Electrophysiological Characterization of Ts6 and Ts7, K+ Channel Toxins Isolated through an Improved Tityus serrulatus Venom Purification Procedure

 ,
, 

Abstract
:1. Introduction
2. Results
2.1. Purification and Biochemical Characterization of Ts6 and Ts7 Toxins



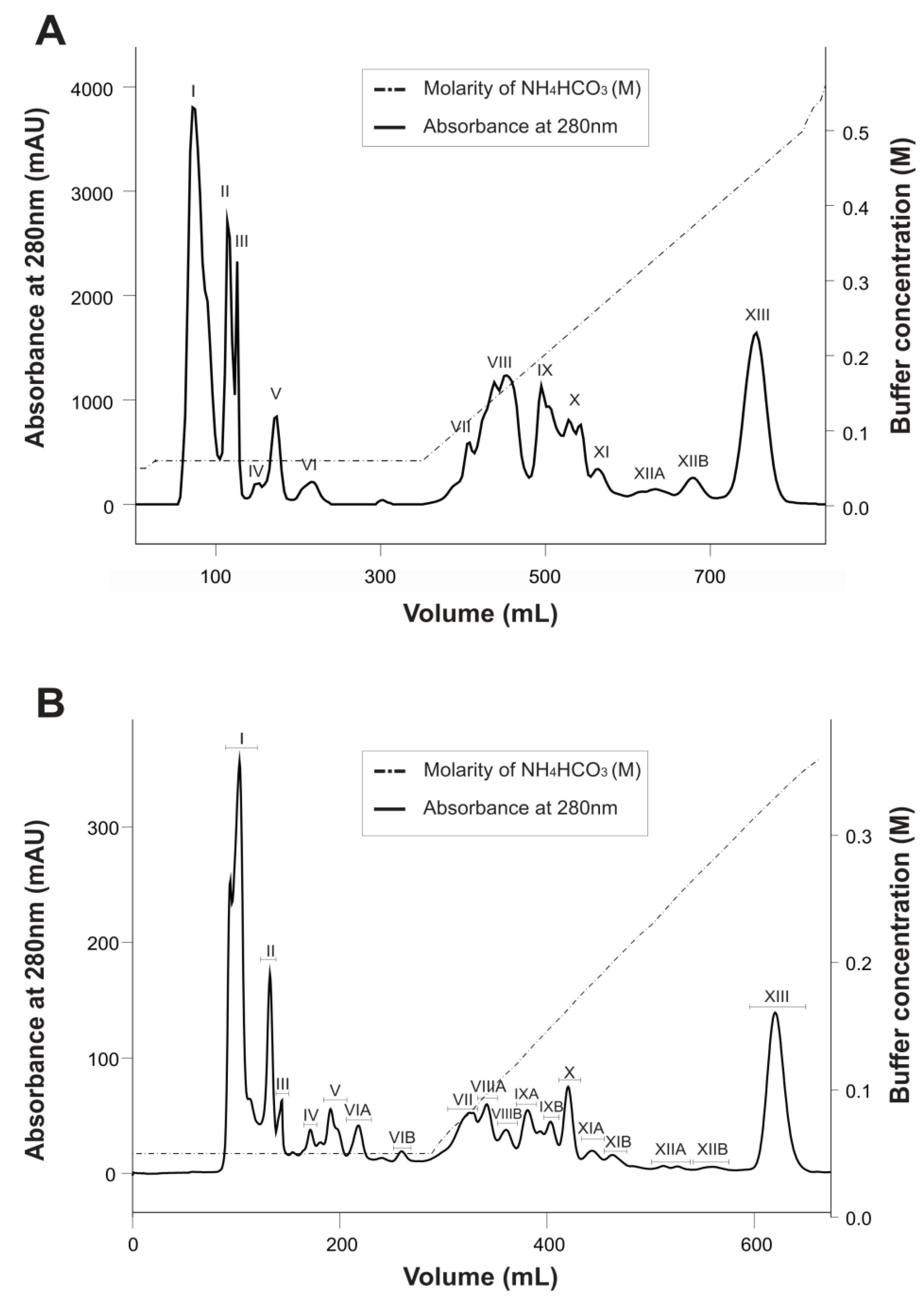{kind=link}
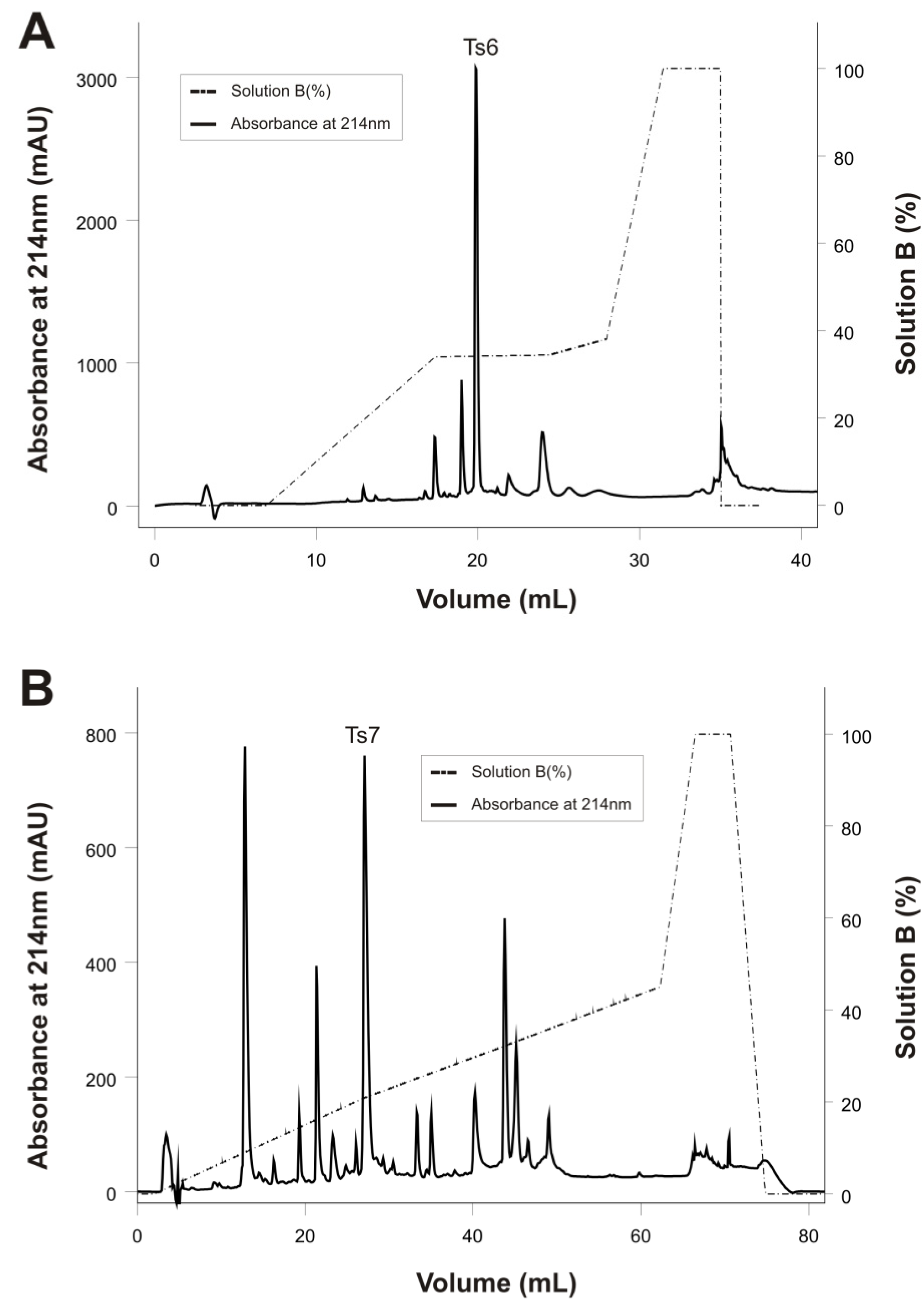{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction/Toxin | Column | Recovery % |
|---|---|---|
| X | CMC52 | 4.62 |
| Ts6 | C18 | 1.82 |
| XIIA | CMC52 | 0.95 |
| Ts7 | C18 | 0.24 |
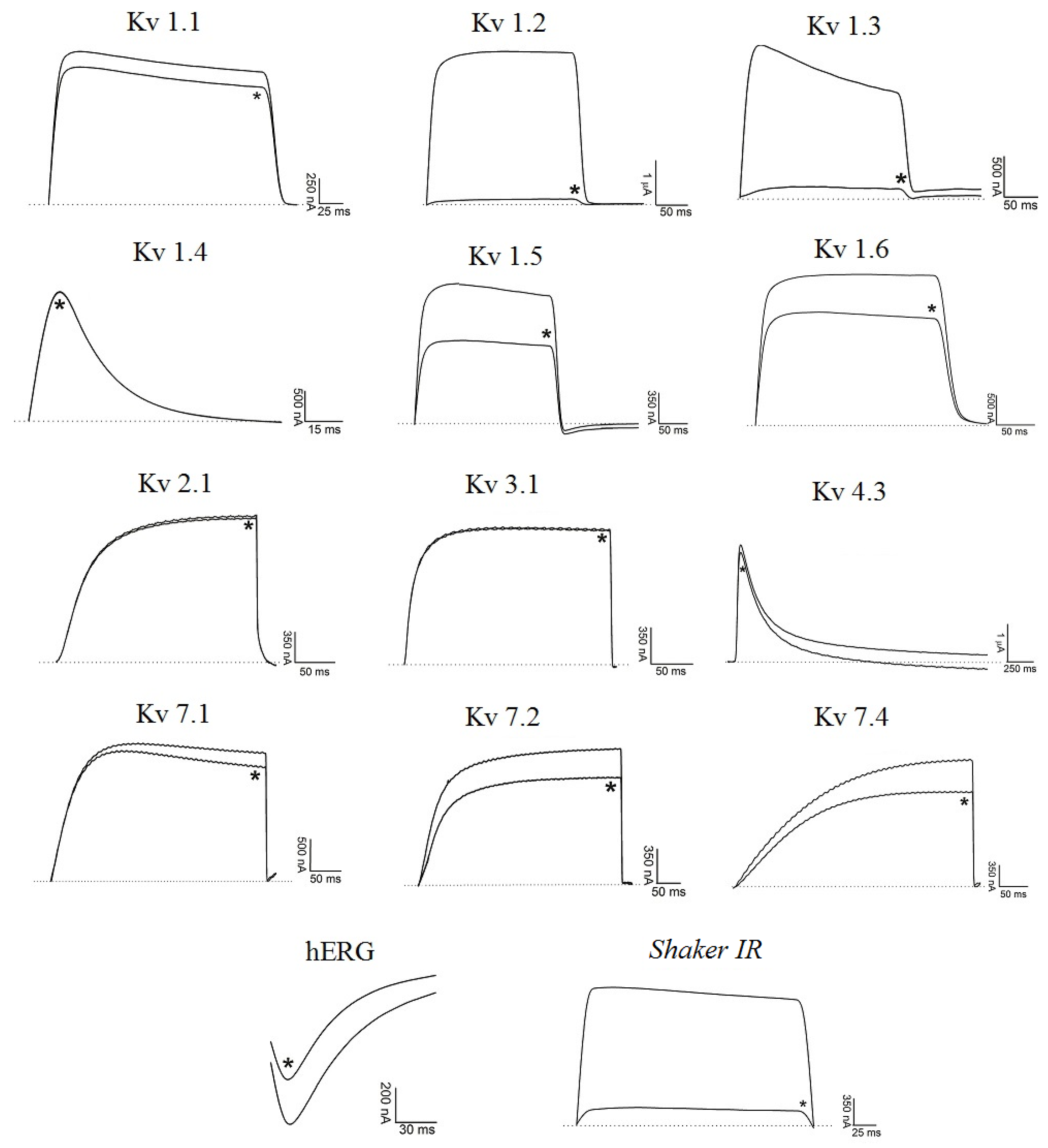
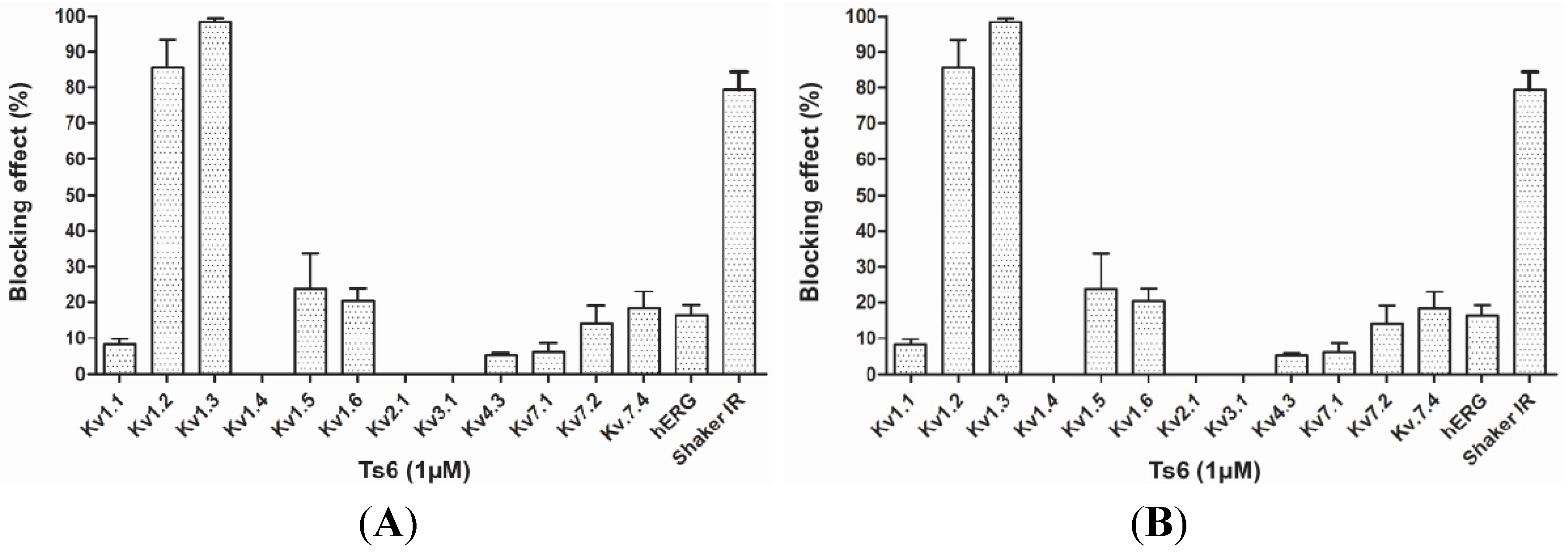
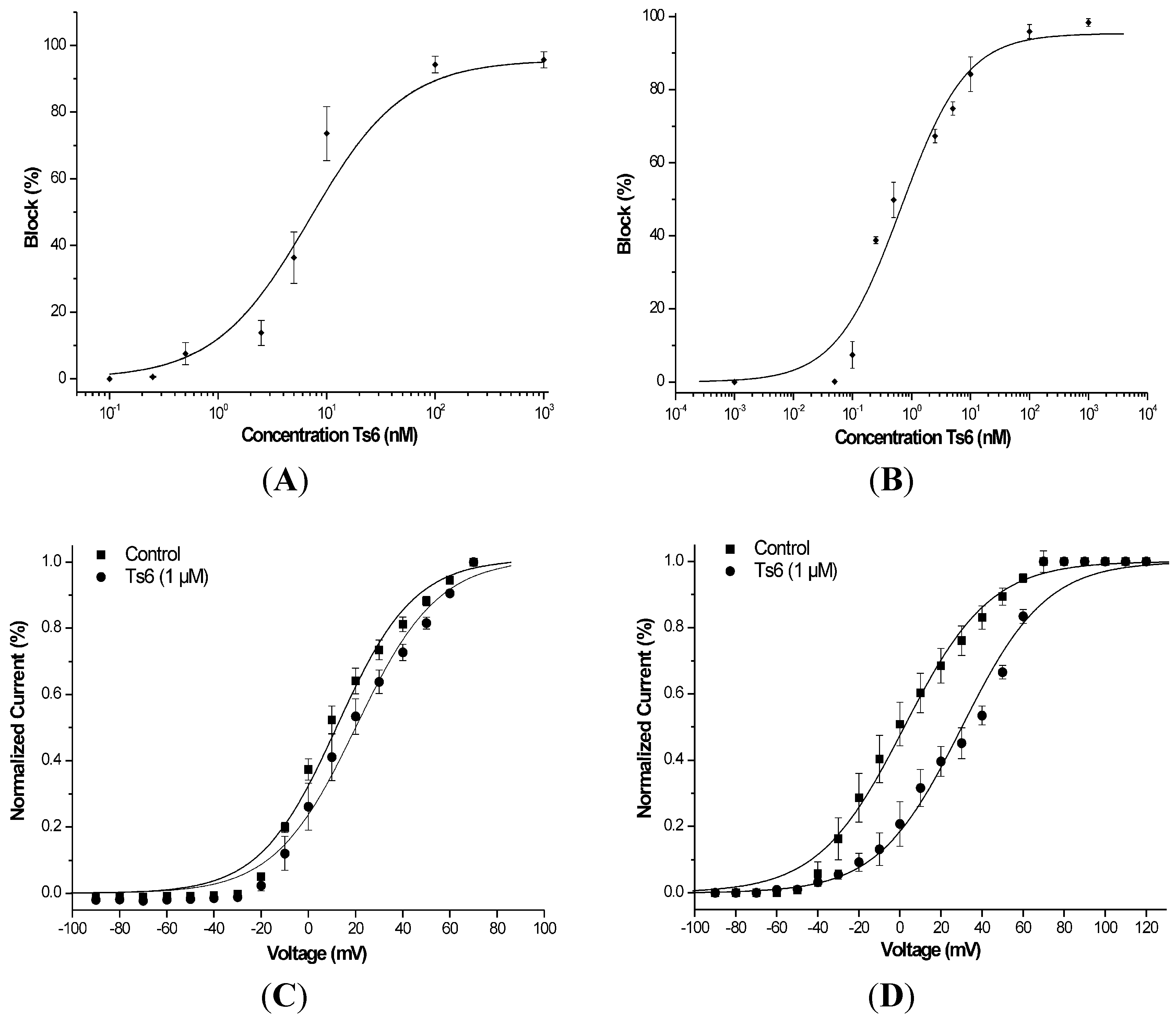
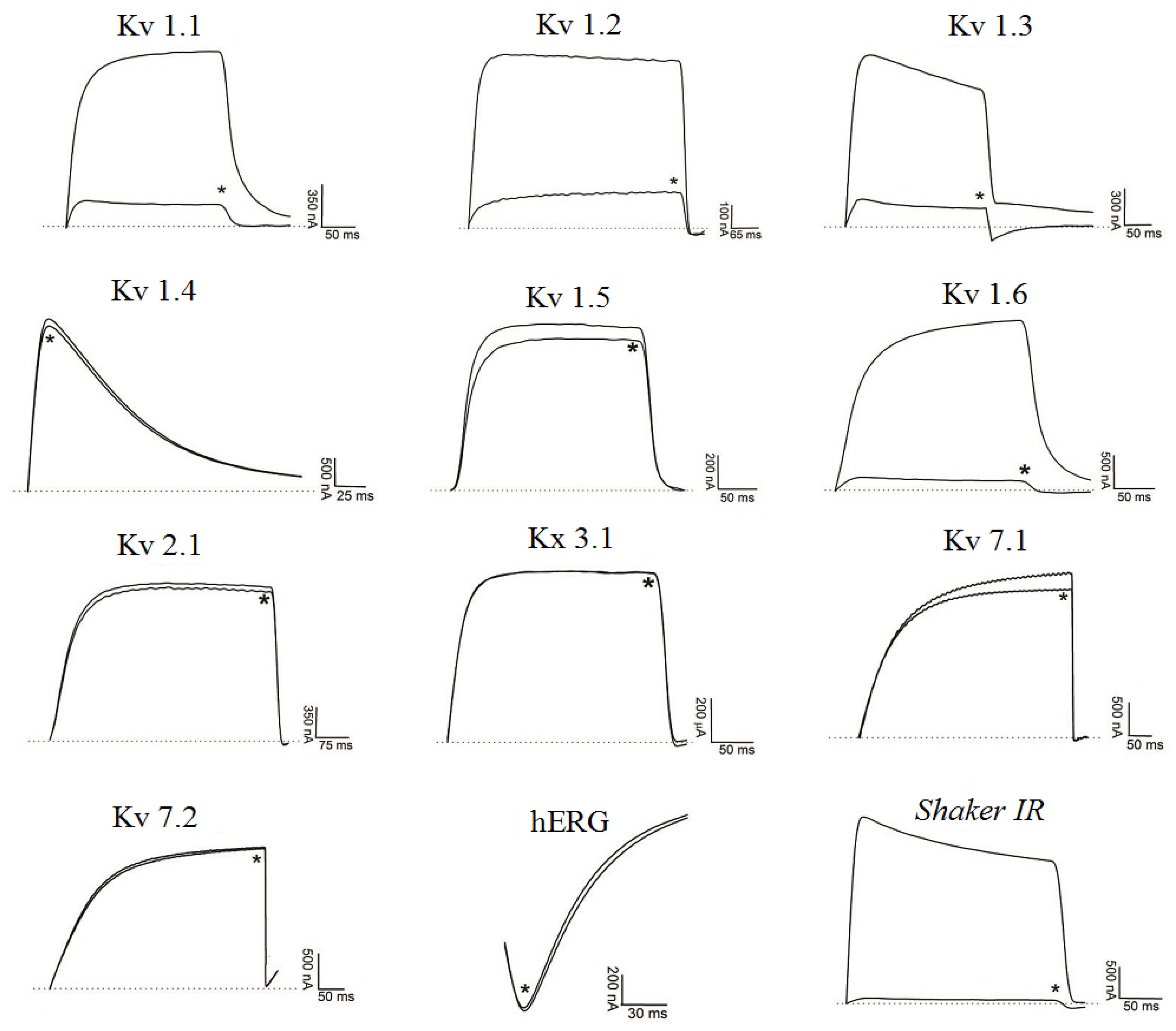
2.2. Electrophysiological Experiments
2.2.1. Ts6 Toxin



2.2.2. Ts7 Toxin

3. Discussion

4. Experimental Section
4.1. Tityus Serrulatus Venom
4.2. Toxins Isolation
4.2.1. Fast Protein Liquid Chromatography (FPLC) Using a CM-Cellulose-52 Column
4.2.2. Reversed-Phase (RP) FPLC Using a C18 Column
4.3. Biochemical Characterization of Toxins
4.4. Electrophysiological Experiments
4.4.1. Potassium Channel Expression
4.4.2. Electrophysiological Measurements
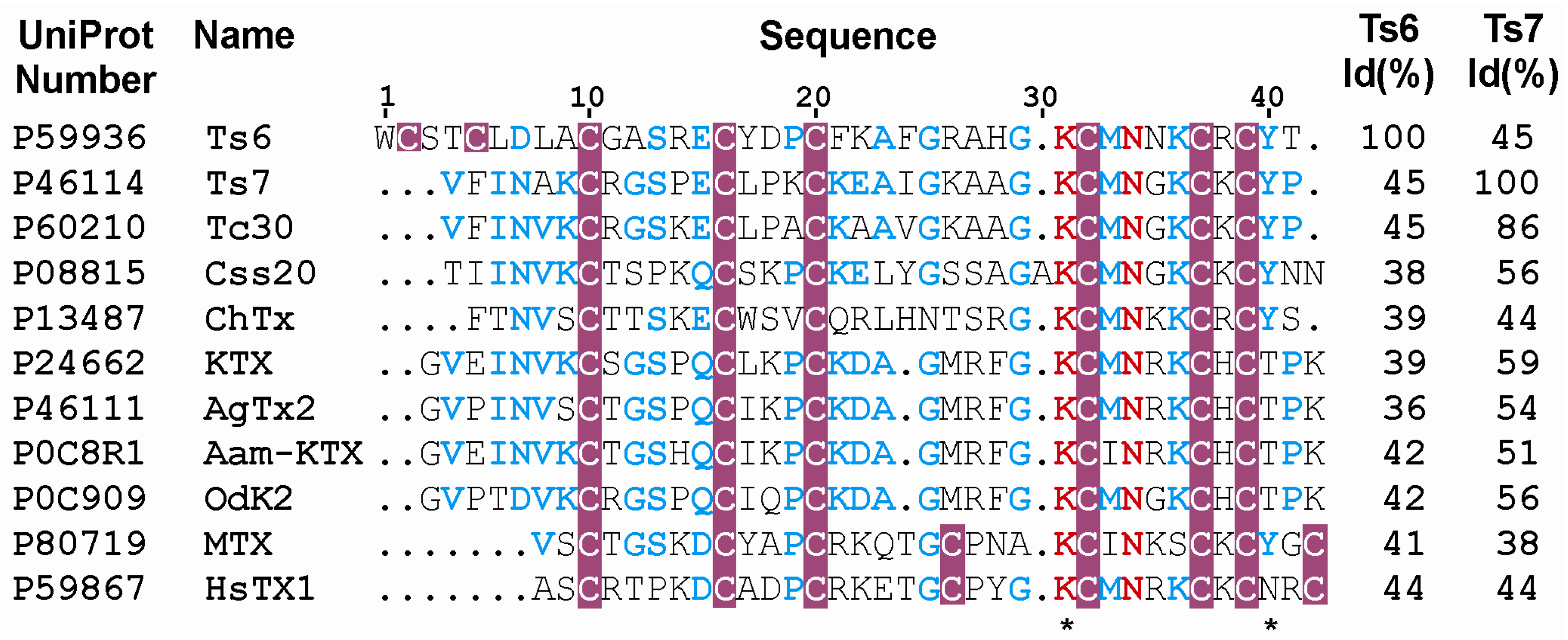
4.5. Toxins Sequence Alignment
5. Conclusion
Acknowledgments
Conflicts of Interest
Supplementary Data
Toxins Sequence Alignment and Comparison of Kv Block Effect
| Name | α-KTx | Sensitive Kv channels/IC50 | Insensitive Kv channels | Reference |
|---|---|---|---|---|
| Ts6 | 12.1 | Kv1.2/6.19 nM, Kv1.3/0.55 nM, Shaker IR/* | Kv1.1, Kv1.4, Kv1.5, Kv1.6, Kv2.1, Kv3.1, Kv4.3, Kv7.1, Kv7.2, Kv7.4, hERG, KCa | #,[14] |
| Ts7 | 4.1 | Kv1.1/*, Kv1.2/*, Kv1.3/*, Kv1.6/*, Shaker IR/* | Kv1.4, Kv1.5, Kv2.1, Kv3.1, Kv7.1, hEG | #,[24,25,26,71] |
| native sqKv/20 nM, | ||||
| cloned sqKv/10 nM | ||||
| heteromultimer Kv1.2/1.1 SSM/10 nM | ||||
| heteromultimer Kv1.2/1.4 EAM/0.7 nM | ||||
| Tc30 | 4.4 | Kv1.3/* , Shaker B/* | - | [49] |
| Css20 | 2.13 | Kv1.2/1.3 nM, Kv1.3/7.2 nM | Kv1.1, Kv1.4, Kv1.5, Kv2.1, Kv11.1 (hERG) KCa1, BKCa | [80] |
| ChTx | 1.1 | Shaker/*, Kv1.3/*, KCa | - | [81,82,83] |
| KTX | 3.1 | Kv1.1/1.1 nM–Kv1.2/25 nM–Kv1.3/0.1 nM | - | [58] |
| AgTx2 | 3.2 | Shaker/*, Kv1.1/*, Kv1.3/*, Kv1.6/* | Kv2.1 | [84] |
| Aam-KTX | 3.12 | Kv1.2/*, Kv1.3/* | Kv1.1 | [58] |
| OdK2 | 3.11 | Kv1.3/7.2 nM | Kv1.1, Kv1.2, Kv1.4, Kv1.5, Kv1.6, hERG , Shaker | [51] |
| MTX | 6.2 | Kv1.1/45 nM, Kv1.2/0.8 nM, Kv1.3/180 nM, Shaker/3.4 nM | - | [85,86] |
| HsTX1 | 6.3 | Kv1.1/7 nM, Kv1.3/12 pM, IK/625 nM | Kv1.2 | [50] |
References
- Verano-Braga, T.; Rocha-Resende, C.; Silva, D.M.; Ianzer, D.; Martin-Eauclaire, M.F.; Bougis, P.E.; de Lima, M.E.; Santos, R.A.; Pimenta, A.M. Tityus serrulatus hypotensins: A new family of peptides from scorpion venom. Biochem. Biophys. Res. Commun. 2008, 371, 515–520. [Google Scholar] [CrossRef]
- Pessini, A.C.; Takao, T.T.; Cavalheiro, E.C.; Vichnewski, W.; Sampaio, S.V.; Giglio, J.R.; Arantes, E.C. A hyaluronidase from tityus serrulatus scorpion venom: Isolation, characterization and inhibition by flavonoids. Toxicon 2001, 39, 1495–1504. [Google Scholar] [CrossRef]
