Aedes aegypti Mos20 Cells Internalizes Cry Toxins by Endocytosis, and Actin Has a Role in the Defense against Cry11Aa Toxin

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
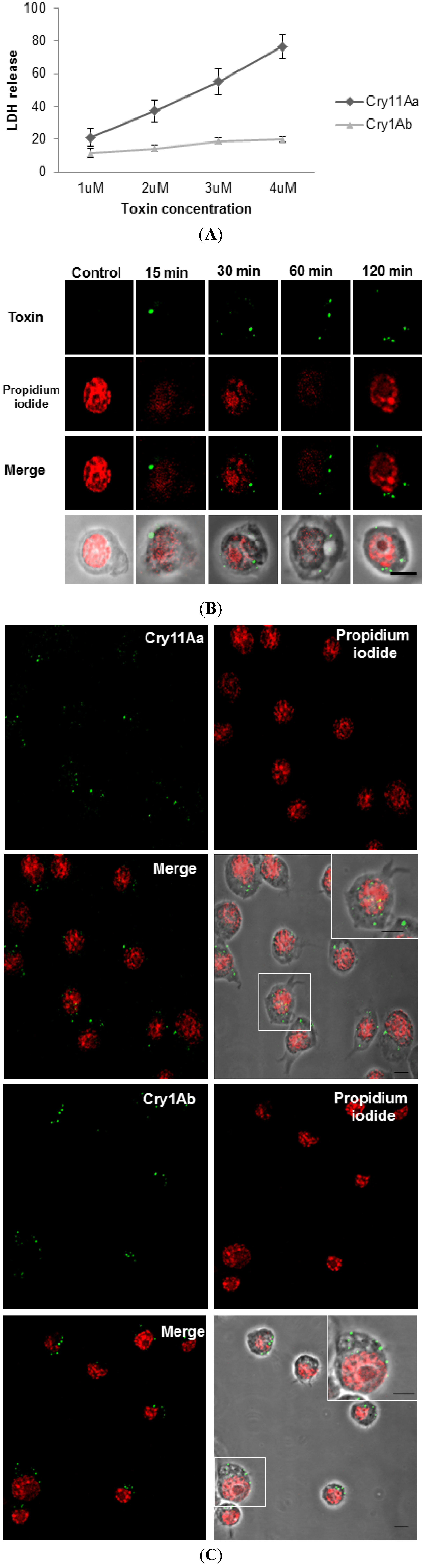
2.1. Both Cry11Aa and Cry1Ab Toxins Are Internalized into Mos20 Cells at Sub-Lethal Doses

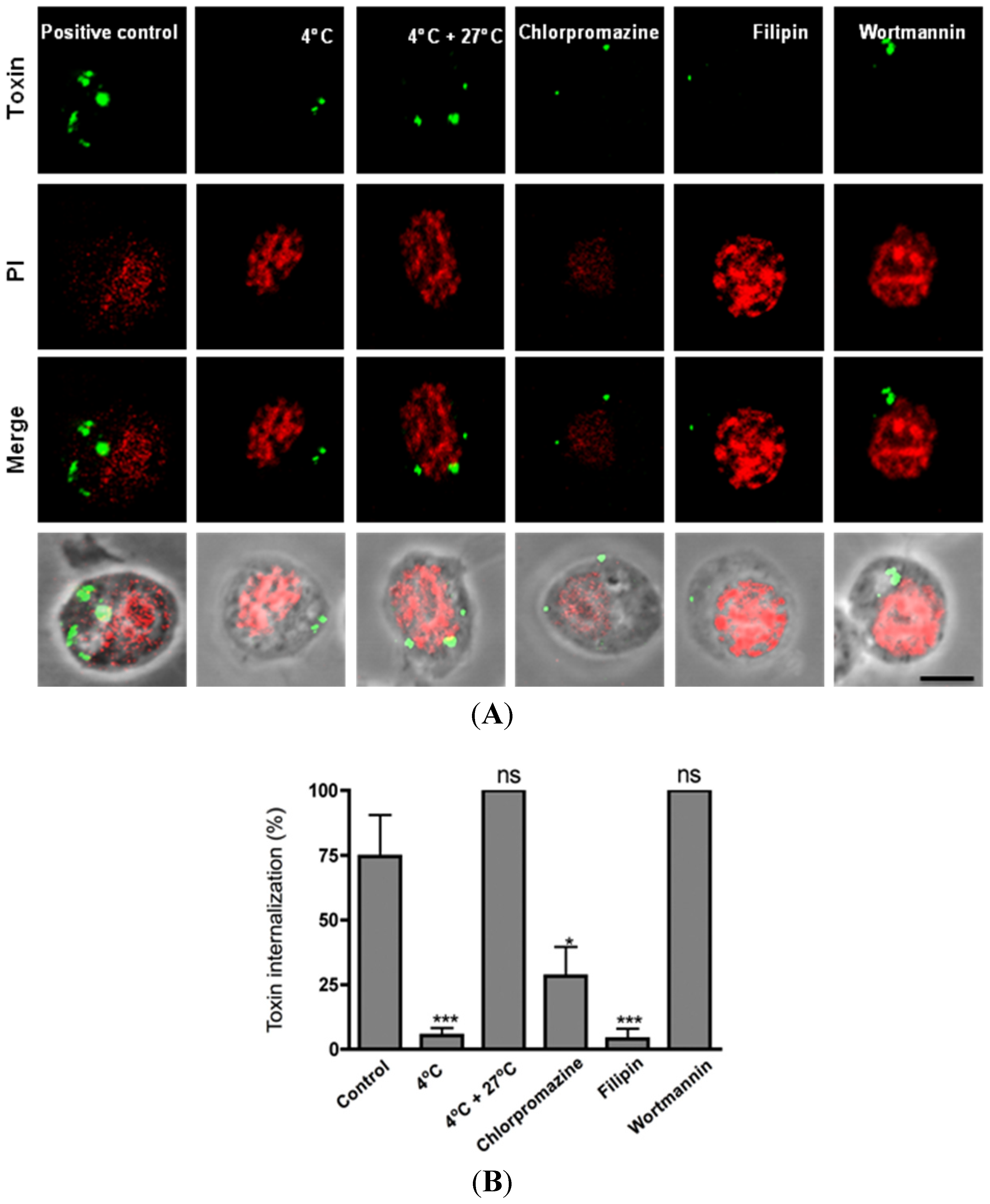
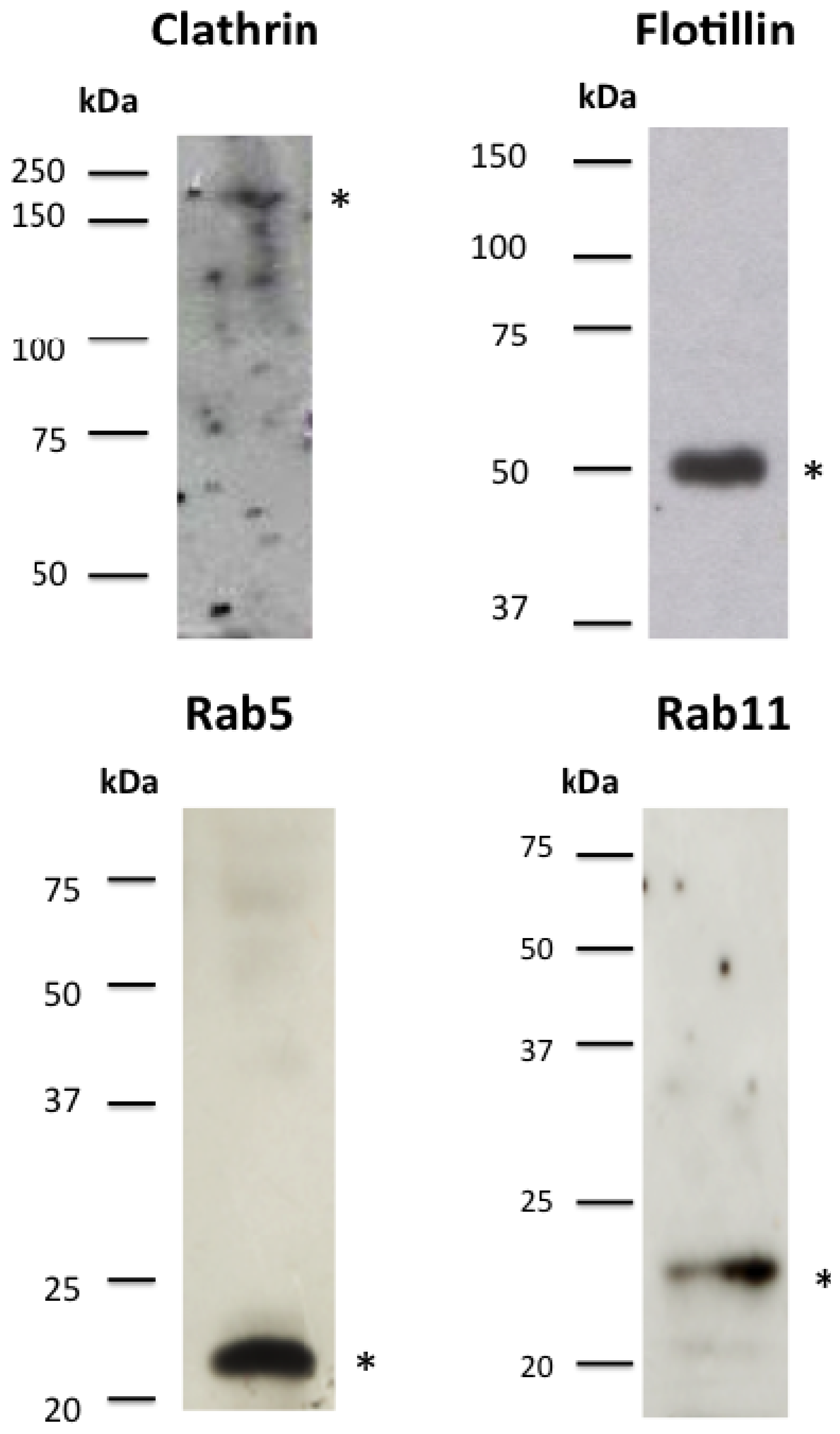
2.2. Clathrin-Dependent and Clathrin-Independent Mechanisms Are Involved in the Internalization of Cry11A

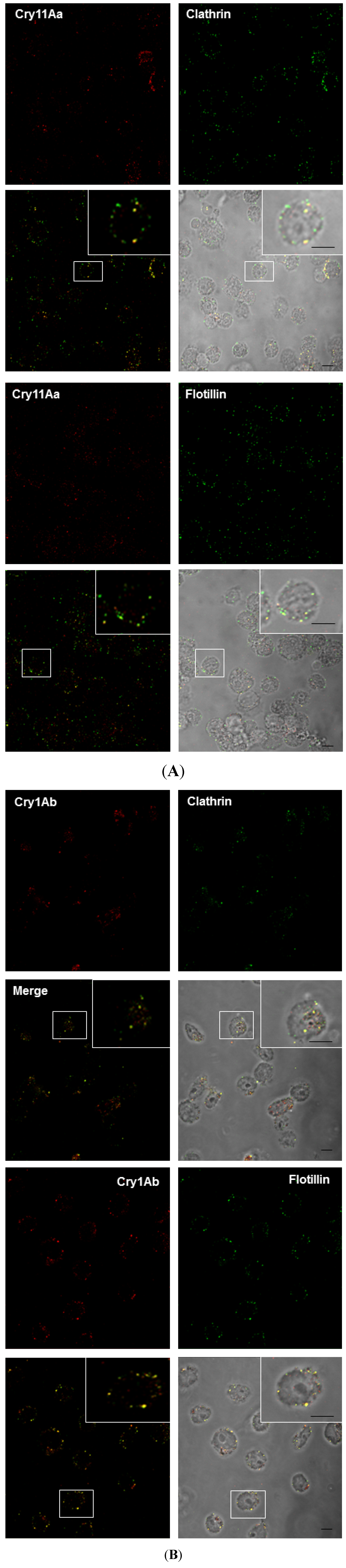
2.3. Cry Toxins Colocalize with Clathrin and Flotillin in the Mos20 Cell Line
2.4. Rab5 Colocalizes with Cry11Aa and Cry1Ab

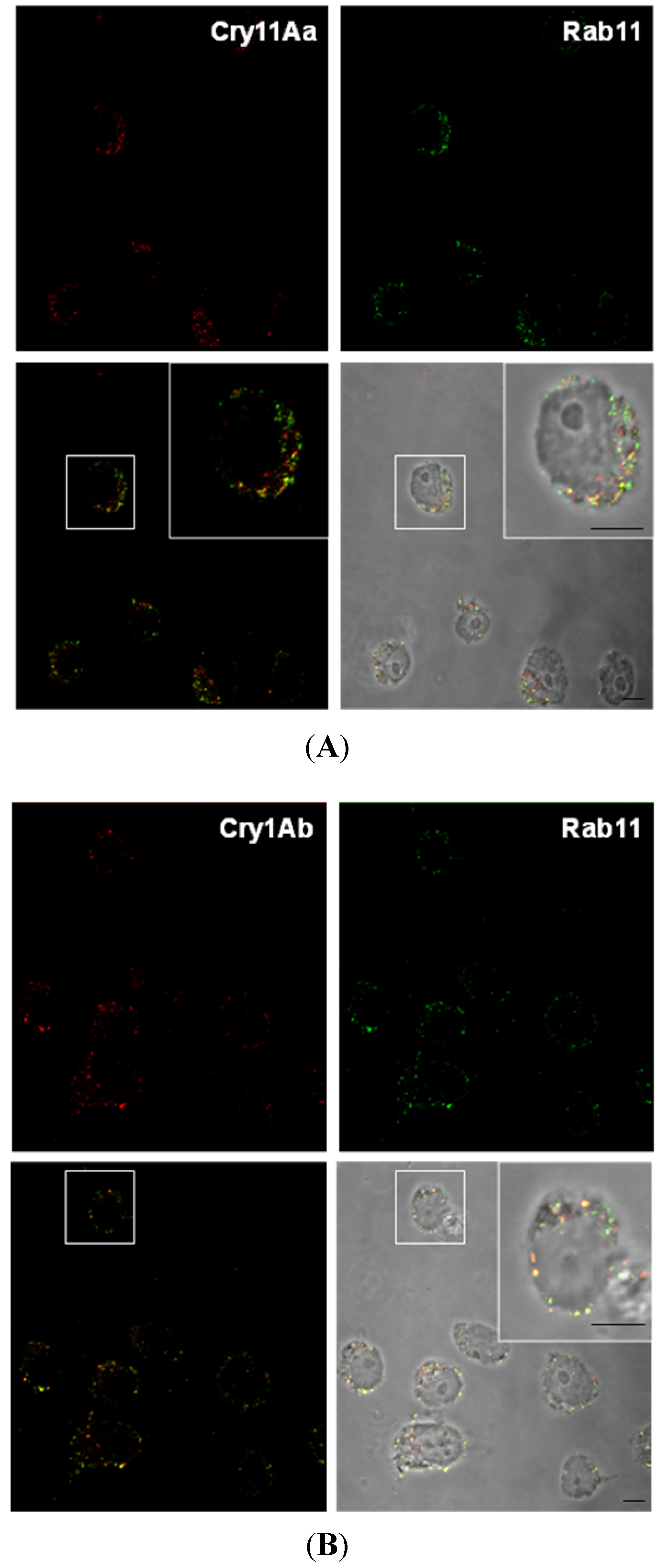
2.5. Rab11 Colocalizes with Cry11Aa and Cry1Ab


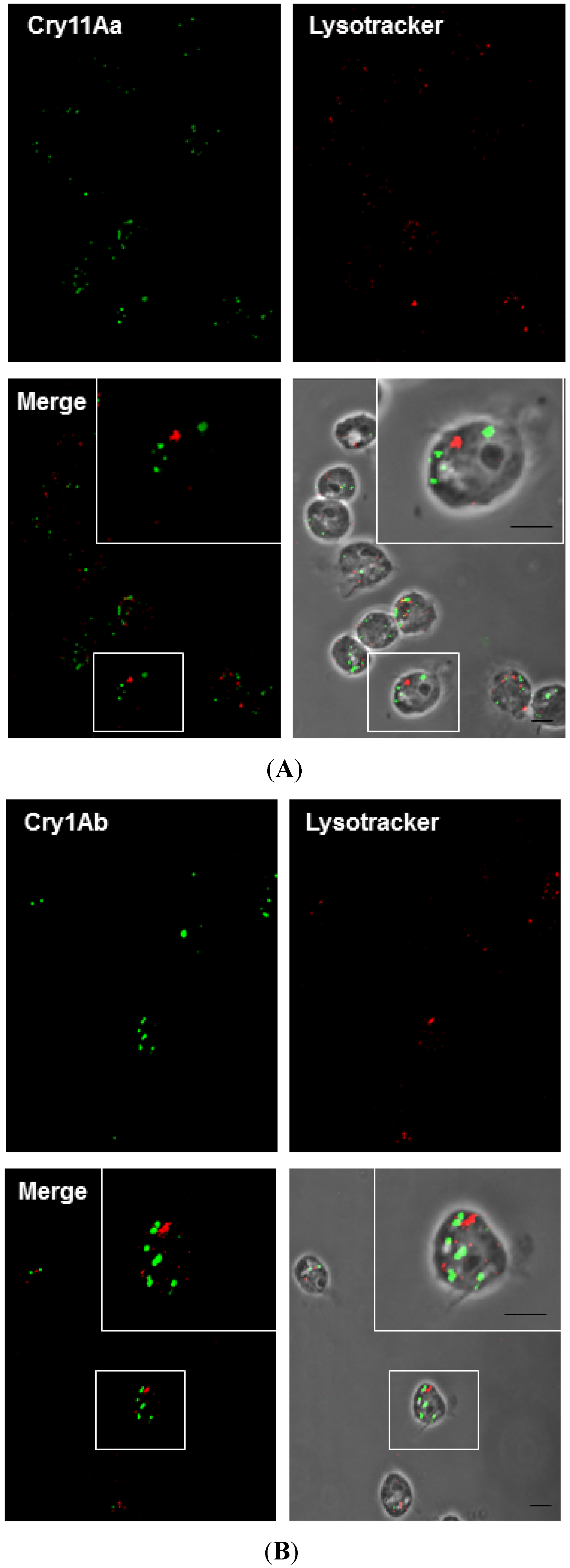
2.6. Cry11Aa and Cry1Ab Do Not Colocalize with Lysosomes

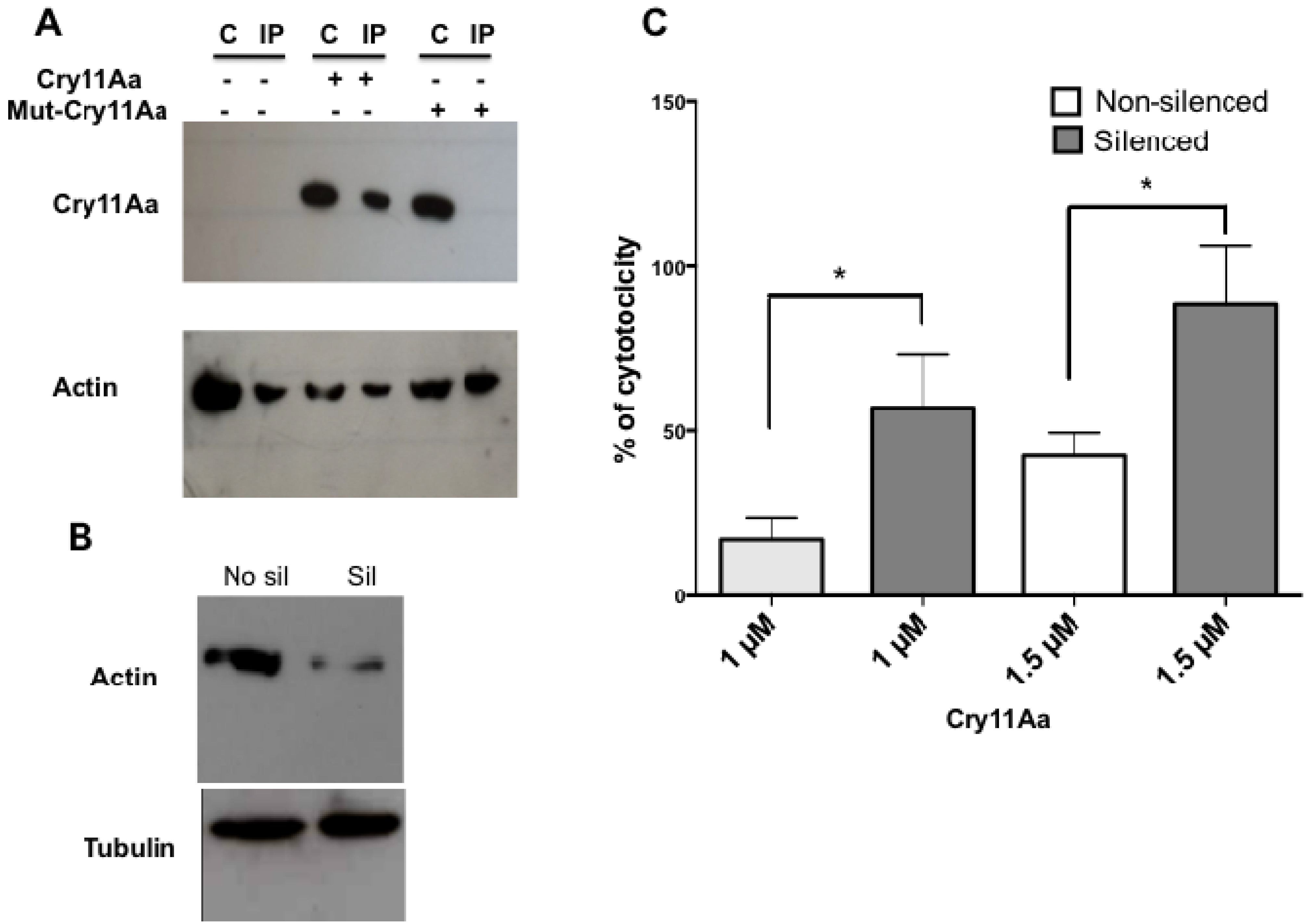
2.7. Actin Is Required for the Toxicity of Cry11Aa

Mos 20 Cells Are More Susceptible to Cry11Aa If Actin Was Silenced
3. Experimental Section
3.1. Cell Lines
3.2. Toxin Purification and Processing
3.3. Cell Viability Measurement
3.4. Inhibition Treatments
3.5. Antibodies
3.6. Immuno Fluorescence Assay
3.7. Western Blots Assay
3.8. Inhibition of Protein Expression by RNA Interference
3.9. Co-immunoprecipitation
3.10. Confocal Microscopy
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Jansen, C.C.; Beebe, N.W. The dengue vector Aedes aegypti: What comes next. Microbes Infect. 2010, 12, 272–279. [Google Scholar] [CrossRef]
