Chemical Compounds Toxic to Invertebrates Isolated from Marine Cyanobacteria of Potential Relevance to the Agricultural Industry


Abstract
:1. Introduction


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
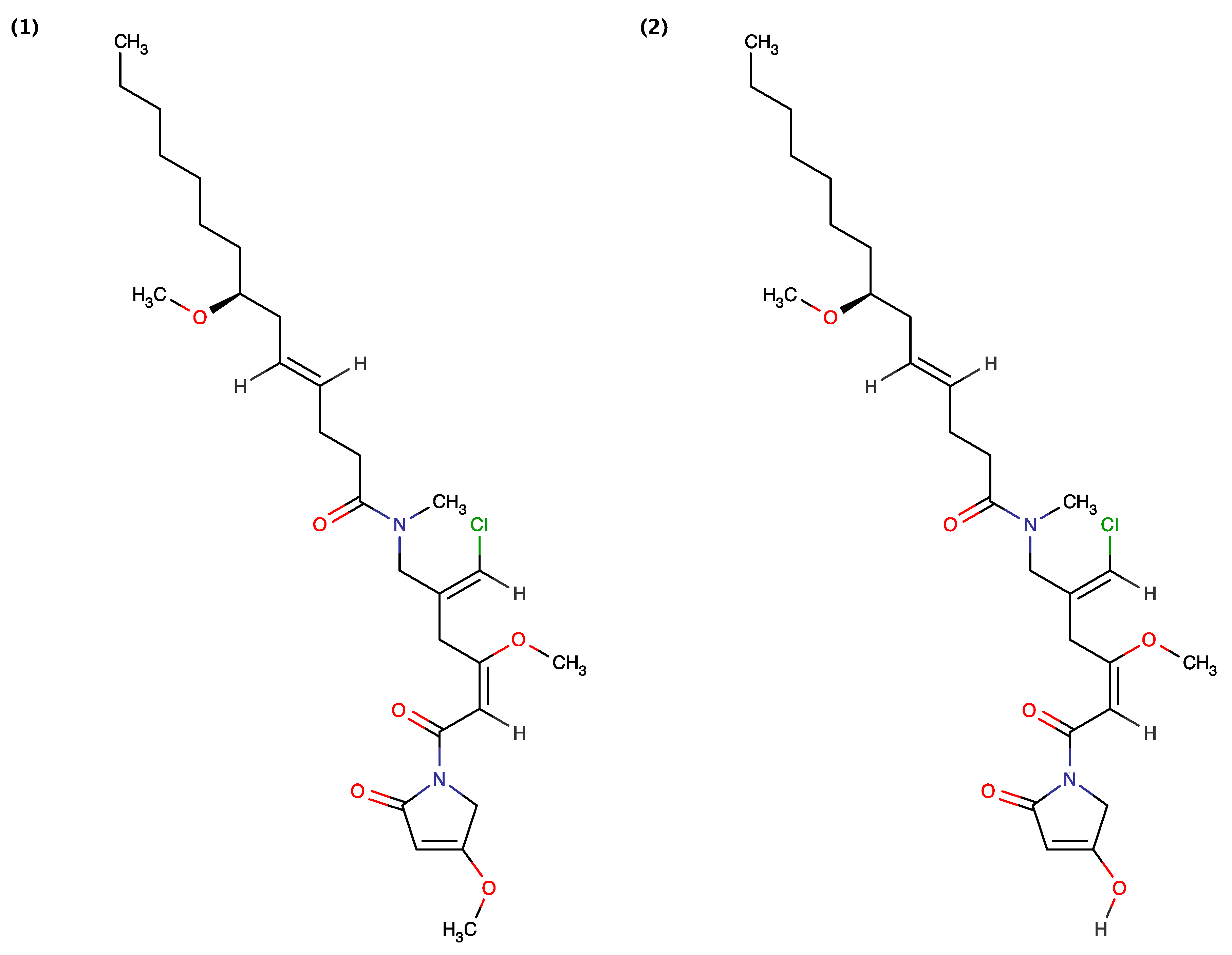{kind=link}
{kind=link}
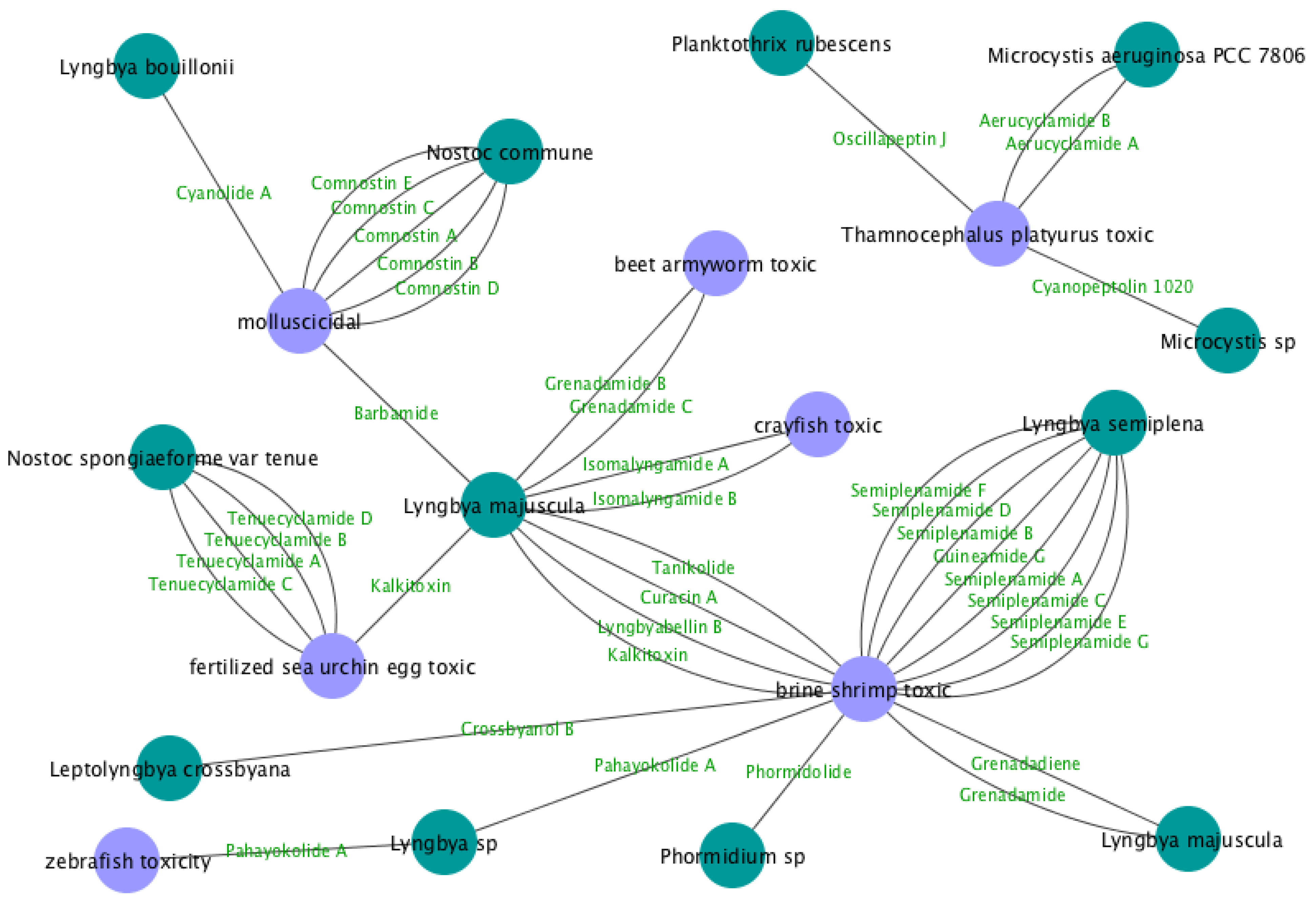
| Compound | Species | Biological Activity | References |
|---|---|---|---|
| Grenadamide B | Lyngbya majuscula | beet armyworm toxic | [6] |
| Grenadamide C | Lyngbya majuscula | beet armyworm toxic | [6] |
| Crossbyanol B | Leptolyngbya crossbyana | brine shrimp toxic | [7] |
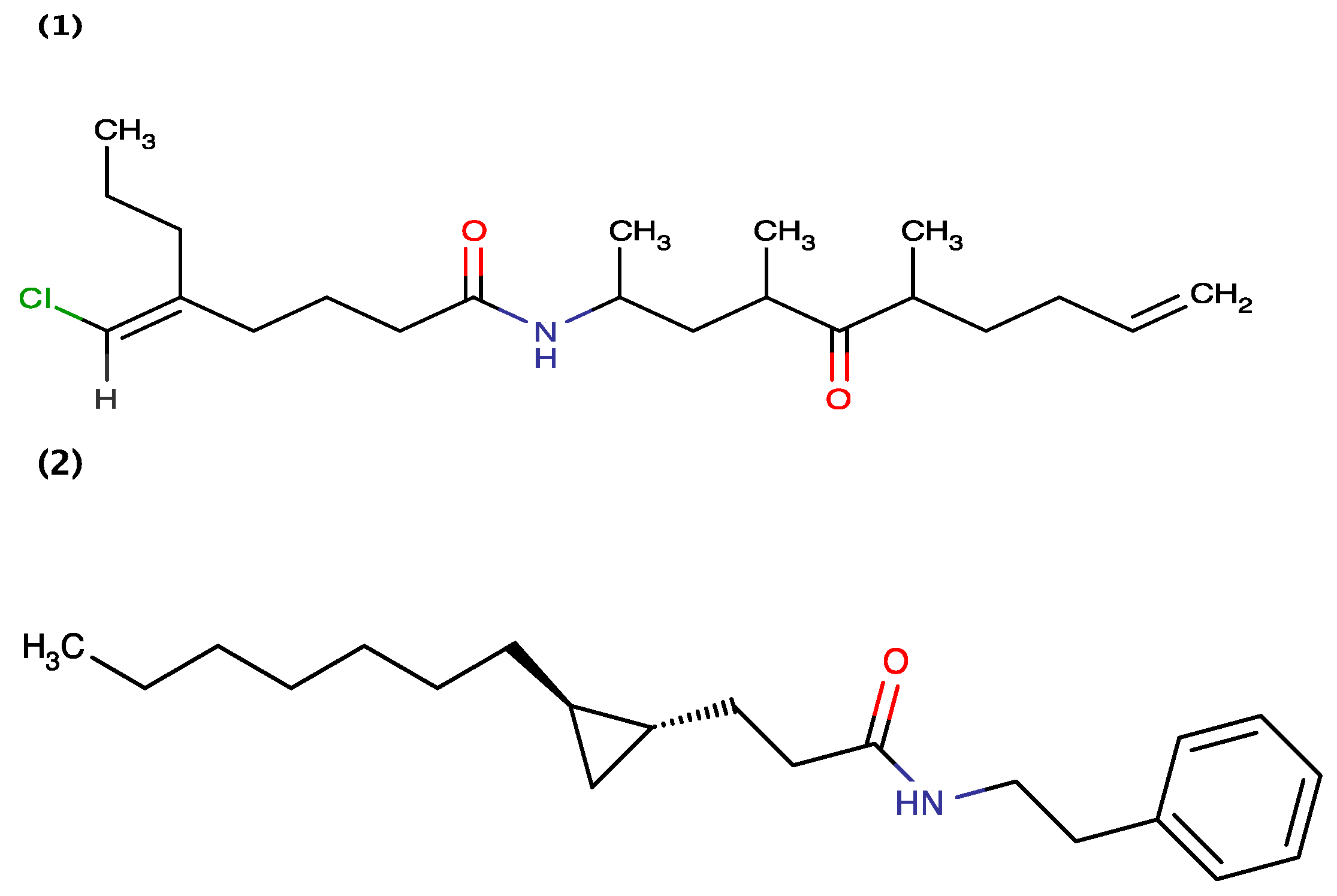
| Curacin A | Lyngbya majuscula | brine shrimp toxic | [8] |
| Grenadadiene | Lyngbya majuscula | brine shrimp toxic | [9] |
| Grenadamide | Lyngbya majuscula | brine shrimp toxic | [9] |
| Guineamide G | Lyngbya semiplena | brine shrimp toxic | [10] |