- Ferreira, L.A.; Alves, E.W.; Henriques, O.B. Peptide t, a novel bradykinin potentiator isolated from tityus serrulatus scorpion venom. Toxicon 1993, 31, 941–947. [Google Scholar] [CrossRef]
- Cologna, C.T.; Marcussi, S.; Giglio, J.R.; Soares, A.M.; Arantes, E.C. Tityus serrulatus scorpion venom and toxins: An overview. Protein Pept. Lett. 2009, 16, 920–932. [Google Scholar] [CrossRef]
- Possani, L.D.; Alagon, A.C.; Fletcher, P.L., Jr.; Erickson, B.W. Purification and properties of mammalian toxins from the venom of brazilian scorpion tityus serrulatus lutz and mello. Arch. Biochem. Biophys. 1977, 180, 394–403. [Google Scholar] [CrossRef]
- Sampaio, S.V.; Laure, C.J.; Giglio, J.R. Isolation and characterization of toxic proteins from the venom of the brazilian scorpion tityus serrulatus. Toxicon 1983, 21, 265–277. [Google Scholar] [CrossRef]
- Possani, L.D.; Becerril, B.; Delepierre, M.; Tytgat, J. Scorpion toxins specific for Na+-channels. Eur. J. Biochem. 1999, 264, 287–300. [Google Scholar] [CrossRef]
- Gomez, M.V.; Diniz, C.R. Separation of toxic components from the brazillian scorpion tityus serrulatus venom. Memorias do Instituto Butantan 1966, 33, 899–902. [Google Scholar]
- Possani, L.D.; Martin, B.M.; Mochcamorales, J.; Svendsen, I. Purification and chemical characterization of the major toxins from the venom of the brazilian scorpion tityus serrulatus lutz and mello. Carlsberg Res. Commun. 1981, 46, 195–205. [Google Scholar] [CrossRef]
- Teixeira, C.E.; Ifa, D.R.; Corso, G.; Santagada, V.; Caliendo, G.; Antunes, E.; De Nucci, G. Sequence and structure-activity relationship of a scorpion venom toxin with nitrergic activity in rabbit corpus cavernosum. FASEB J. 2003, 17, 485–487. [Google Scholar]
- Arantes, E.C.; Prado, W.A.; Sampaio, S.V.; Giglio, J.R. A simplified procedure for the fractionation of tityus serrulatus venom: Isolation and partial characterization of tstx-iv, a new neurotoxin. Toxicon 1989, 27, 907–916. [Google Scholar] [CrossRef]
- Cologna, C.T.; Peigneur, S.; Rosa, J.C.; Selistre-de-Araujo, H.S.; Varanda, W.A.; Tytgat, J.; Arantes, E.C. Purification and characterization of ts15, the first member of a new alpha-ktx subfamily from the venom of the brazilian scorpion tityus serrulatus. Toxicon 2011, 58, 54–61. [Google Scholar] [CrossRef]
- Fletcher, P.L.; Fletcher, M.D.; Weninger, K.; Anderson, T.E.; Martin, B.M. Vesicle-associated membrane protein (vamp) cleavage by a new metalloprotease from the brazilian scorpion Tityus serrulatus. J. Biol. Chem. 2010, 285, 7405–7416. [Google Scholar]
- Novello, J.C.; Arantes, E.C.; Varanda, W.A.; Oliveira, B.; Giglio, J.R.; Marangoni, S. Tstx-IV, a short chain four-disulfide-bridged neurotoxin from tityus serrulatus venom which acts on Ca2+-activated K+ channels. Toxicon 1999, 37, 651–660. [Google Scholar] [CrossRef]
- Tytgat, J.; Chandy, K.G.; Garcia, M.L.; Gutman, G.A.; Martin-Eauclaire, M.F.; van der Walt, J.J.; Possani, L.D. A unified nomenclature for short-chain peptides isolated from scorpion venoms: Alpha-ktx molecular subfamilies. Trends Pharmacol. Sci. 1999, 20, 444–447. [Google Scholar] [CrossRef]
