
Triggering Apoptotic Death of Human Epidermal Keratinocytes by Malic Acid: Involvement of Endoplasmic Reticulum Stress- and Mitochondria-Dependent Signaling Pathways


 , and
, and 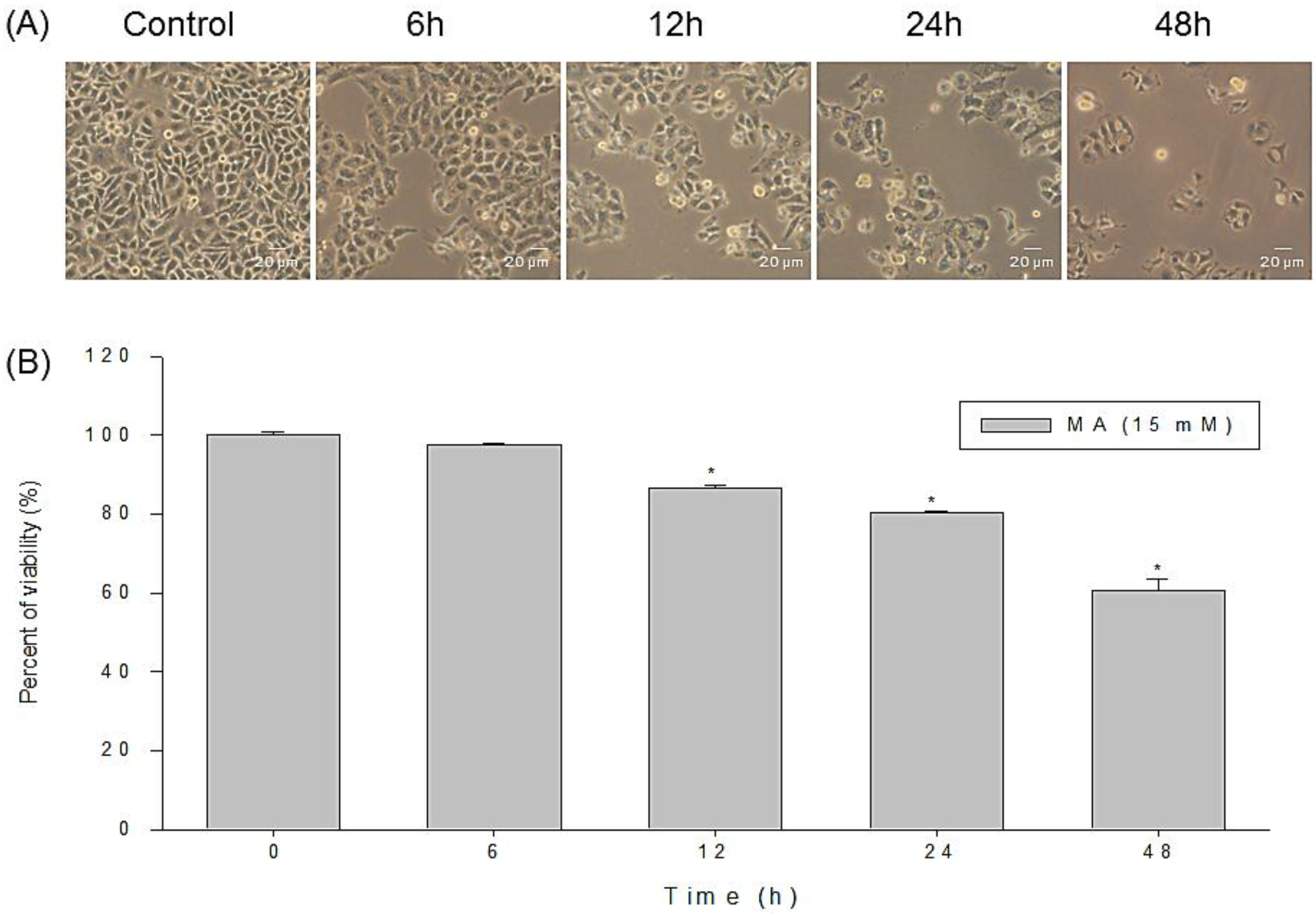{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Anti-Proliferative