| Lyngbyabellin B | Lyngbya majuscula | brine shrimp toxic | [11] |
| Phormidolide | Phormidium sp. | brine shrimp toxic | [12] |
| Semiplenamide A | Lyngbya semiplena | brine shrimp toxic | [13] |
| Semiplenamide B | Lyngbya semiplena | brine shrimp toxic | [13] |
| Semiplenamide C | Lyngbya semiplena | brine shrimp toxic | [13] |
| Semiplenamide D | Lyngbya semiplena | brine shrimp toxic | [13] |
| Semiplenamide E | Lyngbya semiplena | brine shrimp toxic | [13] |
| Semiplenamide F | Lyngbya semiplena | brine shrimp toxic | [13] |
| Semiplenamide G | Lyngbya semiplena | brine shrimp toxic | [13] |
| Tanikolide | Lyngbya majuscula | brine shrimp toxic | [14] |
| Kalkitoxin | Lyngbya majuscula | brine shrimp toxic; fertilized sea urchin egg toxic | [15] |
| Pahayokolide A | Lyngbya sp. | brine shrimp toxic; zebrafish toxicity | [16] |
| Isomalyngamide A | Lyngbya majuscula | crayfish toxic | [17] |
| Isomalyngamide B | Lyngbya majuscula | crayfish toxic | [17] |