- Holaday, S.K., Jr.; Martin, B.M.; Fletcher, P.L., Jr.; Krishna, N.R. Nmr solution structure of butantoxi. Arch. Biochem. Biophys. 2000, 379, 18–27. [Google Scholar] [CrossRef]
- Pimenta, A.M.; Mansuelle, P.; Diniz, C.R.; Martin-Eauclaire, M.F. Covalent structure and some pharmacological features of native and cleaved alpha-ktx12–1, a four disulfide-bridged toxin from tityus serrulatus venom. J. Pept. Sci. 2003, 9, 132–140. [Google Scholar] [CrossRef]
- Coronas, F.V.; de Roodt, A.R.; Portugal, T.O.; Zamudio, F.Z.; Batista, C.V.; Gomez-Lagunas, F.; Possani, L.D. Disulfide bridges and blockage of shaker b K+-channels by another butantoxin peptide purified from the argentinean scorpion tityus trivittatus. Toxicon 2003, 41, 173–179. [Google Scholar] [CrossRef]
- Zoccal, K.F.; Bitencourt Cda, S.; Secatto, A.; Sorgi, C.A.; Bordon Kde, C.; Sampaio, S.V.; Arantes, E.C.; Faccioli, L.H. Tityus serrulatus venom and toxins ts1, ts2 and ts6 induce macrophage activation and production of immune mediators. Toxicon 2011, 57, 1101–1108. [Google Scholar] [CrossRef]
- Zoccal, K.F.; Bitencourt Cda, S.; Sorgi, C.A.; Bordon Kde, C.; Sampaio, S.V.; Arantes, E.C.; Faccioli, L.H. Ts6 and ts2 from tityus serrulatus venom induce inflammation by mechanisms dependent on lipid mediators and cytokine production. Toxicon 2013, 61, 1–10. [Google Scholar]
- Blaustein, M.P.; Rogowski, R.S.; Schneider, M.J.; Krueger, B.K. Polypeptide toxins from the venoms of old world and new world scorpions preferentially block different potassium channels. Mol. Pharmacol. 1991, 40, 932–942. [Google Scholar]
- Rogowski, R.S.; Krueger, B.K.; Collins, J.H.; Blaustein, M.P. Tityustoxin k alpha blocks voltage-gated noninactivating K+ channels and unblocks inactivating K+ channels blocked by alpha-dendrotoxin in synaptosomes. Proc. Natl. Acad. Sci. USA 1994, 91, 1475–1479. [Google Scholar] [CrossRef]
- Eccles, C.U.; Rogowski, R.S.; Gu, X.; Alger, B.E.; Blaustein, M.P. Tityustoxin-k alpha, from scorpion venom, blocks voltage-gated, non-inactivating potassium current in cultured central neurons. Neuropharmacology 1994, 33, 1523–1528. [Google Scholar] [CrossRef]
- Werkman, T.R.; Gustafson, T.A.; Rogowski, R.S.; Blaustein, M.P.; Rogawski, M.A. Tityustoxin-k alpha, a structurally novel and highly potent K+ channel peptide toxin, interacts with the alpha-dendrotoxin binding site on the cloned kv1.2 K+ channel. Mol. Pharmacol. 1993, 44, 430–436. [Google Scholar]
- Hopkins, W.F. Toxin and subunit specificity of blocking affinity of three peptide toxins for heteromultimeric, voltage-gated potassium channels expressed in xenopus oocytes. J. Pharmacol. Exp. Ther. 1998, 285, 1051–1060. [Google Scholar]
- Rodrigues, A.R.; Arantes, E.C.; Monje, F.; Stuhmer, W.; Varanda, W.A. Tityustoxin-k(alpha) blockade of the voltage-gated potassium channel kv1.3. Brit. J. Pharmacol. 2003, 139, 1180–1186. [Google Scholar] [CrossRef]
- Wulff, H.; Castle, N.A.; Pardo, L.A. Voltage-gated potassium channels as therapeutic targets. Nat. Rev. Drug Discov. 2009, 8, 982–1001. [Google Scholar] [CrossRef]