- Aroian, R.; van der Goot, F.G. Pore-forming toxins and cellular non-immune defenses (CNIDs). Curr. Opin. Microbiol. 2007, 10, 57–61. [Google Scholar] [CrossRef]
- Conner, S.D.; Schröter, T.; Schmid, S.L. AAK1-mediated micro2 phosphorylation is stimulated by assembled clathrin. Traffic 2003, 4, 885–890. [Google Scholar]
- Husmann, M.; Beckmann, E.; Boller, K.; Kloft, N.; Tenzer, S.; Bobkiewicz, W.; Neukirch, C.; Bayley, H.; Bhakdi, S. Elimination of a bacterial pore-forming toxin by sequential endocytosis and exocytosis. FEBS Lett. 2009, 583, 337–344. [Google Scholar] [CrossRef]
- Idone, V.; Tam, C.; Goss, J.W.; Toomre, D.; Pypaert, M.; Andrews, N.W. Repair of injured plasma membrane by rapid Ca2+-dependent endocytosis. J. Cell Biol. 2008, 180, 905–914. [Google Scholar] [CrossRef]
- Keefe, D.; Shi, L.; Feske, S.; Massol, R.; Navarro, F.; Kirchhausen, T.; Lieberman, J. Perforin triggers a plasma membrane-repair response that facilitates CTL induction of apoptosis. Immunity 2005, 23, 249–262. [Google Scholar] [CrossRef]
- Griffitts, J.S.; Whitacre, J.L.; Stevens, D.E.; Aroian, R.V. Bt toxin resistance from loss of a putative carbohydrate-modifying enzyme. Science 2001, 293, 860–864. [Google Scholar] [CrossRef]
- Los, F.C.; Kao, C.Y.; Smitham, J.; McDonald, K.L.; Ha, C.; Peixoto, C.A.; Aroian, R.V. RAB-5- and RAB-11-dependent vesicle-trafficking pathways are required for plasma membrane repair after attack by bacterial pore-forming toxin. Cell Host Microbe 2011, 9, 147–157. [Google Scholar]