Effects of MA on Cell Morphology and Viability of HaCaT Cells

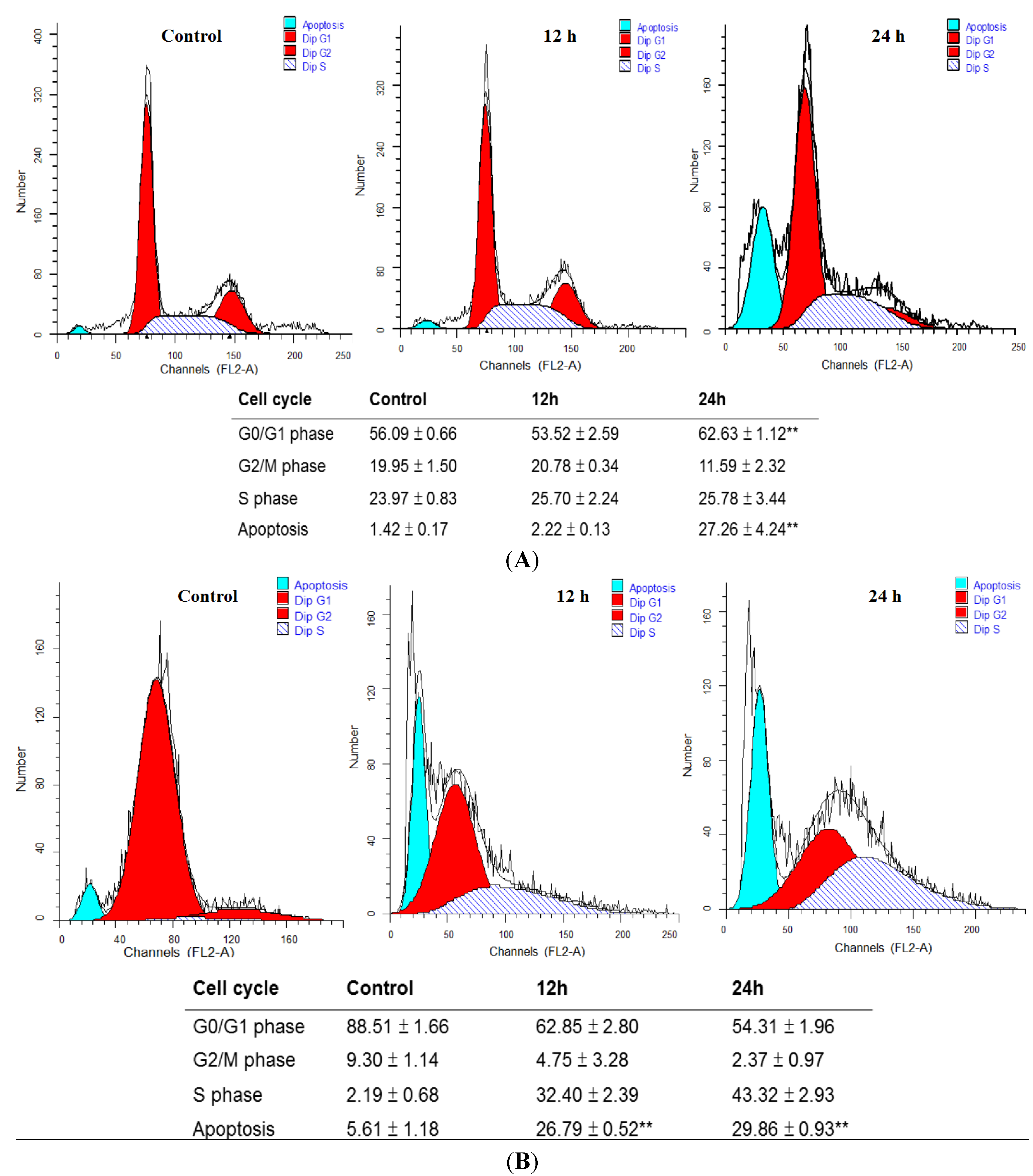
2.2. MA Induced Cell Cycle Arrest and Apoptosis in HaCaT Cells and NHEKs
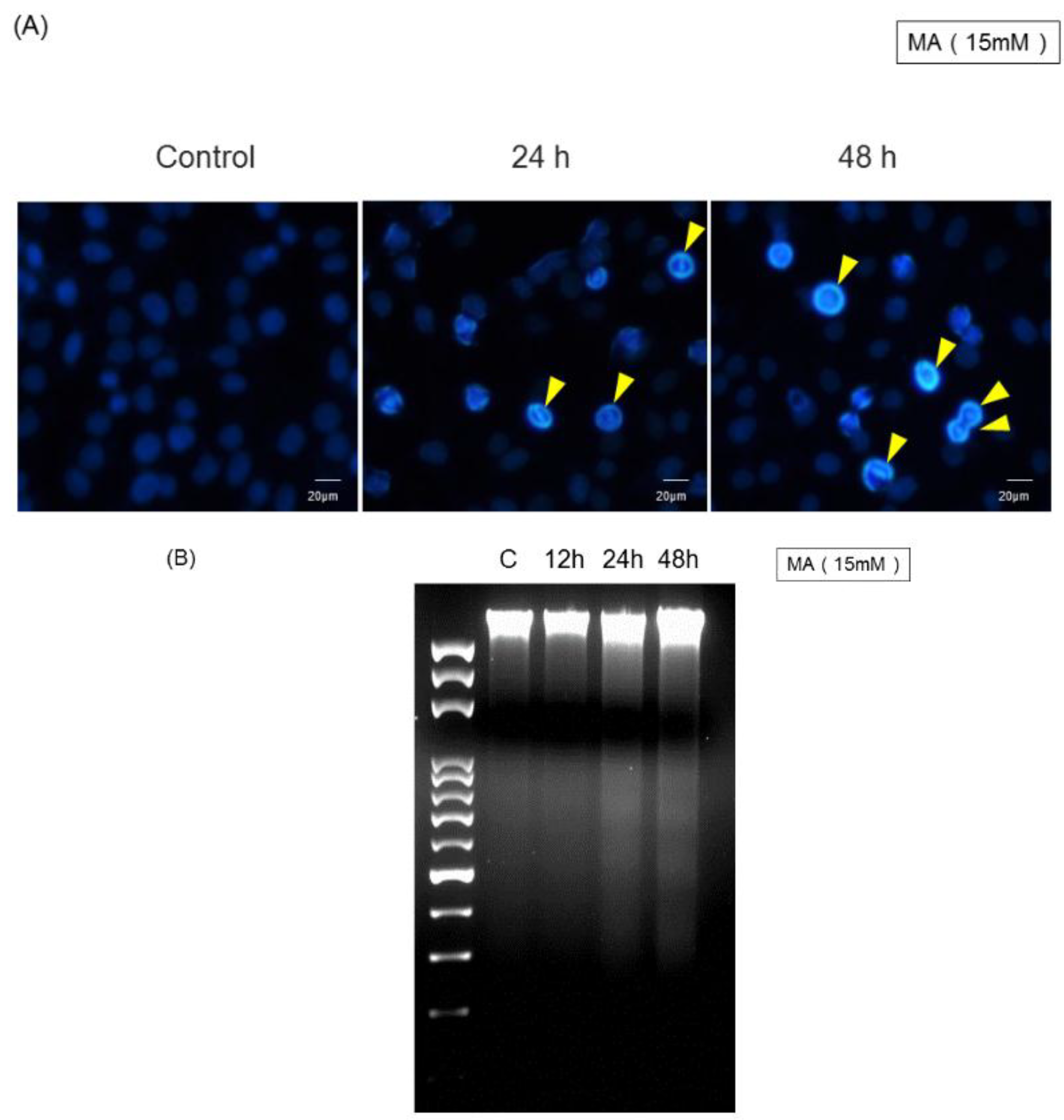
