Effects of Two Varieties of Bacillus thuringiensis Maize on the Biology of Plodia interpunctella

Abstract
:1. Introduction
2. Materials and Methods
2.1. Maize Varieties
2.2. Plodia interpunctella Strains and Rearing Conditions
2.3. Impact of Different Diets on the Biology of Plodia interpunctella
2.4. Choice Experiments: Larval Preference between Two Diets
2.5. Data Analysis
3. Results and Discussion
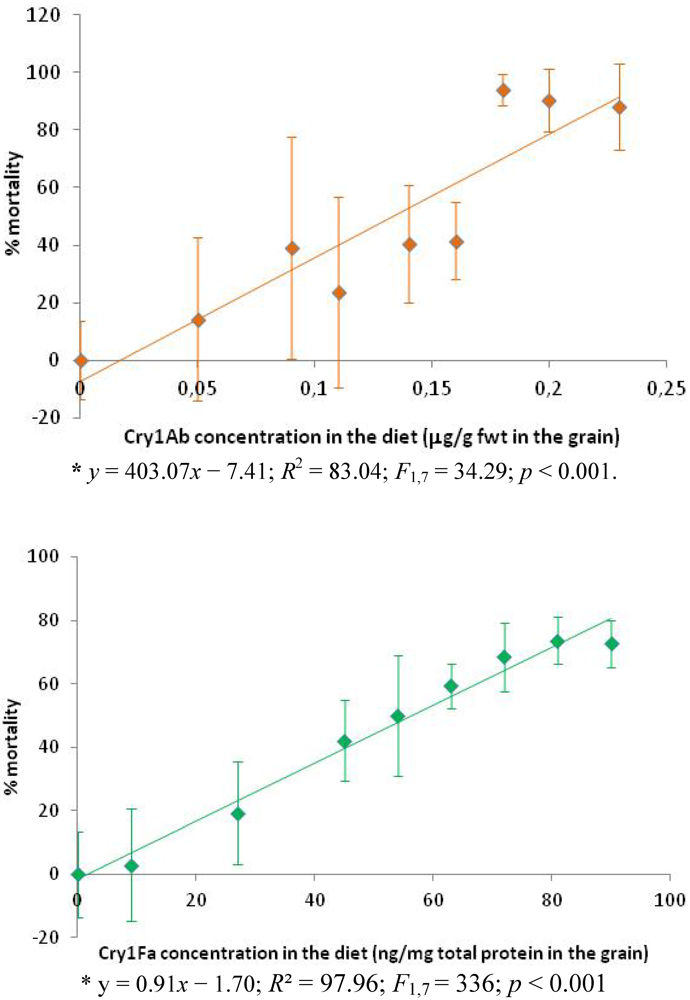
3.1. Susceptibility of Plodia interpunctella to the Cry Toxins


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rearing diet | n | ♀:♂ | χ² | df | p |
|---|---|---|---|---|---|
| Non-Bt maize diet | 152 | 0.45:0.55 | 1.68 | 1 | 0.194 |
| Standard wheat diet | 521 | 0.48:0.52 | 0.85 | 1 | 0.358 |
| Cry1Ab diet | 308 | 0.52:0.48 | 0.32 | 1 | 0.569 |
| Cry1Fa diet | 369 | 0.54:0.46 | 2.95 | 1 | 0.086 |
3.2. Choice Experiments: Larval Preference between Two Diets
| Homogeneity of the replicates | Comparison of the larval distribution | |||||||
|---|---|---|---|---|---|---|---|---|
| (%) | G-Test | df | p | G-Test | df | p | ||
| Larval diffusion | point of departure | 58.0 | 1.87 | 2 | 0.403 | 1.67 | 1 | 0.196 |
| adjacent zone | 42.0 | |||||||
| Control | non Bt zone | 47.0 | 3.02 | 4 | 0.406 | 0.48 | 1 | 0.486 |
| Non Bt zone | 53.0 | |||||||
| MON810 maize case | Cry1Ab zone | 52.8 | 4.86 | 4 | 0.301 | 0.45 | 1 | 0.501 |
| non Bt zone | 47.2 | |||||||
| TC 1507 maize case | Cry1Fa zone | 30.4 | 2.78 | 4 | 0.301 | 2.70 | 1 | <0.000 |
| non Bt zone | 69.6 | |||||||
3.3. Impact of the Diet on the Adult Weight