| Tenuecyclamide A | Nostoc spongiaeforme var. tenue | fertilized sea urchin egg toxic | [18] |
| Tenuecyclamide B | Nostoc spongiaeforme var. tenue | fertilized sea urchin egg toxic | [18] |
| Tenuecyclamide C | Nostoc spongiaeforme var. tenue | fertilized sea urchin egg toxic | [18] |
| Tenuecyclamide D | Nostoc spongiaeforme var. tenue | fertilized sea urchin egg toxic | [18] |
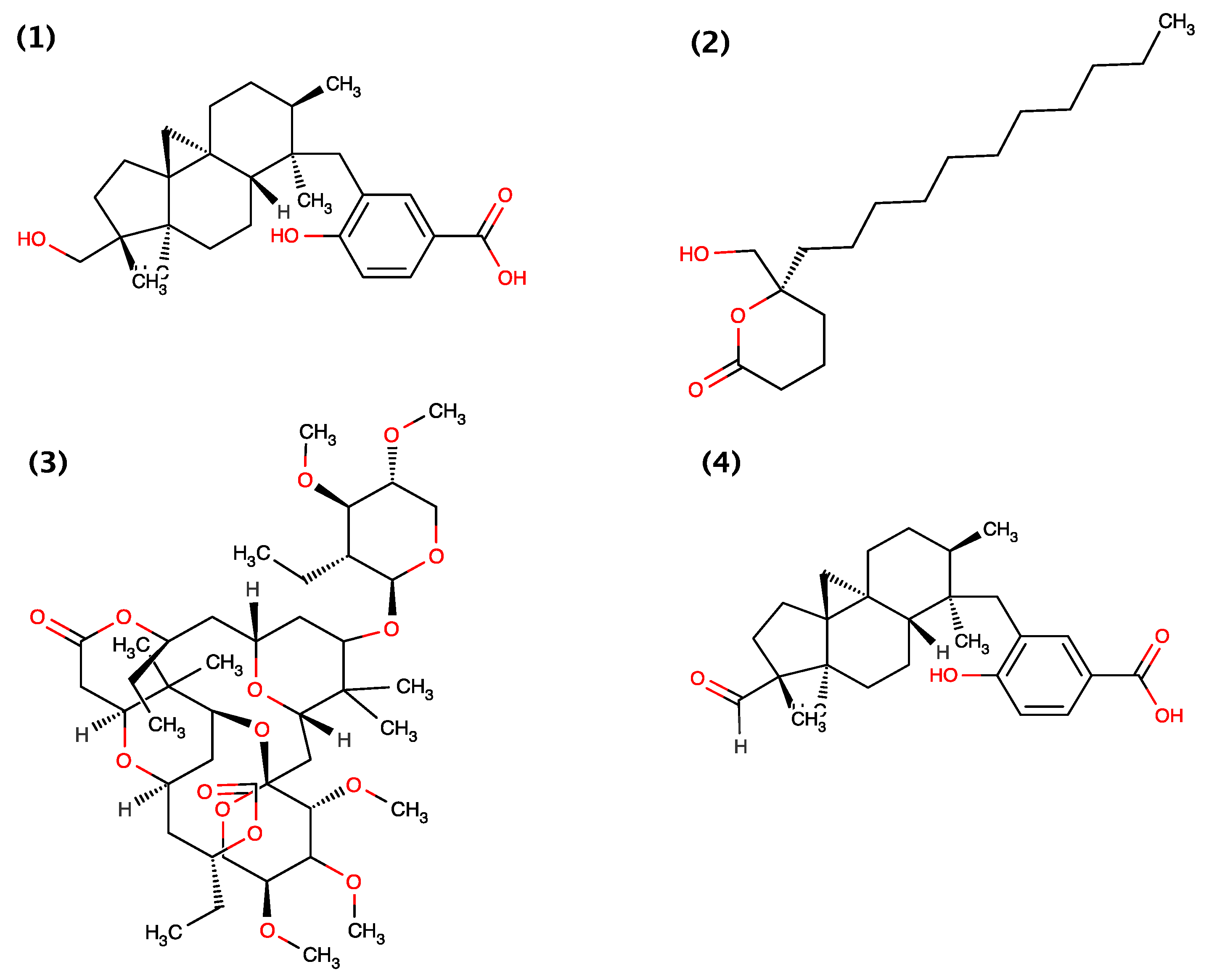
| Barbamide | Lyngbya majuscula | molluscicidal | [19] |
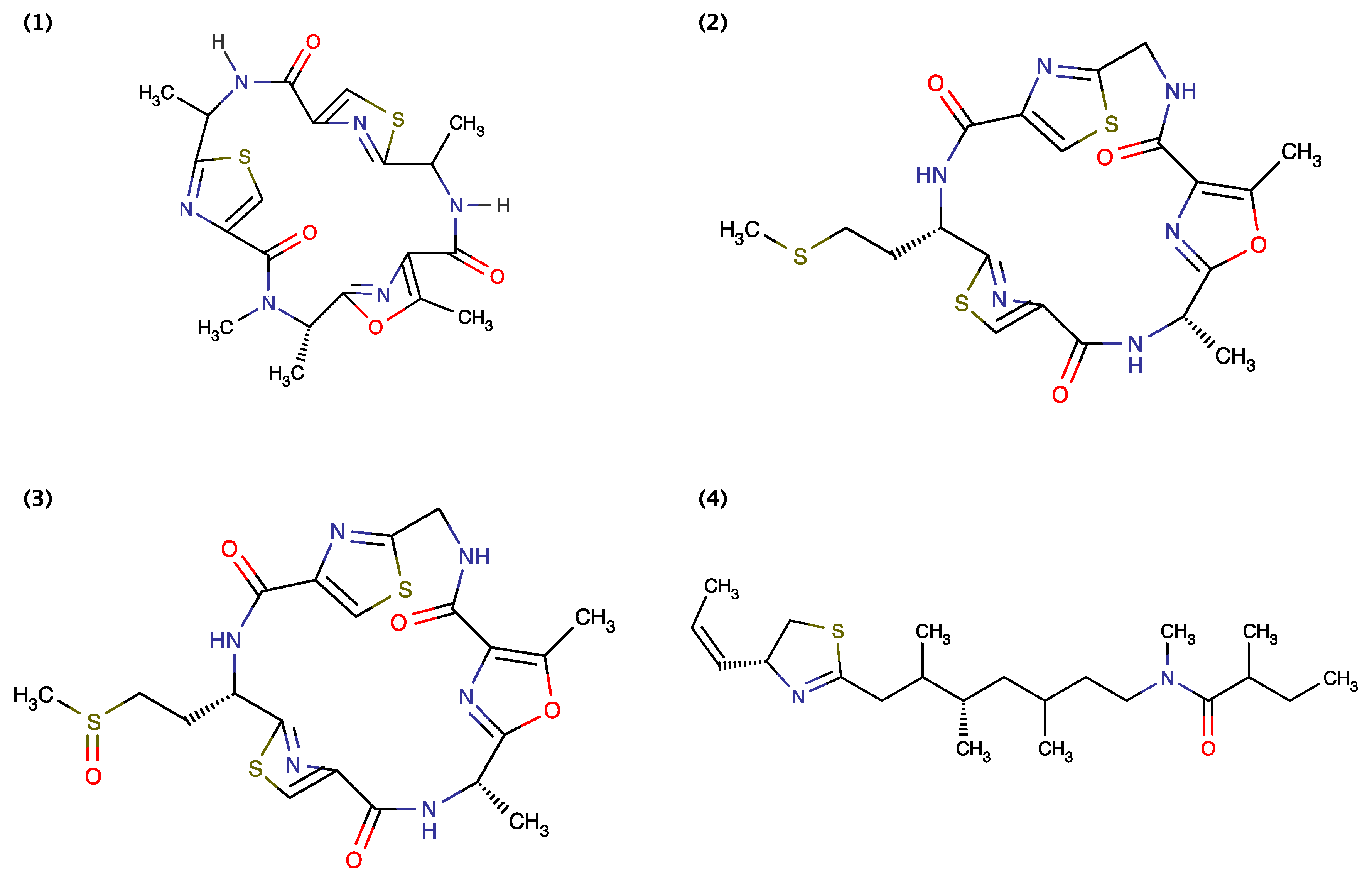
| Comnostin A | Nostoc commune | molluscicidal | [20] |
| Comnostin B | Nostoc commune | molluscicidal | [20] |
| Comnostin C | Nostoc commune | molluscicidal | [20] |
| Comnostin D | Nostoc commune | molluscicidal | [20] |
| Comnostin E | Nostoc commune | molluscicidal | [20] |
| Cyanolide A | Lyngbya bouillonii | molluscicidal | [21] |
| Aerucyclamide A | Microcystis aeruginosa PCC 7806 | Thamnocephalus platyurus toxic | [22] |
| Aerucyclamide B | Microcystis aeruginosa PCC 7806 | Thamnocephalus platyurus toxic | [22] |
| Cyanopeptolin 1020 | Microcystis sp. | Thamnocephalus platyurus toxic | [23] |
| Oscillapeptin J | Planktothrix rubescens | Thamnocephalus platyurus toxic | [24] |
2. Invertebrate Toxicity Related Research
3. Compounds that Exhibit Toxicity against Invertebrates
3.1. Compounds Toxic to Molluscs

3.2. Compounds Toxic to Beet Armyworms

3.3. Compounds Toxic to Fertilized Sea Urchin Eggs

3.4. Compounds Toxic to Brine Shrimp

3.5. Compounds Toxic to Beavertail Fairy Shrimp

3.6. Compounds Toxic to Crayfish

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Grains Research and Development Corporation. Snail management fact sheet. Available online: http://www.grdc.com.au/~/media/4492614E5B6B4540B4E01A43AAD97889.pdf (accessed on 30 May 2013).