2.3. MA Induced Cell Damage Examined by DAPI Staining and DNA Fragmentation


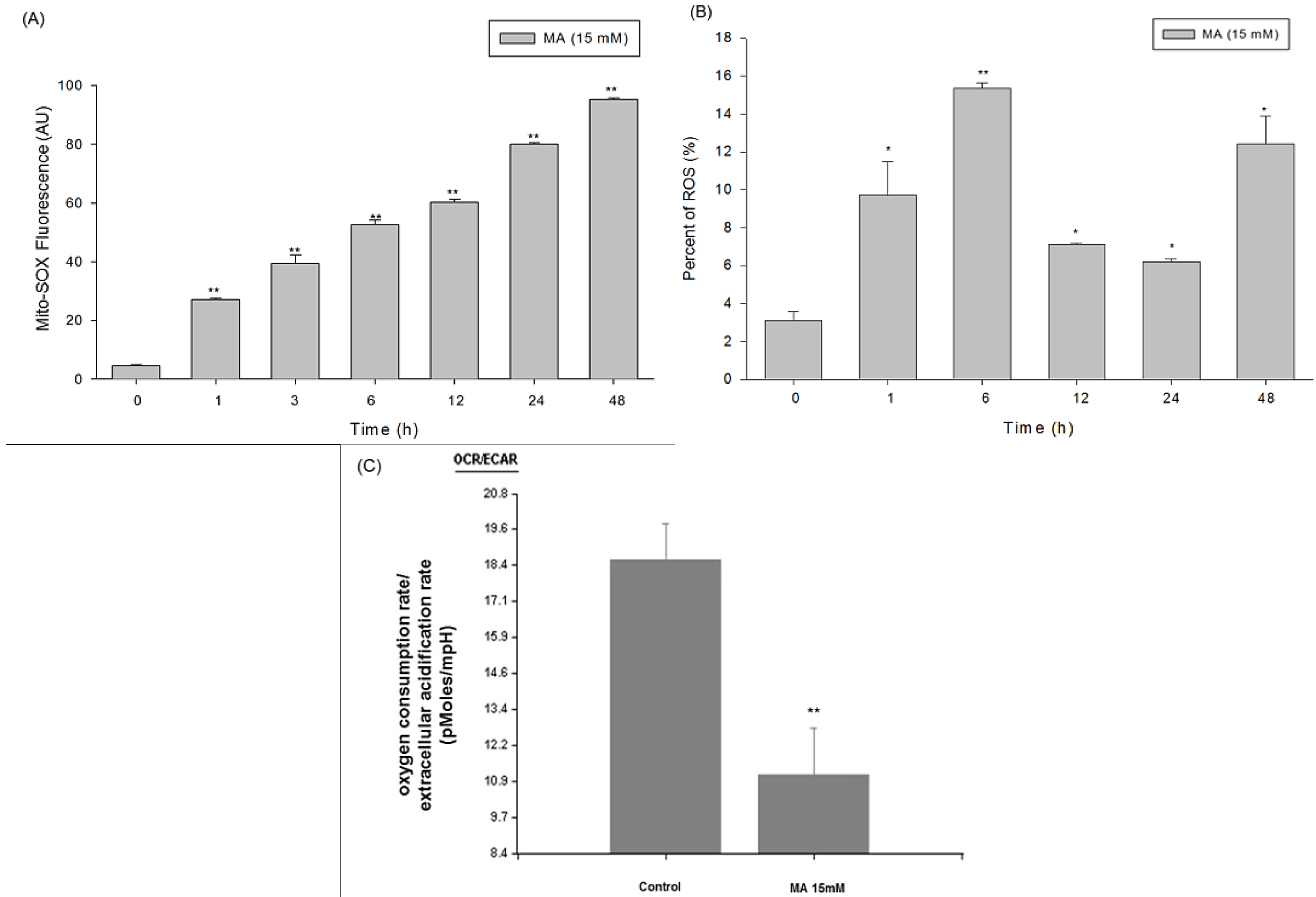
2.4. Intracellular Superoxide Dynamics of HaCaT Cells Incubated in Malic Acid
2.5. Measurement of Oxygen Consumption Rate (OCR) and Extracellular