- Felipe, A.; Bielanska, J.; Comes, N.; Vallejo, A.; Roig, S.; Ramon, Y.C.S.; Condom, E.; Hernandez-Losa, J.; Ferreres, J.C. Targeting the voltage-dependent K(+) channels kv1.3 and kv1.5 as tumor biomarkers for cancer detection and prevention. Curr. Med. Chem. 2012, 19, 661–674. [Google Scholar] [CrossRef]
- Lam, J.; Wulff, H. The lymphocyte potassium channels kv1.3 and kca3.1 as targets for immunosuppression. Drug Dev. Res. 2011, 72, 573–584. [Google Scholar] [CrossRef]
- Yoshida, M.; Nakata, M.; Yamato, S.; Dezaki, K.; Sugawara, H.; Ishikawa, S.E.; Kawakami, M.; Yada, T.; Kakei, M. Voltage-dependent metabolic regulation of kv2.1 channels in pancreatic beta-cells. Biochem. Biophys. Res. Commun. 2010, 396, 304–309. [Google Scholar] [CrossRef]
- Robbins, C.A.; Tempel, B.L. Kv1.1 and kv1.2: Similar channels, different seizure models. Epilepsia 2012, 53 (Suppl. 1), 134–141. [Google Scholar]
- Gallego, M.; Alday, A.; Urrutia, J.; Casis, O. Transient outward potassium channel regulation in healthy and diabetic hearts. Can. J. Physiol. Pharmacol. 2009, 87, 77–83. [Google Scholar] [CrossRef]
- Shepherd, A.J.; Loo, L.; Gupte, R.P.; Mickle, A.D.; Mohapatra, D.P. Distinct modifications in kv2.1 channel via chemokine receptor cxcr4 regulate neuronal survival-death dynamics. J. Neurosci. 2012, 32, 17725–17739. [Google Scholar] [CrossRef]
- He, F.Z.; McLeod, H.L.; Zhang, W. Current pharmacogenomic studies on herg potassium channels. Trends Mol. Med. 2013, 4, 227–228. [Google Scholar]
- Abdel-Mottaleb, Y.; Corzo, G.; Martin-Eauclaire, M.F.; Satake, H.; Ceard, B.; Peigneur, S.; Nambaru, P.; Bougis, P.E.; Possani, L.D.; Tytgat, J. A common “hot spot” confers herg blockade activity to alpha-scorpion toxins affecting K+ channels. Biochem. Pharmacol. 2008, 76, 805–815. [Google Scholar] [CrossRef]
- Papazian, D.M.; Schwarz, T.L.; Tempel, B.L.; Jan, Y.N.; Jan, L.Y. Cloning of genomic and complementary DNA from shaker, a putative potassium channel gene from drosophila. Science 1987, 237, 749–753. [Google Scholar]
- Tempel, B.L.; Papazian, D.M.; Schwarz, T.L.; Jan, Y.N.; Jan, L.Y. Sequence of a probable potassium channel component encoded at shaker locus of drosophila. Science 1987, 237, 770–775. [Google Scholar]
- Pongs, O.; Kecskemethy, N.; Muller, R.; Krahjentgens, I.; Baumann, A.; Kiltz, H.H.; Canal, I.; Llamazares, S.; Ferrus, A. Shaker encodes a family of putative potassium channel proteins in the nervous-system of drosophila. Embo J. 1988, 7, 1087–1096. [Google Scholar]
- Zhu, S.; Peigneur, S.; Gao, B.; Luo, L.; Jin, D.; Zhao, Y.; Tytgat, J. Molecular diversity and functional evolution of scorpion potassium channel toxins. Mol. Cell. Proteomics 2011, 10. [Google Scholar] [CrossRef]
- Heginbotham, L.; MacKinnon, R. The aromatic binding site for tetraethylammonium ion on potassium channels. Neuron 1992, 8, 483–491. [Google Scholar] [CrossRef]
- Chen, Z.; Hu, Y.; Han, S.; Yin, S.; He, Y.; Wu, Y.; Cao, Z.; Li, W. Imktx1, a new kv1.3 channel blocker with a unique primary structure. J. Biochem. Mol. Toxicol. 2011, 25, 244–251. [Google Scholar] [CrossRef]