- Aktories, K. Bacterial protein toxins that modify host regulatory GTPases. Nature Rev. Microbiol. 2011, 9, 487–498. [Google Scholar]
- Gouin, E.; Egile, C.; Dehoux, P.; Villiers, V.; Adams, J.; Gertler, F.; Li, R.; Cossart, P. The RickA protein of Rickettsia conorii activates the Arp2/3 complex. Nature 2004, 427, 457–461. [Google Scholar] [CrossRef]
- Hupp, S.; Fortsch, C.; Wippel, C.; Ma, J.; Mitchell, T.J.; Iliev, A.I. Direct transmembrane interaction between actin and the pore-competent, cholesterol-dependent cytolysin pneumolysin. J. Mol. Biol. 2013, 425, 636–646. [Google Scholar] [CrossRef]
- Abrami, L.; Bischofberger, M.; Kunz, B.; Groux, R.; van der Goot, F.G. Endocytosis of the anthrax toxin is mediated by clathrin, actin and unconventional adaptors. PLoS Pathog. 2010, 6, e1000792. [Google Scholar] [CrossRef]
- McNall, R.J.; Adang, M.J. Identification of novel Bacillus thuringiensis Cry1Ac binding proteins in Manduca sexta midgut through proteomic analysis. Insect Biochem. Mol. Biol. 2003, 33, 999–1010. [Google Scholar] [CrossRef]
- Bayyareddy, K.; Andacht, T.M.; Abdullah, M.A.; Adang, M.J. Proteomic identification of Bacillus thuringiensis subsp. israelensis toxin Cry4Ba binding proteins in midgut membranes from Aedes (Stegomyia) aegypti Linnaeus (Diptera, Culicidae) larvae. Insect Biochem. Mol. Biol. 2009, 39, 279–286. [Google Scholar] [CrossRef]
- Yuan, C.; Ding, X.; Xia, L.; Yin, J.; Huang, S.; Huang, F. Proteomic analysis of BBMV in Helicoverpa armigera midgut with and without Cry1Ac toxin treatment. Biocontrol Sci. Technol. 2011, 21, 139–151. [Google Scholar] [CrossRef]
