Identification of a Key Residue for Oligomerisation and Pore-Formation of Clostridium perfringens NetB

Abstract
:1. Introduction
2. Results and Discussion
2.1. Purification of Wild Type NetB and Variants of NetB


2.2. Binding and Toxicity of NetB to LMH Cells and hRBCs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | α-helix (%) | β-sheet (%) | other (%) |
|---|---|---|---|
| NetB wt | 9.5 | 32.2 | 58.3 |
| NetB K41H | 7.2 | 33.2 | 59.6 |
| NetB K71A | 7.2 | 33.2 | 59.6 |
| NetB P155A | 8.6 | 32.5 | 58.9 |
| NetB D156C | 8.7 | 32.6 | 58.7 |
| NetB D250A | 9.5 | 32.2 | 58.3 |



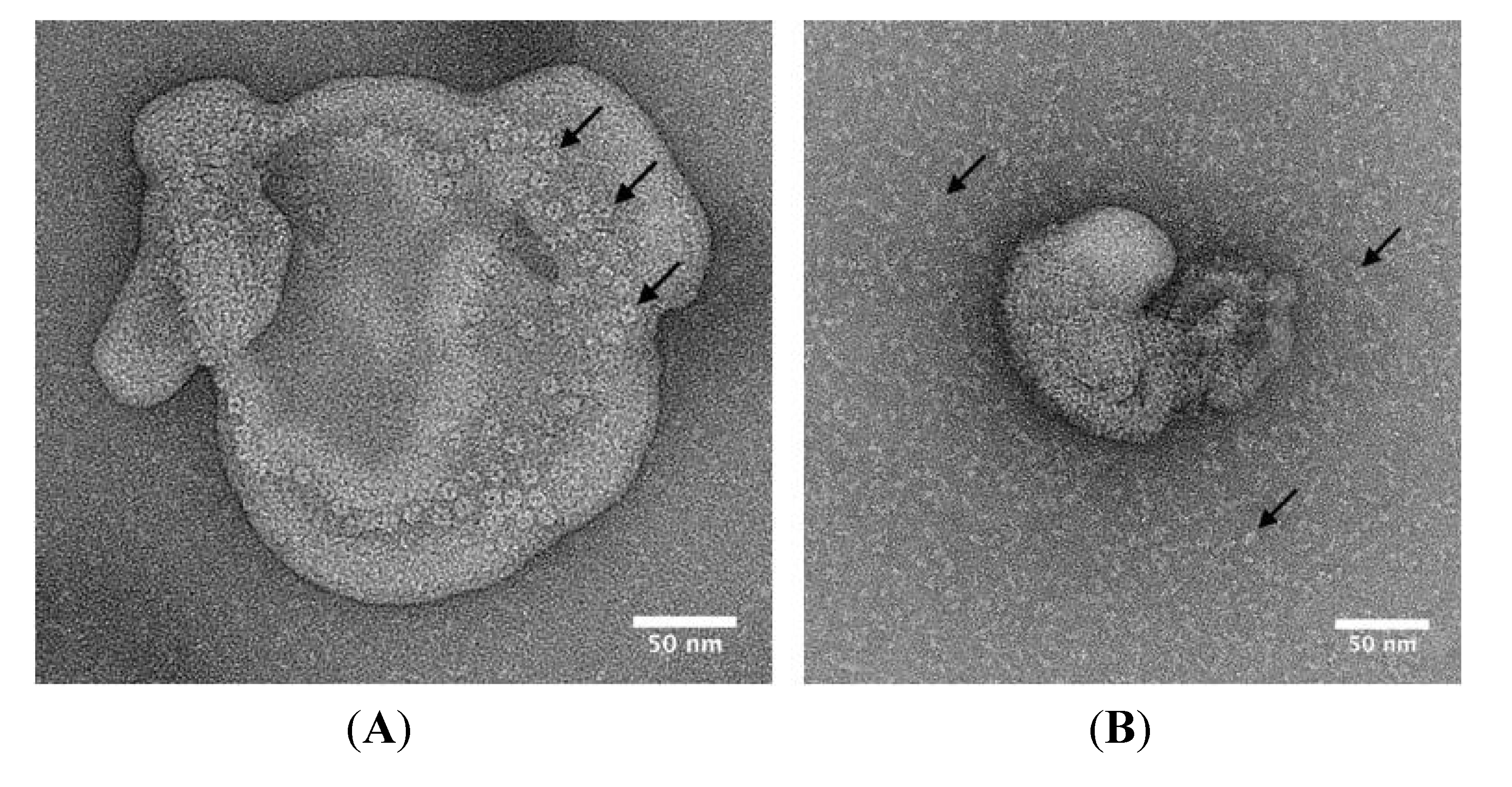
2.3. Oligomerisation and Pore-Formation of NetB



3. Discussions
4. Experimental Section
4.1. Expression and Purification of Wild Type NetB
4.2. Design and Purification of Variant Forms of NetB
4.3. CD Analyses
4.4. SDS-PAGE Analyses
4.5. On-Cell Western™ Assay
4.6. Cytotoxic Effect of NetB on LMH Cells
4.7. Hemolysis Assay
4.8. Osmotic Protection Assay
4.9. Binding and Oligomerisation of NetB to Liposomes
4.10. Electron Microscopy
4.11. Calcein Release Assay
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Keyburn, A.L.; Boyce, J.D.; Vaz, P.; Bannam, T.L.; Ford, M.E.; Parker, D.; di Rubbo, A.; Rood, J.I.; Moore, R.J. NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens. PLoS Pathog. 2008, 4, e26. [Google Scholar] [CrossRef]
- Yan, X.X.; Porter, C.J.; Hardy, S.P.; Steer, D.; Smith, A.I.; Quinsey, N.S.; Hughes, V.; Cheung, J.K.; Keyburn, A.L.; Kaldhusdal, M.; et al. Structural and functional analysis of the pore-forming toxin Netb from Clostridium perfringens. MBio 2013, 4. [Google Scholar] [CrossRef]
- Sluis, V.D. Clostridial enteritis is an often underestimated problem. World Poult. 2000, 16, 42–43. [Google Scholar]
- Savva, C.G.; Fernandes da Costa, S.P.; Bokori-Brown, M.; Naylor, C.E.; Cole, A.R.; Moss, D.S.; Titball, R.W.; Basak, A.K. Molecular architecture and functional analysis of NetB, a pore-forming toxin from Clostridium perfringens. J. Biol. Chem. 2013, 288, 3512–3522. [Google Scholar] [CrossRef]
- Song, L.; Hobaugh, M.R.; Shustak, C.; Cheley, S.; Bayley, H.; Gouaux, J.E. Structure of staphylococcal alpha-hemolysin, a heptameric transmembrane pore. Science 1996, 274, 1859–1866. [Google Scholar] [CrossRef]
- Yamashita, K.; Kawai, Y.; Tanaka, Y.; Hirano, N.; Kaneko, J.; Tomita, N.; Ohta, M.; Kamio, Y.; Yao, M.; Tanaka, I. Crystal structure of the octameric pore of staphylococcal gamma-hemolysin reveals the beta-barrel pore formation mechanism by two components. Proc. Natl. Acad. Sci. USA 2011, 108, 17314–17319. [Google Scholar] [CrossRef]