- Brew, H.M.; Gittelman, J.X.; Silverstein, R.S.; Hanks, T.D.; Demas, V.P.; Robinson, L.C.; Robbins, C.A.; McKee-Johnson, J.; Chiu, S.Y.; Messing, A.; et al. Seizures and reduced life span in mice lacking the potassium channel subunit kv1.2, but hypoexcitability and enlarged kv1 currents in auditory neurons. J. Neurophys. 2007, 98, 1501–1525. [Google Scholar] [CrossRef]
- Xie, G.; Harrison, J.; Clapcote, S.J.; Huang, Y.; Zhang, J.Y.; Wang, L.Y.; Roder, J.C. A new kv1.2 channelopathy underlying cerebellar ataxia. J. Biol. Chem. 2010, 285, 32160–32173. [Google Scholar]
- Fulton, S.; Thibault, D.; Mendez, J.A.; Lahaie, N.; Tirotta, E.; Borrelli, E.; Bouvier, M.; Tempel, B.L.; Trudeau, L.E. Contribution of kv1.2 voltage-gated potassium channel to d2 autoreceptor regulation of axonal dopamine overflow. J. Biol. Chem. 2011, 286, 9360–9372. [Google Scholar] [CrossRef]
- Ghanshani, S.; Wulff, H.; Miller, M.J.; Rohm, H.; Neben, A.; Gutman, G.A.; Cahalan, M.D.; Chandy, K.G. Up-regulation of the ikca1 potassium channel during t-cell activation. Molecular mechanism and functional consequences. J. Biol. Chem. 2000, 275, 37137–37149. [Google Scholar] [CrossRef]
- Hu, L.; Gocke, A.R.; Knapp, E.; Rosenzweig, J.M.; Grishkan, I.V.; Baxi, E.G.; Zhang, H.; Margolick, J.B.; Whartenby, K.A.; Calabresi, P.A. Functional blockade of the voltage-gated potassium channel kv1.3 mediates reversion of t effector to central memory lymphocytes through smad3/p21cip1 signaling. J. Biol. Chem. 2012, 287, 1261–1268. [Google Scholar] [CrossRef]
- Wulff, H.; Calabresi, P.A.; Allie, R.; Yun, S.; Pennington, M.; Beeton, C.; Chandy, K.G. The voltage-gated kv1.3 K(+) channel in effector memory t cells as new target for ms. J. Clin. Investig. 2003, 111, 1703–1713. [Google Scholar] [CrossRef]
- Beeton, C.; Chandy, K.G. Potassium channels, memory t cells, and multiple sclerosis. Neurosci. 2005, 11, 550–562. [Google Scholar]
- Batista, C.V.; Gomez-Lagunas, F.; Rodriguez de la Vega, R.C.; Hajdu, P.; Panyi, G.; Gaspar, R.; Possani, L.D. Two novel toxins from the amazonian scorpion tityus cambridgei that block kv1.3 and shaker b K(+)-channels with distinctly different affinities. Biochim. Biophys. Acta 2002, 1601, 123–131. [Google Scholar] [CrossRef]
- Lebrun, B.; Rimi-Lebrun, R.; Martin-Euclaire, M.F.; Yasuda, A.; Ishiguro, M.; Oyama, Y.; Pongs, O.; Nakajima, T. A four-disulphide-bridged toxin, with high affinity towards voltage-gated K+ channels, isolated from heterometrus spinnifer (scorpionidae) venom. Bichem. J. 1997, 328, 321–327. [Google Scholar]
- Abdel-Mottaleb, Y.; Vandendriessche, T.; Clynen, E.; Landuyt, B.; Jalali, A.; Vatanpour, H.; Schoofs, L.; Tytgat, J. Odk2, a kv1.3 channel-selective toxin from the venom of the iranian scorpion odonthobuthus doriae. Toxicon 2008, 51, 1424–1430. [Google Scholar] [CrossRef]
- Pennington, M.W.; Byrnes, M.E.; Zaydenberg, I.; Khaytin, I.; de Chastonay, J.; Krafte, D.S.; Hill, R.; Mahnir, V.M.; Volberg, W.A.; Gorczyca, W.; et al. Chemical synthesis and characterization of shk toxin: A potent potassium channel inhibitor from a sea anemone. Int. J. Pept. Protein Res. 1995, 46, 354–358. [Google Scholar]