- Cancino-Rodezno, A.; Lozano, L.; Oppert, C.; Castro, J.I.; Lanz-Mendoza, H.; Encarnacion, S.; Evans, A.E.; Gill, S.S.; Soberon, M.; Jurat-Fuentes, J.L.; et al. Comparative proteomic analysis of Aedes aegypti larval midgut after intoxication with Cry11Aa toxin from Bacillus thuringiensis. PloS ONE 2012, 7, e37034. [Google Scholar] [CrossRef]
- Varma, M.G.; Pudney, M. The growth and serial passage of cell lines from Aedes aegypti (L.) larvae in different media. J. Med. Entomol. 1996, 6, 432–439. [Google Scholar]
- Kuismanen, E.; Saraste, J. Low temperature-induced transport blocks as tools to manipulate membrane traffic. Meth. Cell Biol. 1998, 32, 257–274. [Google Scholar] [CrossRef]
- Wang, L.H.; Rothberg, K.G.; Anderson, R.G. Mis-assembly of clathrin lattices on endosomes reveals a regulatory switch for coated pit formation. J. Cell Biol. 1993, 123, 1107–1117. [Google Scholar] [CrossRef]
- Orlandi, P.A.; Fishman, P.H. Filipin-dependent inhibition of cholera toxin: evidence for toxin internalization and activation through caveolae-like domains. J. Cell Biol. 1998, 141, 905–915. [Google Scholar] [CrossRef]
- Glebov, O.O.; Bright, N.A.; Nichols, B.J. Flotillin-1 defines a clathrin-independent endocytic pathway in mammalian cells. Nat. Cell Biol. 2006, 8, 46–54. [Google Scholar] [CrossRef]
- ImageJ. Available online: http://rsb.info.nih.gov/ij/ (accessed on 09 December 2009).