- Grains Research and Development Corporation. Strategic research & development plan 2012–17. Available online: http://strategicplan2012.grdc.com.au/pdf/GRDC_strategic_plan_2012-17.pdf (accessed on 30 May 2013).
- Hayes, G. Nailing Snails—Practical Prevention and Control Measures. Available online: http://www.grdc.com.au/Research-and-Development/GRDC-Update-Papers/2013/02/Nailing-snails-Practical-prevention-and-control-measures (accessed on 30 May 2013).
- Grains Research and Development Corporation. Bash’em Burn’Em Bait’Em: Integrated Snail Management in Crops and Pastures 2010. Available online: https://www.grdc.com.au/uploads/documents/Snails%20BBB.pdf (accessed on 30 May 2013).
- Young, C.L.; Armstrong, G.D. Slugs, Snails and Iron Based Baits: An Increasing Problem and a Low Toxic Specific Action Solution. Available online: http://www.regional.org.au/au/asa/2001/6/c/young.htm (accessed on 30 May 2013).
- Jimenez, J.I.; Vansach, T.; Yoshida, W.Y.; Sakamoto, B.; Porzgen, P.; Horgen, F.D. Halogenated fatty acid amides and cyclic depsipeptides from an eastern caribbean collection of the cyanobacterium lyngbya majuscula. J. Nat. Prod. 2009, 72, 1573–1578. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Engene, N.; Smith, J.E.; Preskitt, L.B.; Gerwick, W.H. Crossbyanols A–D, toxic brominated polyphenyl ethers from the hawai’ian bloom-forming cyanobacterium leptolyngbya crossbyana. J. Nat. Prod. 2010, 73, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H.; Proteau, P.J.; Nagle, D.G.; Hamel, E.; Blokhin, A.; Slate, D.L. Structure of curacin A, a novel antimitotic, antiproliferative, and brine shrimp toxic natural product from the marine cyanobacterium lyngbya majuscula. J. Org. Chem. 1994, 59, 1243–1245. [Google Scholar] [CrossRef]
- Sitachitta, N.; Gerwick, W.H. Grenadadiene and grenadamide, cyclopropyl-containing fatty acid metabolites from the marine cyanobacterium lyngbya majuscula. J. Nat. Prod. 1998, 61, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Gross, H.; McPhail, K.L.; Goeger, D.; Maier, C.S.; Gerwick, W.H. Wewakamide A and guineamide G, cyclic depsipeptides from the marine cyanobacteria lyngbya semiplena and lyngbya majuscula. J. Microbiol. Biotechnol. 2011, 21, 930–936. [Google Scholar] [CrossRef] [PubMed]
- Milligan, K.E.; Marquez, B.L.; Williamson, R.T.; Gerwick, W.H. Lyngbyabellin B, a toxic and antifungal secondary metabolite from the marine cyanobacterium lyngbya majuscula. J. Nat. Prod. 2000, 63, 1440–1443. [Google Scholar] [CrossRef] [PubMed]
- Williamson, R.T.; Boulanger, A.; Vulpanovici, A.; Roberts, M.A.; Gerwick, W.H. Structure and absolute stereochemistry of phormidolide, a new toxic metabolite from the marine cyanobacterium phormidium sp. J. Org. Chem. 2002, 67, 7927–7936. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; McPhail, K.L.; Ligresti, A.; di Marzo, V.; Gerwick, W.H. Semiplenamides A–G, fatty acid amides from a papua new guinea collection of the marine cyanobacterium lyngbya semiplena. J. Nat. Prod. 2003, 66, 1364–1368. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.P.; Milligan, K.E.; Gerwick, W.H. Tanikolide, a toxic and antifungal lactone from the marine cyanobacterium lyngbya majuscula. J. Nat. Prod. 1999, 62, 1333–1335. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Okino, T.; Nogle, L.M.; Marquez, B.L.; Williamson, R.T.; Sitachitta, N.; Berman, F.W.; Murray, T.F.; McGough, K.; Jacobs, R.; et al. Structure, synthesis, and biological properties of kalkitoxin, a novel neurotoxin from the marine cyanobacterium lyngbya majuscula. J. Am. Chem. Soc. 2000, 122, 12041–12042. [Google Scholar] [CrossRef]
- An, T.; Kumar, T.K.; Wang, M.; Liu, L.; Lay, J.O., Jr.; Liyanage, R.; Berry, J.; Gantar, M.; Marks, V.; Gawley, R.E.; et al. Structures of pahayokolides A and B, cyclic peptides from a lyngbya sp. J. Nat. Prod. 2007, 70, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Kan, Y.; Sakamoto, B.; Fujita, T.; Nagai, H. New malyngamides from the hawaiian cyanobacterium lyngbya majuscula. J. Nat. Prod. 2000, 63, 1599–1602. [Google Scholar] [CrossRef] [PubMed]
