: Effects on mating, oviposition, larval development and survival. J. Insect Sci. 2009, 9, 24. [Google Scholar]
- Shelton, A.M.; Tang, J.D.; Roush, R.T.; Metz, T.D.; Earle, E.D. Field tests on managing resistance to Bt-engineered plants. Nat. Biotech. 2000, 18, 339–342. [Google Scholar] [CrossRef]
- Gould, F. The evolutionary potential of crop pests. Am. Sci. 1991, 79, 496–507. [Google Scholar]
- Zhao, J.Z.; Cao, J.; Li, Y.; Collins, H.L.; Roush, R.T.; Earle, E.D.; Shelton, A.M. Transgenic plants expressing two Bacillus thuringiensis toxins delay insect resistance evolution. Nature Biotechnol. 2003, 21, 1493–1497. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gryspeirt, A.; Grégoire, J.-C. Effectiveness of the High Dose/Refuge Strategy for Managing Pest Resistance to Bacillus thuringiensis (Bt) Plants Expressing One or Two Toxins. Toxins 2012, 4, 810-835. https://doi.org/10.3390/toxins4100810
Gryspeirt A, Grégoire J-C. Effectiveness of the High Dose/Refuge Strategy for Managing Pest Resistance to Bacillus thuringiensis (Bt) Plants Expressing One or Two Toxins. Toxins. 2012; 4(10):810-835. https://doi.org/10.3390/toxins4100810
Chicago/Turabian StyleGryspeirt, Aiko, and Jean-Claude Grégoire. 2012. "Effectiveness of the High Dose/Refuge Strategy for Managing Pest Resistance to Bacillus thuringiensis (Bt) Plants Expressing One or Two Toxins" Toxins 4, no. 10: 810-835. https://doi.org/10.3390/toxins4100810
APA StyleGryspeirt, A., & Grégoire, J. -C. (2012). Effectiveness of the High Dose/Refuge Strategy for Managing Pest Resistance to Bacillus thuringiensis (Bt) Plants Expressing One or Two Toxins. Toxins, 4(10), 810-835. https://doi.org/10.3390/toxins4100810