| Concentration in the rearing diet | Adult weight (mg) | Adult lifespan (day) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| N | Female | N | Male | N | Female | N | Male | ||
| Cry1Ab g/g fwt in the grain | 0 | 68 | 12.15 (3.35) | 84 | 8.45 (1.45) | 65 | 20.00 (4.82) | 83 | 20.00 (6.96) |
| 0.05 | 41 | 14.90 (2.20) | 31 | 9.02 (1.15) | 41 | 20.97 (3.92) | 31 | 21.07 (5.61) | |
| 0.09 | 28 | 14.17 (3.21) | 22 | 8.96 (0.85) | 28 | 20.91 (4.82) | 22 | 20.91 (5.90) | |
| 0.11 | 30 | 13.63 (3.21) | 31 | 9.38 (1.81) | 30 | 20.48 (4.27) | 31 | 20.58 (3.68) | |
| 0.14 | 25 | 15.00 (3.15) | 19 | 9.26 (1.74) | 25 | 20.41 (4.60) | 17 | 20.41 (3.84) | |
| 0.16 | 15 | 15.24 (2.95) | 28 | 9.98 (2.52) | 14 | 17.85 (5.36) | 26 | 17.85 (5.86) | |
| 0.18 | 11 | 16.08 (2.64) | 9 | 7.44 (1.66) | 11 | 17.00 (2.71) | 9 | 17.00 (4.58) | |
| Cry1Fa ng/mg tot prot in the grain | 0 | 68 | 12.15 (3.35) | 84 | 8.65 (1.55) | 65 | 20.20 (4.82) | 83 | 20.20 (6.96) |
| 9 | 44 | 9.17 (2.23) | 41 | 5.96 (1.38) | 44 | 21.22 (6.47) | 41 | 21.22 (6.27) | |
| 27 | 46 | 7.60 (2.13) | 28 | 4.39 (1.29) | 46 | 18.41 (5.73) | 27 | 18.41 (6.86) | |
| 45 | 34 | 7.94 (2.63) | 19 | 4.80 (1.37) | 34 | 20.26 (5.68) | 19 | 20.36 (6.43) | |
| 54 | 25 | 6.65 (1.75) | 22 | 4.43 (1.45) | 25 | 17.77 (5.38) | 22 | 17.87 (6.25) | |
| 63 | 12 | 7.69 (1.33) | 18 | 4.85 (1.23) | 12 | 19.17 (5.59) | 18 | 19.27 (6.58) | |
| 72 | 15 | 8.63 (1.92) | 15 | 5.86 (1.03) | 15 | 22.93 (5.97) | 15 | 22.93 (4.50) | |
| 81 | 8 | 7.76 (2.40) | 10 | 5.10 (1.12) | 8 | 21.70 (5.71) | 10 | 21.70 (5.52) | |