- Olson, R.; Gouaux, E. Crystal structure of the Vibrio cholerae cytolysin (VCC) pro-toxin and its assembly into a heptameric transmembrane pore. J. Mol. Biol. 2005, 350, 997–1016. [Google Scholar] [CrossRef]
- Valeva, A.; Palmer, M.; Bhakdi, S. Staphylococcal alpha-toxin: Formation of the heptameric pore is partially cooperative and proceeds through multiple intermediate stages. Biochemistry 1997, 36, 13298–13304. [Google Scholar] [CrossRef]
- Gouaux, E. Alpha-hemolysin from Staphylococcus aureus: An archetype of beta-barrel, channel-forming toxins. J. Struct. Biol. 1998, 121, 110–122. [Google Scholar] [CrossRef]
- Gouaux, E.; Hobaugh, M.; Song, L. Alpha-hemolysin, gamma-hemolysin, and leukocidin from Staphylococcus aureus: Distant in sequence but similar in structure. Protein Sci. 1997, 6, 2631–2635. [Google Scholar] [CrossRef]
- Kawate, T.; Gouaux, E. Arresting and releasing staphylococcal alpha-hemolysin at intermediate stages of pore formation by engineered disulfide bonds. Protein Sci. 2003, 12, 997–1006. [Google Scholar] [CrossRef]
- Walker, B.; Bayley, H. Key residues for membrane-binding, oligomerization, and pore-forming activity of staphylococcal alpha-hemolysin identified by cysteine scanning mutagenesis and targeted chemical modification. J. Biol. Chem. 1995, 270, 23065–23071. [Google Scholar] [CrossRef]
- Fang, Y.; Cheley, S.; Bayley, H.; Yang, J. The heptameric prepore of a staphylococcal alpha-hemolysin mutant in lipid bilayers imaged by atomic force microscopy. Biochemistry 1997, 36, 9518–9522. [Google Scholar] [CrossRef]
- Walker, B.; Braha, O.; Cheley, S.; Bayley, H. An intermediate in the assembly of a pore-forming protein trapped with a genetically-engineered switch. Chem. Biol. 1995, 2, 99–105. [Google Scholar]
- Steinthorsdottir, V.; Fridriksdottir, V.; Gunnarsson, E.; Andresson, O.S. Site-directed mutagenesis of Clostridium perfringens beta-toxin: Expression of wild-type and mutant toxins in Bacillus subtilis. FEMS Microbiol. Lett. 1998, 158, 17–23. [Google Scholar] [CrossRef]
- Meunier, O.; Ferreras, M.; Supersac, G.; Hoeper, F.; BabaMoussa, L.; Monteil, H.; Colin, D.A.; Menestrina, G.; Prevost, G. A predicted beta-sheet from class S components of staphylococcal gamma-hemolysin is essential for the secondary interaction of the class F component. Biochim. Biophys. Acta-Biomembr. 1997, 1326, 275–286. [Google Scholar] [CrossRef]
- Jursch, R.; Hildebrand, A.; Hobom, G.; Tranum-Jensen, J.; Ward, R.; Kehoe, M.; Bhakdi, S. Histidine residues near the N terminus of staphylococcal alpha-toxin as reporters of regions that are critical for oligomerization and pore formation. Infect. Immun. 1994, 62, 2249–2256. [Google Scholar]