- Fisher, H.M.; Brewis, I.A.; Barratt, C.L.; Cooke, I.D.; Moore, H.D. Phosphoinositide 3-kinase is involved in the induction of the human sperm acrosome reaction downstream of tyrosine phosphorylation. Mol. Hum. Reprod. 1998, 4, 849–855. [Google Scholar] [CrossRef]
- Hansen, S.H.; Olsson, A.; Casanova, J.E. Wortmannin, an inhibitor of phosphoinositide 3-kinase, inhibits transcytosis in polarized epithelial cells. J. Biol. Chem. 1995, 270, 28425–28432. [Google Scholar] [CrossRef]
- Bayyareddy, K.; Zhu, X.; Orlando, R.; Adang, M.J. Proteome analysis of Cry4Ba toxin-interacting Aedes aegypti lipid rafts using geLC-MS/MS. J. Proteome Research 2012, 11, 5843–5855. [Google Scholar]
- Le Roy, C.; Wrana, J.L. Clathrin- and non-clathrin-mediated endocytic regulation of cell signalling. Nat. Rev. Mol. Cell Biol. 2005, 6, 112–126. [Google Scholar]
- Maxfield, F.R.; McGraw, T.E. Endocytic recycling. Nat. Rev. Mol. Cell Biol. 2004, 5, 121–132. [Google Scholar] [CrossRef]
- Olkkonen, V.M.; Stenmark, H. Role of Rab GTPases in membrane traffic. Int. Rev. Cytol. 1997, 176, 1–85. [Google Scholar]
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef]
- Grant, B.D.; Donaldson, J.G. Pathways and mechanisms of endocytic recycling. Nat. Rev. Mol. Cell Biol. 2009, 10, 597–608. [Google Scholar] [CrossRef]
- McNeil, P.L.; Kirchhausen, T. An emergency response team for membrane repair. Nat. Rev. Mol. Cell Biol. 2005, 6, 499–505. [Google Scholar] [CrossRef]
- Baxt, L.A.; Garza-Mayers, A.C.; Goldberg, M.B. Bacterial subversion of host innate immune pathways. Science 2013, 340, 697–701. [Google Scholar] [CrossRef]