| 90 | 10 | 9.22 (3.18) | 10 | 5.92 (0.90) | 10 | 23.80 (7.54) | 10 | 23.80 (4.05) | |
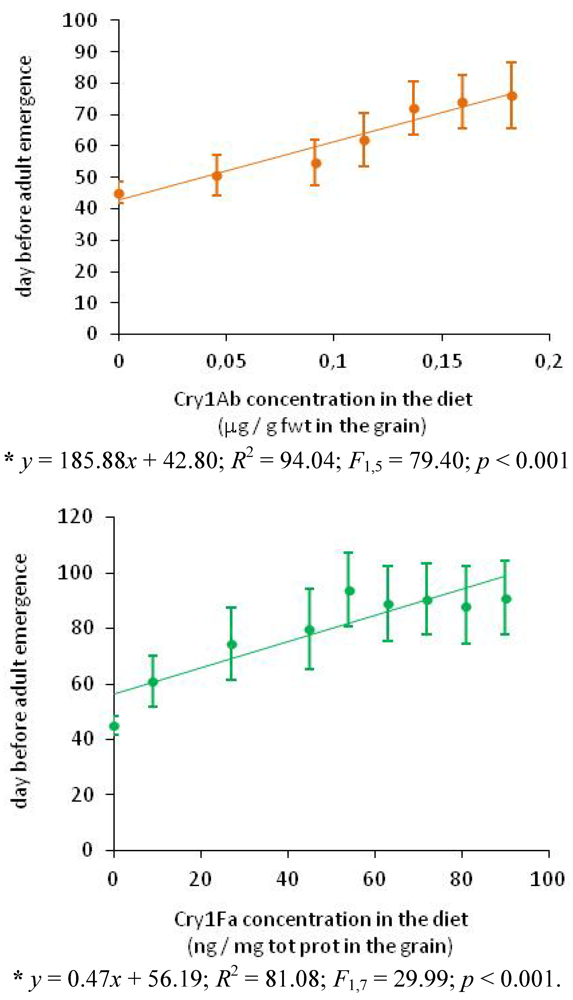
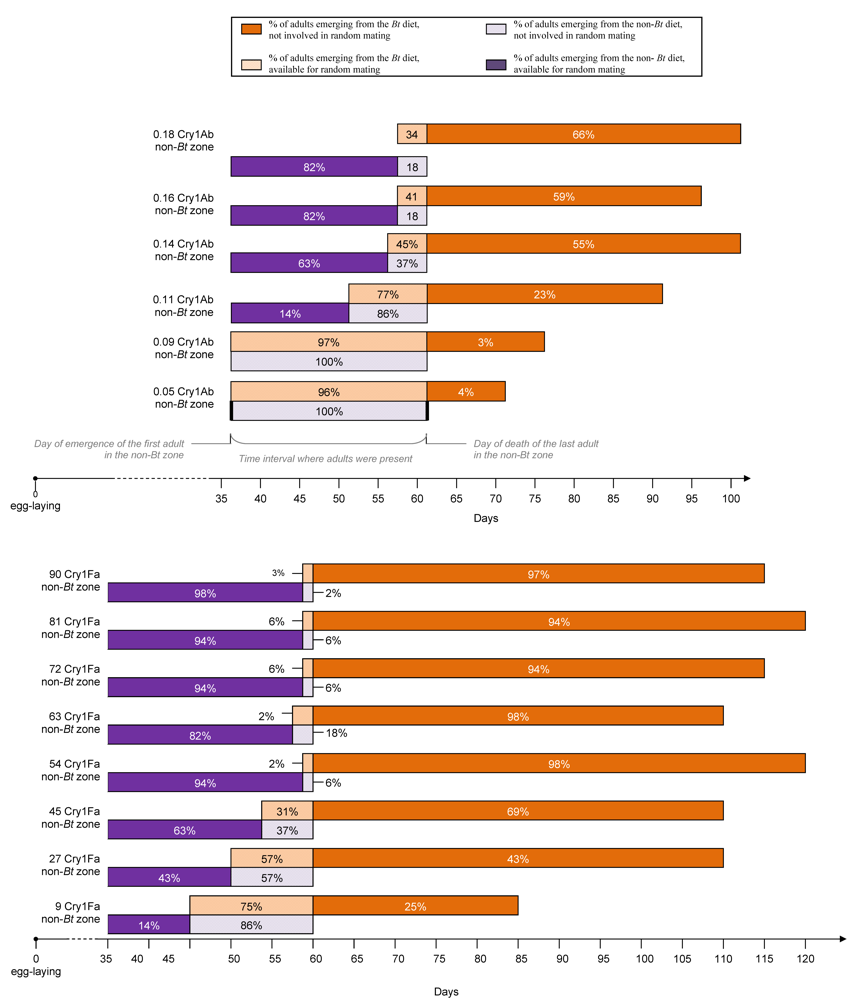
3.4. Impact of Bt Maize on the Life Cycle of Plodia interpunctella (from Egg to Adult Emergence-Adult Lifespan)

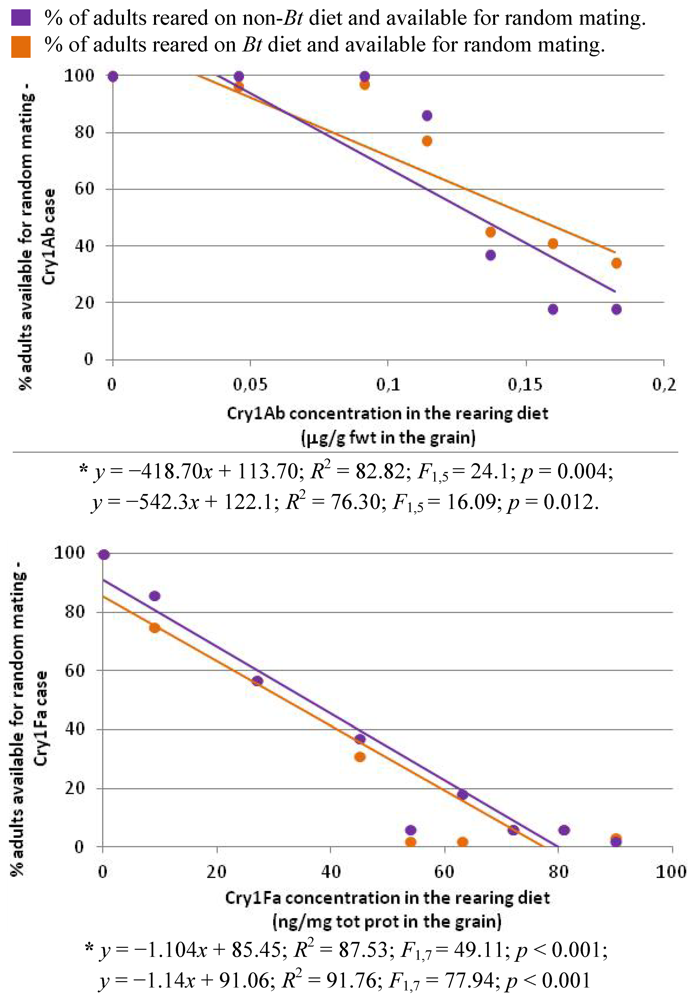
3.5. Impact of the Bt Maize on the Percentage of Adults Available for Random Mating


Acknowledgments
Conflict of Interest
References
- Vaeck, M.; Reynaerts, A.; Hofte, H.; Jansens, S.; De Beuckeleer, M.; Dean, C.; Zabeau, M.; Montagu, M.V.; Leemans, J. Transgenic plants protected from insect attack. Nature 1987, 328, 33–37. [Google Scholar]
- Perlak, F.J.; Fuchs, R.L.; Dean, D.A.; McPherson, S.L.; Fischhoff, D.A. Modification of the coding sequence enhances plant expression of insect control protein genes. Proc. Natl. Acad. Sci. USA 1991, 88, 3324–3328. [Google Scholar]
- Gill, S.S.; Cowles, E.A.; Pietrantonio, P.V. The mode of action of Bacillus thuringiensis endotoxins. Annu. Rev. Entomol. 1992, 37, 615–634. [Google Scholar] [CrossRef]
- Ferré, J.; Escriche, B.; Bel, Y.; Rie, J. Biochemistry and genetics of insect resistance to Bacillus thuringiensis insecticidal crystal proteins. FEMS Microbiol. Lett. 1995, 132, 1–7. [Google Scholar]
- Alyokhin, A.; Ferro, D. Relative fitness of Colorado potato beetle (Coleoptera:Chrysomelidae) resistant and susceptible to the Bacillus thuringiensis Cry3A toxin. J. Econ. Entomol. 1999, 92, 510–515. [Google Scholar]