- Beeton, C.; Pennington, M.W.; Wulff, H.; Singh, S.; Nugent, D.; Crossley, G.; Khaytin, I.; Calabresi, P.A.; Chen, C.Y.; Gutman, G.A.; et al. Targeting effector memory t cells with a selective peptide inhibitor of kv1.3 channels for therapy of autoimmune diseases. Mol. Pharmacol. 2005, 67, 1369–1381. [Google Scholar] [CrossRef]
- Lin, C.S.; Boltz, R.C.; Blake, J.T.; Nguyen, M.; Talento, A.; Fischer, P.A.; Springer, M.S.; Sigal, N.H.; Slaughter, R.S.; Garcia, M.L.; et al. Voltage-gated potassium channels regulate calcium-dependent pathways involved in human t lymphocyte activation. J. Exp. Med. 1993, 177, 637–645. [Google Scholar] [CrossRef]
- Koo, G.C.; Blake, J.T.; Talento, A.; Nguyen, M.; Lin, S.; Sirotina, A.; Shah, K.; Mulvany, K.; Hora, D., Jr.; Cunningham, P.; et al. Blockade of the voltage-gated potassium channel kv1.3 inhibits immune responses in vivo. J. Immunol. 1997, 158, 5120–5128. [Google Scholar]
- Middleton, R.E.; Sanchez, M.; Linde, A.R.; Bugianesi, R.M.; Dai, G.; Felix, J.P.; Koprak, S.L.; Staruch, M.J.; Bruguera, M.; Cox, R.; et al. Substitution of a single residue in stichodactyla helianthus peptide, shk-dap22, reveals a novel pharmacological profile. Biochemistry 2003, 42, 13698–13707. [Google Scholar] [CrossRef]
- Lutimax Homepage. Available online: http://www.lutimax.com (accessed on 1 October 2014).
- Abbas, N.; Belghazi, M.; Abdel-Mottaleb, Y.; Tytgat, J.; Bougis, P.E.; Martin-Eauclaire, M.F. A new kaliotoxin selective towards kv1.3 and kv1.2 but not kv1.1 channels expressed in oocytes. Biochem. Biophys. Res. Commun. 2008, 376, 525–530. [Google Scholar] [CrossRef]
- Jouirou, B.; Mouhat, S.; Andreotti, N.; De Waard, M.; Sabatier, J.M. Toxin determinants required for interaction with voltage-gated K+ channels. Toxicon 2004, 43, 909–914. [Google Scholar] [CrossRef]
- Dauplais, M.; Lecoq, A.; Song, J.; Cotton, J.; Jamin, N.; Gilquin, B.; Roumestand, C.; Vita, C.; de Medeiros, C.L.; Rowan, E.G.; et al. On the convergent evolution of animal toxins. Conservation of a diad of functional residues in potassium channel-blocking toxins with unrelated structures. J. Biol. Chem. 1997, 272, 4302–4309. [Google Scholar] [CrossRef]
- Banerjee, A.; Lee, A.; Campbell, E.; Mackinnon, R. Structure of a pore-blocking toxin in complex with a eukaryotic voltage-dependent K(+) channel. eLife 2013, 2, e00594. [Google Scholar] [CrossRef]
- Rodriguez de la Vega, R.C.; Possani, L.D. Current views on scorpion toxins specific for K+-channels. Toxicon 2004, 43, 865–875. [Google Scholar] [CrossRef]
- Rates, B.; Verano-Braga, T.; Santos, D.M.; Nunes, K.P.; Pimenta, A.M.; De Lima, M.E. From the stretcher to the pharmacy’s shelf: Drug leads from medically important brazilian venomous arachnid species. Inflamm. Allergy Drug Targets 2011, 10, 411–419. [Google Scholar] [CrossRef]
- Mouhat, S.; Mosbah, A.; Visan, V.; Wulff, H.; Delepierre, M.; Darbon, H.; Grissmer, S.; De Waard, M.; Sabatier, J.M. The “functional” dyad of scorpion toxin pi1 is not itself a prerequisite for toxin binding to the voltage-gated kv1.2 potassium channels. Biochem. J. 2004, 377, 25–36. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, M.; Jiang, M.; Tseng, G.N. Structural and functional role of the extracellular s5-p linker in the herg potassium channel. J. Gen. Phys. 2002, 120, 723–737. [Google Scholar] [CrossRef]