- Luzio, J.P.; Pryor, P.R.; Bright, N.A. Lysosomes: Fusion and function. Nat. Rev. Mol. Cell Biol. 2007, 8, 622–632. [Google Scholar] [CrossRef]
- Fujimoto, L.M.; Roth, R.; Heuser, J.E.; Schmid, S.L. Actin assembly plays a variable, but not obligatory role in receptor-mediated endocytosis in mammalian cells. Traffic 2000, 1, 161–171. [Google Scholar] [CrossRef]
- Saffarian, S.; Cocucci, E.; Kirchhausen, T. Distinct dynamics of endocytic clathrin-coated pits and coated plaques. PLoS Biol. 2009, 7, e1000191. [Google Scholar] [CrossRef]
- Carmona, D.; Rodriguez-Almazan, C.; Muñoz-Garay, C.; Portugal, L.; Perez, C.; de Maagd, R.A.; Bakker, P.; Soberon, M.; Bravo, A. Dominant negative phenotype of Bacillus thuringiensis Cry1Ab, Cry11Aa and Cry4Ba mutants suggest hetero-oligomer formation among different Cry toxins. PloS ONE 2011, 6, e19952. [Google Scholar] [CrossRef]
- Vachon, V.; Prefontaine, G.; Rang, C.; Coux, F.; Juteau, M.; Schwartz, J.L.; Brousseau, R.; Frutos, R.; Laprade, R.; Masson, L. Helix 4 mutants of the Bacillus thuringiensis insecticidal toxin Cry1Aa display altered pore-forming abilities. Appl. Environ. Microbiol. 2004, 70, 6123–6130. [Google Scholar] [CrossRef]
- Gazit, E.; La Rocca, P.; Sansom, M.S.; Shai, Y. The structure and organization within the membrane of the helices composing the pore-forming domain of Bacillus thuringiensis delta-endotoxin are consistent with an “umbrella-like” structure of the pore. Proc. Natl. Acad. Sci USA 1998, 95, 12289–12294. [Google Scholar] [CrossRef]