- Gonzalez-Cabrera, J.; Herrero, S.; Sayyed, A.H.; Escriche, B.; Liu, Y.B.; Meyer, S.K.; Wright, D.J.; Tabashnik, B.E.; Ferré, J. Variation in susceptibility to Bacillus thuringiensis toxins among unselected strains of Plutella xylostella. Appl. Environ. Microbiol. 2001, 67, 4610. [Google Scholar]
- Giles, K.L.; Hellmich, R.L.; Iverson, C.T.; Lewis, L.C. Effects of transgenic Bacillus thuringiensis maize grain on B. thuringiensis-susceptible Plodia interpunctella (Lepidoptera:Pyralidae). J. Econ. Entomol. 2000, 93, 1011–1016. [Google Scholar] [CrossRef]
- Bird, L.J.; Akhurst, R.J. Relative fitness of Cry1A-resistant and -susceptible Helicoverpa armigera (Lepidoptera:Noctuidae) on conventional and transgenic cotton. J. Econ. Entomol. 2004, 97, 1699–1709. [Google Scholar] [CrossRef]
- Lopez, M.D.; Sumerford, D.V.; Lewis, L.C. Nosema pyrausta and Cry1Ab‐incorporated diet led to decreased survival and developmental delays in European corn borer. Entomol. Exp. Appl. 2010, 134, 146–153. [Google Scholar] [CrossRef]
- Oppert, B. Effects of Cry1F and Cry34Ab1/35Ab1 on storage pests. J. Stored Prod. Res. 2010, 46, 143–148. [Google Scholar] [CrossRef]
- Koziel, M.G.; Beland, G.L.; Bowman, C.; Carozzi, N.B.; Crenshaw, R.; Crossland, L.; Dawson, J.; Desai, N.; Hill, M.; Kadwell, S.; et al. Field performance of elite transgenic maize plants expressing an insecticidal protein derived from Bacillus thuringiensis. Nat. Biotechnol. 1993, 11, 194–200. [Google Scholar]
- Sedlacek, J.D.; Komaravalli, S.R.; Hanley, A.M.; Price, B.D.; Davis, P.M. Life history attributes of Indian meal moth (Lepidoptera:Pyralidae) and angoumois grain moth (Lepidoptera:Gelechiidae) reared on transgenic corn kernels. J. Econ. Entomol. 2001, 94, 586–592. [Google Scholar] [CrossRef]