- Banker, R.; Carmeli, S. Tenuecyclamides A–D, cyclic hexapeptides from the cyanobacterium nostoc spongiaeforme var. Tenue. J. Nat. Prod. 1998, 61, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Orjala, J.; Gerwick, W.H. Barbamide, a chlorinated metabolite with molluscicidal activity from the caribbean cyanobacterium lyngbya majuscula. J. Nat. Prod. 1996, 59, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Jaki, B.; Orjala, J.; Heilmann, J.; Linden, A.; Vogler, B.; Sticher, O. Novel extracellular diterpenoids with biological activity from the cyanobacterium nostoc commune. J. Nat. Prod. 2000, 63, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.R.; McCue, C.F.; Gerwick, W.H. Cyanolide A, a glycosidic macrolide with potent molluscicidal activity from the papua new guinea cyanobacterium lyngbya bouillonii. J. Nat. Prod. 2010, 73, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Portmann, C.; Blom, J.F.; Gademann, K.; Juttner, F. Aerucyclamides A and B: Isolation and synthesis of toxic ribosomal heterocyclic peptides from the cyanobacterium microcystis aeruginosa pcc 7806. J. Nat. Prod. 2008, 71, 1193–1196. [Google Scholar] [CrossRef] [PubMed]
- Gademann, K.; Portmann, C.; Blom, J.F.; Zeder, M.; Juttner, F. Multiple toxin production in the cyanobacterium microcystis: Isolation of the toxic protease inhibitor cyanopeptolin 1020. J. Nat. Prod. 2010, 73, 980–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blom, J.F.; Bister, B.; Bischoff, D.; Nicholson, G.; Jung, G.; Sussmuth, R.D.; Juttner, F. Oscillapeptin J, a new grazer toxin of the freshwater cyanobacterium planktothrix rubescens. J. Nat. Prod. 2003, 66, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Mas-Coma, M.S.; Esteban, J.G.; Bargues, M.D. Epidemiology of human fascioliasis: A review and proposed new classification. Bull. World Health Organ. 1999, 77, 340–346. [Google Scholar] [PubMed]
- Chen, J.X.; Chen, M.X.; Ai, L.; Xu, X.N.; Jiao, J.M.; Zhu, T.J.; Su, H.Y.; Zang, W.; Luo, J.J.; Guo, Y.H.; et al. An outbreak of human fascioliasis gigantica in southwest china. PLoS One 2013, 8, e71520. [Google Scholar] [CrossRef] [PubMed]
- Cuervo, P.; Sidoti, L.; Fantozzi, C.; Neira, G.; Gerbeno, L.; Sierra, R.M. Fasciola hepatica infection and association with gastrointestinal parasites in creole goats from western argentina. Rev. Bras. Parasitol. Vet. 2013, 22, 53–57. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Pesticides and Their Application for the Control of Vectors and Pests of Public Health Importance. Available online: http://www.who.int/iris/handle/10665/69223#sthash.OVco5rEP.dpuf (accessed on 30 June 2013).
- Carballo, J.L.; Hernandez-Inda, Z.L.; Perez, P.; Garcia-Gravalos, M.D. A comparison between two brine shrimp assays to detect in vitro cytotoxicity in marine natural products. BMC Biotechnol. 2002, 2, 17. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Rittschof, D.; Orihuela, B.; Kwok, K.W.; Stafslien, S.; Chisholm, B. The effects of model polysiloxane and fouling-release coatings on embryonic development of a sea urchin (arbacia punctulata) and a fish (oryzias latipes). Aquat. Toxicol. 2012, 110–111, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Khosrovyan, A.; Rodriguez-Romero, A.; Salamanca, M.J.; Del Valls, T.A.; Riba, I.; Serrano, F. Comparative performances of eggs and embryos of sea urchin (paracentrotus lividus) in toxicity bioassays used for assessment of marine sediment quality. Mar. Pollut. Bull. 2013, 70, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Migliore, L.; Civitareale, C.; Brambilla, G.; Di Delupis, G.D. Toxicity of several important agricultural antibiotics to artemia. Water Res. 1997, 31, 1801–1806. [Google Scholar] [CrossRef]
- Chang, Z.; Flatt, P.; Gerwick, W.H.; Nguyen, V.A.; Willis, C.L.; Sherman, D.H. The barbamide biosynthetic gene cluster: A novel marine cyanobacterial system of mixed polyketide synthase (pks)-non-ribosomal peptide synthetase (nrps) origin involving an unusual trichloroleucyl starter unit. Gene 2002, 296, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Flatt, P.M.; O’Connell, S.J.; McPhail, K.L.; Zeller, G.; Willis, C.L.; Sherman, D.H.; Gerwick, W.H. Characterization of the initial enzymatic steps of barbamide biosynthesis. J. Nat. Prod. 2006, 69, 938–944. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Lee, J.H.; Choi, H.; Pereira, A.R.; Ban, Y.H.; Yoo, Y.J.; Kim, E.; Park, J.W.; Sherman, D.H.; Gerwick, W.H.; et al. Heterologous production of 4-O-demethylbarbamide, a marine cyanobacterial natural product. Org. Lett. 2012, 14, 5824–5827. [Google Scholar] [CrossRef] [PubMed]
- Ilardi, E.A.; Zakarian, A. Efficient total synthesis of dysidenin, dysidin, and barbamide. Chem. Asian J. 2011, 6, 2260–2263. [Google Scholar] [CrossRef] [PubMed]