- Pardo-Lopez, L.; Zhang, M.; Liu, J.; Jiang, M.; Possani, L.D.; Tseng, G.N. Mapping the binding site of a human ether-a-go-go-related gene-specific peptide toxin (ergtx) to the channel’s outer vestibule. J. Biol. Chem. 2002, 277, 16403–16411. [Google Scholar]
- Frenal, K.; Xu, C.Q.; Wolff, N.; Wecker, K.; Gurrola, G.B.; Zhu, S.Y.; Chi, C.W.; Possani, L.D.; Tytgat, J.; Delepierre, M. Exploring structural features of the interaction between the scorpion toxincnerg1 and erg K+ channels. Proteins 2004, 56, 367–375. [Google Scholar] [CrossRef]
- Oyama, S., Jr.; Pristovsek, P.; Franzoni, L.; Pertinhez, T.A.; Schinina, E.; Lucke, C.; Ruterjans, H.; Arantes, E.C.; Spisni, A. Probing the ph-dependent structural features of alpha-ktx12.1, a potassium channel blocker from the scorpion Tityus serrulatus. Protein Sci. 2005, 14, 1025–1038. [Google Scholar] [CrossRef]
- Naranjo, D.; Niller, C.A. Strongly interacting pair of residues on the contact surface of charybdotoxin and a shaker K+ channel. Neuron 1996, 16, 123–130. [Google Scholar] [CrossRef]
- Blanc, E.; Lecomte, C.; Rietschoten, J.V.; Sabatier, J.M.; Darbon, H. Solution structure of tskapa, a charybdotoxin-like scorpion toxin from Tityus serrulatus with high affinity for apamin-sensitive Ca2+–activated K+ channels. Proteins 1997, 29, 359–369. [Google Scholar] [CrossRef]
- Ellis, K.C.; Tenenholz, T.C.; Jerng, H.; Hayhurst, M.; Dudlak, C.S.; Gilly, W.F.; Blaustein, M.P.; Weber, D.J. Interaction of a toxin from the scorpion tityus serrulatus with a cloned K+ channel from squid (sqkv1a). Biochemistry 2001, 40, 5942–5953. [Google Scholar] [CrossRef]
- Goldstein, S.A.; Miller, C. A point mutation in a shaker K+ channel changes its charybdotoxin binding site from low to high affinity. Biophys. J. 1992, 62, 5–7. [Google Scholar] [CrossRef]
- Bergeron, Z.L.; Bingham, J.P. Scorpion toxins specific for potassium (K+) channels: A historical overview of peptide bioengineering. Toxins 2012, 4, 1082–1119. [Google Scholar] [CrossRef]
- Possani, L.D.; Merino, E.; Corona, M.; Bolivar, F.; Becerril, B. Peptides and genes coding for scorpion toxins that affect ion-channels. Biochimie 2000, 82, 861–868. [Google Scholar] [CrossRef]
- Lowe, R.M.; Farrell, P.M. A portable device for the electrical extraction of scorpion venom. Toxicon 2011, 57, 244–247. [Google Scholar] [CrossRef]
- Edman, P.; Begg, G. A protein sequenator. Eur. J. Biochem. 1967, 1, 80–91. [Google Scholar] [CrossRef]
- Tytgat, J.; Maertens, C.; Daenens, P. Effect of fluoxetine on a neuronal, voltage-dependent potassium channel (kv1.1). Brit. J. Pharmacol. 1997, 122, 1417–1424. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; MacGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Gouet, P.; Courcelle, E.; Stuart, D.I.; Métoz, F. Espript: Analysis of multiple sequence alignments in postscript. Bioinformatics 1999, 15, 305–308. [Google Scholar] [CrossRef]