- McGaughey, W.H. Insect resistance to the biological insecticide Bacillus thuringiensis. Science 1985, 229, 193. [Google Scholar]
- McGaughey, W.H. Effects of larval age on the susceptibility of almond moths and indianmeal moths to Bacillus thuringiensis. J. Econ. Entomol. 1978, 71, 923–925. [Google Scholar]
- McGaughey, W.H.; Johnson, D.E. Toxicity of different serotypes and toxins of Bacillus thuringiensis to resistant and susceptible Indianmeal moths (Lepidoptera:Pyralidae). J. Econ. Entomol. 1987, 80, 1122–1126. [Google Scholar]
- Johnson, D.E.; McGaughey, W.H.; Barnett, B.D. Small scale bioassay for the determination of Bacillus thuringiensis toxicity toward Plodia interpunctella. J. Invertebr. Pathol. 1991, 57, 159–165. [Google Scholar] [CrossRef]
- Gould, F. Sustainability of transgenic insecticidal cultivars: Integrating pest genetics and ecology. Annu. Rev. Entomol. 1998, 43, 701–726. [Google Scholar] [CrossRef]
- McGaughey, W.H.; Johnson, D.E. Indianmeal Moth (Lepidoptera:Pyralidae) resistance to different strains and mixtures of Bacillus thuringiensis. J. Econ. Entomol. 1992, 85, 1594–1600. [Google Scholar]
- Oppert, B.; Kramer, K.J.; Beeman, R.W.; Johnson, D.; McGaughey, W.H. Proteinase-Mediated insect resistance to Bacillus thuringiensis toxins. J. Biol. Chem. 1997, 272, 23473–23476. [Google Scholar]
- Anonymous Safety assessment of Yieldgard insect-protected corn Event MON810. Available online: http://www.monsanto.com/products/Documents/safety-summaries/yieldgard_corn_pss.pdf (accessed on May 23 2012).