- Krauss, J. Total synthesis of (±) tanikolide. Nat. Prod. Lett. 2001, 15, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Schomaker, J.M.; Borhan, B. Total synthesis of (+)-tanikolide via oxidative lactonization. Org. Biomol. Chem. 2004, 2, 621–624. [Google Scholar] [CrossRef] [PubMed]
- Arasaki, H.; Iwata, M.; Makida, M.; Masaki, Y. Synthesis of (R)-(+)-tanikolide through stereospecific C–H insertion reaction of dichlorocarbene with optically active secondary alcohol derivatives. Chem. Pharm. Bull. 2004, 52, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Kita, Y.; Matsuda, S.; Fujii, E.; Horai, M.; Hata, K.; Fujioka, H. Domino reaction of 2,3-epoxy-1-alcohols and pifa in the presence of h2o and the concise synthesis of (+)-tanikolide. Angew. Chem. 2005, 44, 5857–5860. [Google Scholar] [CrossRef]
- Fujioka, H.; Matsuda, S.; Horai, M.; Fujii, E.; Morishita, M.; Nishiguchi, N.; Hata, K.; Kita, Y. Facile and efficient synthesis of lactols by a domino reaction of 2,3-epoxy alcohols with a hypervalent iodine(iii) reagent and its application to the synthesis of lactones and the asymmetric synthesis of (+)-tanikolide. Chemistry 2007, 13, 5238–5248. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Hong, J. Total synthesis of cyanolide A and confirmation of its absolute configuration. Org. Lett. 2010, 12, 2880–2883. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Xie, X.; Jing, P.; Zhao, G.; Zheng, J.; Zhao, C.; She, X. Total synthesis of cyanolide A. Org. Biomol. Chem. 2011, 9, 984–986. [Google Scholar] [CrossRef] [PubMed]
- Hajare, A.K.; Ravikumar, V.; Khaleel, S.; Bhuniya, D.; Reddy, D.S. Synthesis of molluscicidal agent cyanolide A macrolactone from D-(−)-pantolactone. J. Org. Chem. 2011, 76, 963–966. [Google Scholar] [CrossRef] [PubMed]
- Pabbaraja, S.; Satyanarayana, K.; Ganganna, B.; Yadav, J.S. Formal total synthesis of cyanolide A. J. Org. Chem. 2011, 76, 1922–1925. [Google Scholar] [CrossRef] [PubMed]
- Gesinski, M.R.; Rychnovsky, S.D. Total synthesis of the cyanolide A aglycon. J. Am. Chem. Soc. 2011, 133, 9727–9729. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, R.J.; Jennings, M.P. A formal synthesis of (−)-cyanolide A featuring a stereoselective mukaiyama aldol reaction and oxocarbenium reduction. J. Org. Chem. 2011, 76, 8027–8032. [Google Scholar] [CrossRef] [PubMed]
- Waldeck, A.R.; Krische, M.J. Total synthesis of cyanolide A in the absence of protecting groups, chiral auxiliaries, or premetalated carbon nucleophiles. Angew. Chem. 2013, 52, 4470–4473. [Google Scholar] [CrossRef]
- Tay, G.C.; Gesinski, M.R.; Rychnovsky, S.D. Formation of highly substituted tetrahydropyranones: Application to the total synthesis of cyanolide A. Org. Lett. 2013, 15, 4536–4539. [Google Scholar] [PubMed]
- White, J.D.; Xu, Q.; Lee, C.S.; Valeriote, F.A. Total synthesis and biological evaluation of +-kalkitoxin, a cytotoxic metabolite of the cyanobacterium lyngbya majuscula. Org. Biomol. Chem. 2004, 2, 2092–2102. [Google Scholar] [CrossRef] [PubMed]
- LePage, K.T.; Goeger, D.; Yokokawa, F.; Asano, T.; Shioiri, T.; Gerwick, W.H.; Murray, T.F. The neurotoxic lipopeptide kalkitoxin interacts with voltage-sensitive sodium channels in cerebellar granule neurons. Toxicol. Lett. 2005, 158, 133–139. [Google Scholar] [CrossRef] [PubMed]
- White, J.D.; Lee, C.S.; Xu, Q. Total synthesis of (+)-kalkitoxin. Chem. Commun. 2003, 16, 2012–2013. [Google Scholar] [CrossRef]
- Umezawa, T.; Sueda, M.; Kamura, T.; Kawahara, T.; Han, X.; Okino, T.; Matsuda, F. Synthesis and biological activity of kalkitoxin and its analogues. J. Org. Chem. 2012, 77, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.D.; Gerwick, W.H. Curacins B and C, new antimitotic natural products from the marine cyanobacterium lyngbya majuscula. J. Nat. Prod. 1995, 58, 1961–1965. [Google Scholar] [CrossRef]
- Marquez, B.; Verdier-Pinard, P.; Hamel, E.; Gerwick, W.H. Curacin D, an antimitotic agent from the marine cyanobacterium lyngbya majuscula. Phytochemistry 1998, 49, 2387–2389. [Google Scholar] [CrossRef] [PubMed]
- Wipf, P.; Xu, W. Total synthesis of the antimitotic marine natural product (+)-curacin A. J. Org. Chem. 1996, 61, 6556–6562. [Google Scholar] [CrossRef] [PubMed]