- Taveecharoenkool, T.; Angsuthanasombat, C.; Kanchanawarin, C. Combined molecular dynamics and continuum solvent studies of the pre-pore Cry4Aa trimer suggest its stability in solution and how it may form pore. PMC Biophys. 2010, 3. [Google Scholar] [CrossRef]
- Zavala, L.E.; Pardo-Lopez, L.; Canton, P.E.; Gomez, I.; Soberon, M.; Bravo, A. Domains II and III of Bacillus thuringiensis Cry1Ab toxin remain exposed to the solvent after insertion of part of domain I into the membrane. J. Biol. Chem. 2011, 286, 19109–19117. [Google Scholar]
- Chen, J.; Aimanova, K.G.; Fernandez, L.E.; Bravo, A.; Soberon, M.; Gill, S.S. Aedes aegypti cadherin serves as a putative receptor of the Cry11Aa toxin from Bacillus thuringiensis subsp. israelensis. Biochem. J. 2009, 424, 191–200. [Google Scholar] [CrossRef]
- Chang, C.; Yu, Y.M.; Dai, S.M.; Law, S.K.; Gill, S.S. High-level cryIVD and cytA gene expression in Bacillus thuringiensis does not require the 20-kilodalton protein, and the coexpressed gene products are synergistic in their toxicity to mosquitoes. Appl. Environ. Microbiol. 1993, 59, 815–821. [Google Scholar]
- Wu, D.; Johnson, J.J.; Federici, B.A. Synergism of mosquitocidal toxicity between CytA and CryIVD proteins using inclusions produced from cloned genes of Bacillus thuringiensis. Mol. Microbiol. 1994, 13, 965–972. [Google Scholar] [CrossRef]