- Menzies, B.E.; Kernodle, D.S. Site-directed mutagenesis of the alpha-toxin gene of Staphylococcus-aureus—Role of histidines in toxin activity in-vitro and in a murine model. Infect. Immun. 1994, 62, 1843–1847. [Google Scholar]
- Valeva, A.; Palmer, M.; Hilgert, K.; Kehoe, M.; Bhakdi, S. Correct oligomerization is a prerequisite for insertion of the central molecular domain of staphylococcal alpha-toxin into the lipid bilayer. Biochim. Biophys. Acta Biomembr. 1995, 1236, 213–218. [Google Scholar]
- Fernandes da Costa, S.P.; Mot, D.; Bokori-Brown, M.; Savva, C.G.; Basak, A.K.; van Immerseel, F.; Titball, R.W. Protection against avian necrotic enteritis after immunisation with NetB genetic or formaldehyde toxoids. Vaccine 2013, 31, 4003–4008. [Google Scholar] [CrossRef]
- Menzies, B.E.; Kernodle, D.S. Passive immunization with antiserum to a nontoxic alpha-toxin mutant from Staphylococcus aureus is protective in a murine model. Infect. Immun. 1996, 64, 1839–1841. [Google Scholar]
- Ragle, B.E.; Wardenburg, J.B. Anti-alpha-hemolysin monoclonal antibodies mediate protection against Staphylococcus aureus pneumonia. Infect. Immun. 2009, 77, 2712–2718. [Google Scholar] [CrossRef]
- Wardenburg, J.B.; Schneewind, O. Vaccine protection against Staphylococcus aureus pneumonia. J. Exp. Med. 2008, 205, 287–294. [Google Scholar] [CrossRef]
- Savitzky, A.; Golay, M.J.E. Smoothing and differentiation of data by simplified least squares procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fernandes da Costa, S.P.; Savva, C.G.; Bokori-Brown, M.; Naylor, C.E.; Moss, D.S.; Basak, A.K.; Titball, R.W. Identification of a Key Residue for Oligomerisation and Pore-Formation of Clostridium perfringens NetB. Toxins 2014, 6, 1049-1061. https://doi.org/10.3390/toxins6031049
Fernandes da Costa SP, Savva CG, Bokori-Brown M, Naylor CE, Moss DS, Basak AK, Titball RW. Identification of a Key Residue for Oligomerisation and Pore-Formation of Clostridium perfringens NetB. Toxins. 2014; 6(3):1049-1061. https://doi.org/10.3390/toxins6031049
Chicago/Turabian StyleFernandes da Costa, Sérgio P., Christos G. Savva, Monika Bokori-Brown, Claire E. Naylor, David S. Moss, Ajit K. Basak, and Richard W. Titball. 2014. "Identification of a Key Residue for Oligomerisation and Pore-Formation of Clostridium perfringens NetB" Toxins 6, no. 3: 1049-1061. https://doi.org/10.3390/toxins6031049
APA StyleFernandes da Costa, S. P., Savva, C. G., Bokori-Brown, M., Naylor, C. E., Moss, D. S., Basak, A. K., & Titball, R. W. (2014). Identification of a Key Residue for Oligomerisation and Pore-Formation of Clostridium perfringens NetB. Toxins, 6(3), 1049-1061. https://doi.org/10.3390/toxins6031049