- Verdier-Pinard, P.; Sitachitta, N.; Rossi, J.V.; Sackett, D.L.; Gerwick, W.H.; Hamel, E. Biosynthesis of radiolabeled curacin A and its rapid and apparently irreversible binding to the colchicine site of tubulin. Arch. Biochem. Biophys. 1999, 370, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Sitachitta, N.; Rossi, J.V.; Roberts, M.A.; Flatt, P.M.; Jia, J.; Sherman, D.H.; Gerwick, W.H. Biosynthetic pathway and gene cluster analysis of curacin A, an antitubulin natural product from the tropical marine cyanobacterium lyngbya majuscula. J. Nat. Prod. 2004, 67, 1356–1367. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Jia, J.; Liu, H.; Hakansson, K.; Gerwick, W.H.; Sherman, D.H. Metabolic coupling of dehydration and decarboxylation in the curacin A pathway: Functional identification of a mechanistically diverse enzyme pair. J. Am. Chem. Soc. 2006, 128, 9014–9015. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Wang, B.; Kulkarni, A.; Gehret, J.J.; Lloyd, K.R.; Gerwick, L.; Gerwick, W.H.; Wipf, P.; Hakansson, K.; Smith, J.L.; et al. Polyketide decarboxylative chain termination preceded by O-sulfonation in curacin A biosynthesis. J. Am. Chem. Soc. 2009, 131, 16033–16035. [Google Scholar] [CrossRef] [PubMed]
- Khare, D.; Wang, B.; Gu, L.; Razelun, J.; Sherman, D.H.; Gerwick, W.H.; Hakansson, K.; Smith, J.L. Conformational switch triggered by alpha-ketoglutarate in a halogenase of curacin A biosynthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 14099–14104. [Google Scholar] [CrossRef] [PubMed]
- Gehret, J.J.; Gu, L.; Gerwick, W.H.; Wipf, P.; Sherman, D.H.; Smith, J.L. Terminal alkene formation by the thioesterase of curacin A biosynthesis: Structure of a decarboxylating thioesterase. J. Biol. Chem. 2011, 286, 14445–14454. [Google Scholar] [CrossRef] [PubMed]
- Busche, A.; Gottstein, D.; Hein, C.; Ripin, N.; Pader, I.; Tufar, P.; Eisman, E.B.; Gu, L.; Walsh, C.T.; Sherman, D.H.; et al. Characterization of molecular interactions between acp and halogenase domains in the curacin A polyketide synthase. ACS Chem. Biol. 2012, 7, 378–386. [Google Scholar] [CrossRef]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Isolation and structure of the cytotoxin lyngbyabellin B and absolute configuration of lyngbyapeptin A from the marine cyanobacterium lyngbya majuscula. J. Nat. Prod. 2000, 63, 1437–1439. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Bearden, D.W.; Rein, K.S. Biosynthetic origin of the 3-amino-2,5,7,8-tetrahydroxy-10-methylundecanoic acid moiety and absolute configuration of pahayokolides A and B. J. Nat. Prod. 2011, 74, 1535–1538. [Google Scholar] [CrossRef] [PubMed]
- Williamson, R.T.; Singh, I.P.; Gerwick, W.H. Taveuniamides: New chlorinated toxins from a mixed assemblage of marine cyanobacteria. Tetrahedron 2004, 60, 7025–7033. [Google Scholar] [CrossRef]
- Tan, L.T.; Sitachitta, N.; Gerwick, W.H. The guineamides, novel cyclic depsipeptides from a papua new guinea collection of the marine cyanobacterium lyngbya majuscula. J. Nat. Prod. 2003, 66, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.T.; Goh, B.P.; Tripathi, A.; Lim, M.G.; Dickinson, G.H.; Lee, S.S.; Teo, S.L. Natural antifoulants from the marine cyanobacterium lyngbya majuscula. Biofouling 2010, 26, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.T.; More, S.V.; Lu, I.H.; Hsu, J.C.; Chen, T.J.; Jen, Y.C.; Lu, C.K.; Li, W.S. Isomalyngamide A, A-1 and their analogs suppress cancer cell migration in vitro. Eur. J. Med. Chem. 2011, 46, 3810–3819. [Google Scholar] [CrossRef] [PubMed]
- More, S.V.; Chang, T.T.; Chiao, Y.P.; Jao, S.C.; Lu, C.K.; Li, W.S. Glycosylation enhances the anti-migratory activities of isomalyngamide A analogs. Eur. J. Med. Chem. 2013, 64, 169–178. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Essack, M.; Alzubaidy, H.S.; Bajic, V.B.; Archer, J.A.C. Chemical Compounds Toxic to Invertebrates Isolated from Marine Cyanobacteria of Potential Relevance to the Agricultural Industry. Toxins 2014, 6, 3058-3076. https://doi.org/10.3390/toxins6113058
Essack M, Alzubaidy HS, Bajic VB, Archer JAC. Chemical Compounds Toxic to Invertebrates Isolated from Marine Cyanobacteria of Potential Relevance to the Agricultural Industry. Toxins. 2014; 6(11):3058-3076. https://doi.org/10.3390/toxins6113058
Chicago/Turabian StyleEssack, Magbubah, Hanin S. Alzubaidy, Vladimir B. Bajic, and John A. C. Archer. 2014. "Chemical Compounds Toxic to Invertebrates Isolated from Marine Cyanobacteria of Potential Relevance to the Agricultural Industry" Toxins 6, no. 11: 3058-3076. https://doi.org/10.3390/toxins6113058
APA StyleEssack, M., Alzubaidy, H. S., Bajic, V. B., & Archer, J. A. C. (2014). Chemical Compounds Toxic to Invertebrates Isolated from Marine Cyanobacteria of Potential Relevance to the Agricultural Industry. Toxins, 6(11), 3058-3076. https://doi.org/10.3390/toxins6113058