- Grassé, P. Traité de Zoologie: Anatomie, Systématique, Biologie; Masson: Paris, France, 1977; pp. 512–523. [Google Scholar]
- Anonymous The R Project for Statistical Computing. Available online: http://www.r-project.org/ (accessed on 23 May 2012).
- Horner, T.A.; Dively, G.P.; Herbert, D.A. Development, survival and fitness performance of Helicoverpa zea (Lepidoptera:Noctuidae) in MON810 Bt field corn. J. Econ. Entomol. 2003, 96, 914–924. [Google Scholar] [CrossRef]
- Khalique, F.; Ahmed, K. Impact of Bacillus thuringiensis subsp. kurstaki on biology of Helicoverpa zea armigera. Pak. J. Biol. Sci. 2003, 6, 615–621. [Google Scholar] [CrossRef]
- Liu, Y.-B.; Tabashnik, B.E.; Dennehy, T.J.; Patin, A.L.; Sims, M.A.; Meyer, S.K.; Carrière, Y. Effects of Bt cotton and Cry1Ac toxin on survival and development of pink Bollworm (Lepidoptera:Gelechiidae). J. Econ. Entomol. 2001, 94, 1237–1242. [Google Scholar]
- Gierl, A.; Frey, M. Evolution of benzoxazinone biosynthesis and indole production in maize. Planta 2001, 213, 493–498. [Google Scholar]
- Gould, F.; Anderson, A. Effects of Bacillus thuringiensis and HD-73 Delta-Endotoxin on growth, behavior, and fitness of susceptible and toxin-adapted strains of Heliothis virescens (Lepidoptera:Noctuidae. Environ. Entomol. 1991, 20, 30–38. [Google Scholar]
- Parker, C.D., Jr.; Luttrell, R.G. Interplant movement of Heliothis virescens (Lepidoptera:Noctuidae) larvae in pure and mixed plantings of cotton with and without expression of the Cry1Ac delta-endotoxin protein of Bacillus thuringiensis Berliner. J. Econ. Entomol. 1999, 92, 837–845. [Google Scholar]
- Goldstein, J.A.; Mason, C.E.; Pesek, J. Dispersal and movement behavior of neonate European corn borer (Lepidoptera:Crambidae) on non-Bt and transgenic Bt corn. J. Econ. Entomol. 2010, 103, 331–339. [Google Scholar]
- Sutherland, P.; Harris, M.; Markwick, N. Effects of starvation and the Bacillus thuringiensis endotoxin CrylAc on the midgut cells, feeding behavior, and growth of lightbrown apple moth larva. Ann. Entomol. Soc. Am. 2003, 96, 250–264. [Google Scholar]
- Harris, M.O.; Mafile’o, F.; Dhana, S. Behavioral responses of lightbrown apple moth neonate larvae on diets containing Bacillus thuringiensis formulations or endotoxins. Entomol. Exp. Appl. 1997, 84, 207–219. [Google Scholar]
- Harris, M.O.; Markwick, N.; Sandanayake, M. Is resistance to Bacillus thuringiensis endotoxin Cry1Ac associated with a change in the behavior of light brown apple moth larvae (Lepidoptera:Tortricidae)? J. Econ. Entomol. 2006, 99, 508–518. [Google Scholar]
- Honěk, A. Intraspecific Variation in Body Size and Fecundity in Insects: A General Relationship. Oikos 1993, 66, 483–492. [Google Scholar]