- Thomas, W.E.; Ellar, D.J. Mechanism of action of Bacillus thuringiensis var israelensis insecticidal delta-endotoxin. FEBS Lett. 1983, 154, 362–368. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Cancino-Rodezno, A.; Alexander, C.; Villasenor, R.; Pacheco, S.; Porta, H.; Pauchet, Y.; Soberon, M.; Gill, S.S.; Bravo, A. The mitogen-activated protein kinase p38 is involved in insect defense against Cry toxins from Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2010, 40, 58–63. [Google Scholar] [CrossRef]
Supplementary Files

© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Vega-Cabrera, A.; Cancino-Rodezno, A.; Porta, H.; Pardo-Lopez, L. Aedes aegypti Mos20 Cells Internalizes Cry Toxins by Endocytosis, and Actin Has a Role in the Defense against Cry11Aa Toxin. Toxins 2014, 6, 464-487. https://doi.org/10.3390/toxins6020464
Vega-Cabrera A, Cancino-Rodezno A, Porta H, Pardo-Lopez L. Aedes aegypti Mos20 Cells Internalizes Cry Toxins by Endocytosis, and Actin Has a Role in the Defense against Cry11Aa Toxin. Toxins. 2014; 6(2):464-487. https://doi.org/10.3390/toxins6020464
Chicago/Turabian StyleVega-Cabrera, Adriana, Angeles Cancino-Rodezno, Helena Porta, and Liliana Pardo-Lopez. 2014. "Aedes aegypti Mos20 Cells Internalizes Cry Toxins by Endocytosis, and Actin Has a Role in the Defense against Cry11Aa Toxin" Toxins 6, no. 2: 464-487. https://doi.org/10.3390/toxins6020464
APA StyleVega-Cabrera, A., Cancino-Rodezno, A., Porta, H., & Pardo-Lopez, L. (2014). Aedes aegypti Mos20 Cells Internalizes Cry Toxins by Endocytosis, and Actin Has a Role in the Defense against Cry11Aa Toxin. Toxins, 6(2), 464-487. https://doi.org/10.3390/toxins6020464