- Corzo, G.; Papp, F.; Varga, Z.; Barraza, O.; Espino-Solis, P.G.; Rodriguez de la Vega, R.C.; Gaspar, R.; Panyi, G.; Possani, L.D. A selective blocker of kv1.2 and kv1.3 potassium channels from the venom of the scorpion centruroides suffusus suffusus. Biochem. Pharmacol. 2008, 76, 1142–1154. [Google Scholar] [CrossRef]
- Anderson, C.S.; MacKinnon, R.; Smith, C.; Miller, C. Charybdotoxin block of single Ca2+-activated K+ channels. Effects of channel gating, voltage, and ionic strength. J. Gen. Phys. 1988, 91, 317–333. [Google Scholar] [CrossRef]
- MacKinnon, R.; Reinhart, P.H.; White, M.M. Charybdotoxin block of shaker K+ channels suggests that different types of K+ channels share common structural features. Neuron 1988, 1, 997–1001. [Google Scholar] [CrossRef]
- Khabiri, M.; Nikouee, A.; Cwiklik, L.; Grissmer, S.; Ettrich, R. Charybdotoxin unbinding from the mkv1.3 potassium channel: A combined computational and experimental study. J. Phys. Chem. B 2011, 115, 11490–11500. [Google Scholar] [CrossRef]
- Garcia, M.L.; Garcia-Calvo, M.; Hidalgo, P.; Lee, A.; MacKinnon, R. Purification and characterization of three inhibitors of voltage-dependent K+ channels from leiurus quinquestriatus var. Hebraeus venom. Biochemistry 1994, 33, 6834–6839. [Google Scholar] [CrossRef]
- Kharrat, R.; Mansuelle, P.; Sampieri, F.; Crest, M.; Oughideni, R.; Van Rietschoten, J.; Martin-Eauclaire, M.F.; Rochat, H.; El Ayeb, M. Maurotoxin, a four disulfide bridge toxin from scorpio maurus venom: Purification, structure and action on potassium channels. FEBS Lett. 1997, 406, 284–290. [Google Scholar] [CrossRef]
- Avdonin, V.; Nolan, B.; Sabatier, J.M.; De Waard, M.; Hoshi, T. Mechanisms of maurotoxin action on shaker potassium channels. Biophys. J. 2000, 79, 776–787. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cerni, F.A.; Pucca, M.B.; Peigneur, S.; Cremonez, C.M.; Bordon, K.C.F.; Tytgat, J.; Arantes, E.C. Electrophysiological Characterization of Ts6 and Ts7, K+ Channel Toxins Isolated through an Improved Tityus serrulatus Venom Purification Procedure. Toxins 2014, 6, 892-913. https://doi.org/10.3390/toxins6030892
Cerni FA, Pucca MB, Peigneur S, Cremonez CM, Bordon KCF, Tytgat J, Arantes EC. Electrophysiological Characterization of Ts6 and Ts7, K+ Channel Toxins Isolated through an Improved Tityus serrulatus Venom Purification Procedure. Toxins. 2014; 6(3):892-913. https://doi.org/10.3390/toxins6030892
Chicago/Turabian StyleCerni, Felipe A., Manuela B. Pucca, Steve Peigneur, Caroline M. Cremonez, Karla C. F. Bordon, Jan Tytgat, and Eliane C. Arantes. 2014. "Electrophysiological Characterization of Ts6 and Ts7, K+ Channel Toxins Isolated through an Improved Tityus serrulatus Venom Purification Procedure" Toxins 6, no. 3: 892-913. https://doi.org/10.3390/toxins6030892
APA StyleCerni, F. A., Pucca, M. B., Peigneur, S., Cremonez, C. M., Bordon, K. C. F., Tytgat, J., & Arantes, E. C. (2014). Electrophysiological Characterization of Ts6 and Ts7, K+ Channel Toxins Isolated through an Improved Tityus serrulatus Venom Purification Procedure. Toxins, 6(3), 892-913. https://doi.org/10.3390/toxins6030892