- Abdel-Razek, A.; Salama, H.; White, N.; Morris, O. Effect of Bacillus thuringiensis on feeding and energy use by Plodia interpunctella (Lepidoptera:Pyralidae) and Tribolium castaneum (Coleoptera:Tenebrionidae). Can. Entomol. 1999, 131, 433–440. [Google Scholar]
- Sayyed, A.H.; Cerda, H.; Wright, D.J. Could Bt transgenic crops have nutritionally favourable effects on resistant insects? Ecol. Lett. 2003, 6, 167–169. [Google Scholar]
- Tabashnik, B.E.; Carrière, Y. Bt transgenic crops do not have favorable effects on resistant insects. J. Insect Sci. 2004, 4, 3. [Google Scholar]
- Storer, N.P.; Van Duyn, J.W.; Kennedy, G.G. Life history traits of Helicoverpa zea (Lepidoptera:Noctuidae) on non-Bt and Bt transgenic corn hybrids in eastern North Carolina. J. Econ. Entomol. 2001, 94, 1268–1279. [Google Scholar]
- Caprio, M.A. Evaluating resistance management strategies for multiple toxins in the presence of external refuges. J. Econ. Entomol. 1998, 91, 1021–1031. [Google Scholar]
- Peck, S.L.; Gould, F.; Ellner, S.P. Spread of resistance in spatially extended regions of transgenic cotton: Implications for management of Heliothis virescens (Lepidoptera:Noctuidae). J. Econ. Entomol. 1999, 92, 1–16. [Google Scholar]
- Horner, T.A.; Dively, G.P.; Herbert, D.A. Effects of MON810 Bt field corn on adult emergence of Helicoverpa zea (Lepidoptera:Noctuidae). J. Econ. Entomol. 2003, 96, 925–930. [Google Scholar] [CrossRef]
- Huang, F.; Subramanyam, B. Effects of delayed mating on reproductive performance of Plodia interpunctella (Hübner)(Lepidoptera:Pyralidae). J. Stored Prod. Res. 2003, 39, 53–63. [Google Scholar] [CrossRef]
- Johnson, J.; Wofford, P.; Whitehand, L. Effect of diet and temperature on development rates, survival, and reproduction of the indianmeal moth (Lepidoptera, Pyralida). J. Econ. Entomol. 1992, 85, 561–566. [Google Scholar]
- Mohandass, S.; Arthur, F.H.; Zhu, K.Y.; Throne, J.E. Biology and management of Plodia interpunctella (Lepidoptera:Pyralidae) in stored products. J. Stored Prod. Res. 2007, 43, 302–311. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gryspeirt, A.; Grégoire, J.-C. Effects of Two Varieties of Bacillus thuringiensis Maize on the Biology of Plodia interpunctella. Toxins 2012, 4, 373-389. https://doi.org/10.3390/toxins4050373
Gryspeirt A, Grégoire J-C. Effects of Two Varieties of Bacillus thuringiensis Maize on the Biology of Plodia interpunctella. Toxins. 2012; 4(5):373-389. https://doi.org/10.3390/toxins4050373
Chicago/Turabian StyleGryspeirt, Aiko, and Jean-Claude Grégoire. 2012. "Effects of Two Varieties of Bacillus thuringiensis Maize on the Biology of Plodia interpunctella" Toxins 4, no. 5: 373-389. https://doi.org/10.3390/toxins4050373
APA StyleGryspeirt, A., & Grégoire, J. -C. (2012). Effects of Two Varieties of Bacillus thuringiensis Maize on the Biology of Plodia interpunctella. Toxins, 4(5), 373-389. https://doi.org/10.3390/toxins4050373