Acidification Rate (ECAR) of Malic Acid-Treated HaCaT Cells by Seahorse XF24 Analyzer

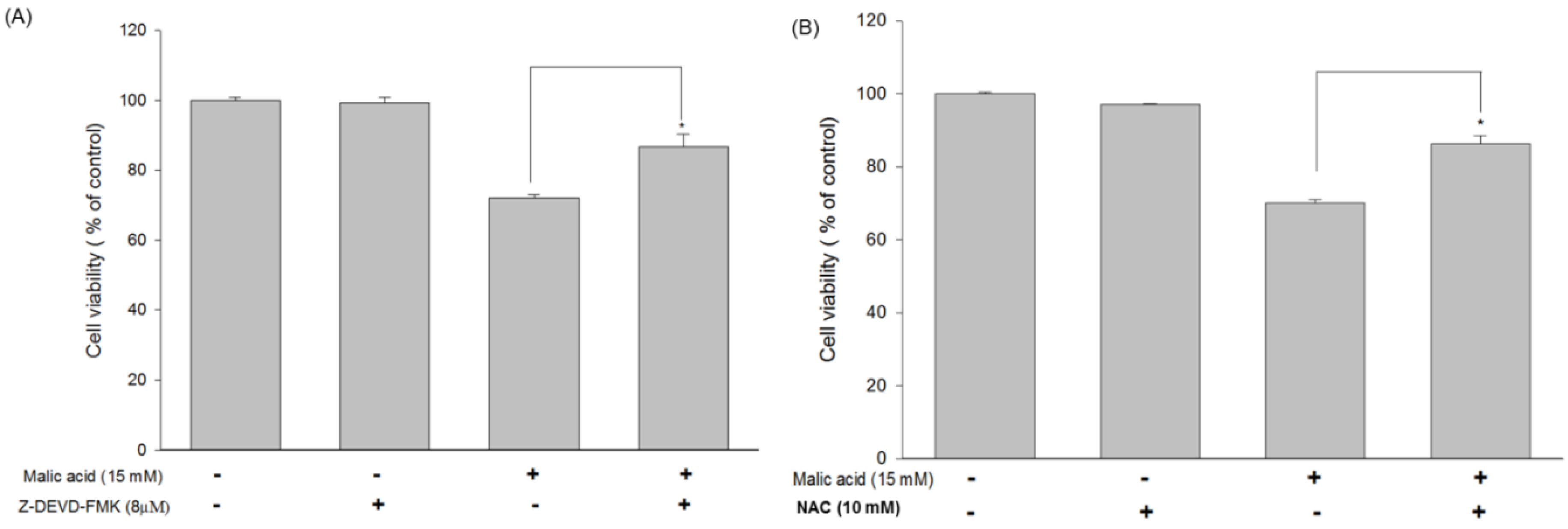
2.6. Effects of MA on Caspase-3 and Inhibition of MA-Induced Apoptosis by the Caspase-3 Inhibitor Z-DEVD-FMK in HaCaT Cells



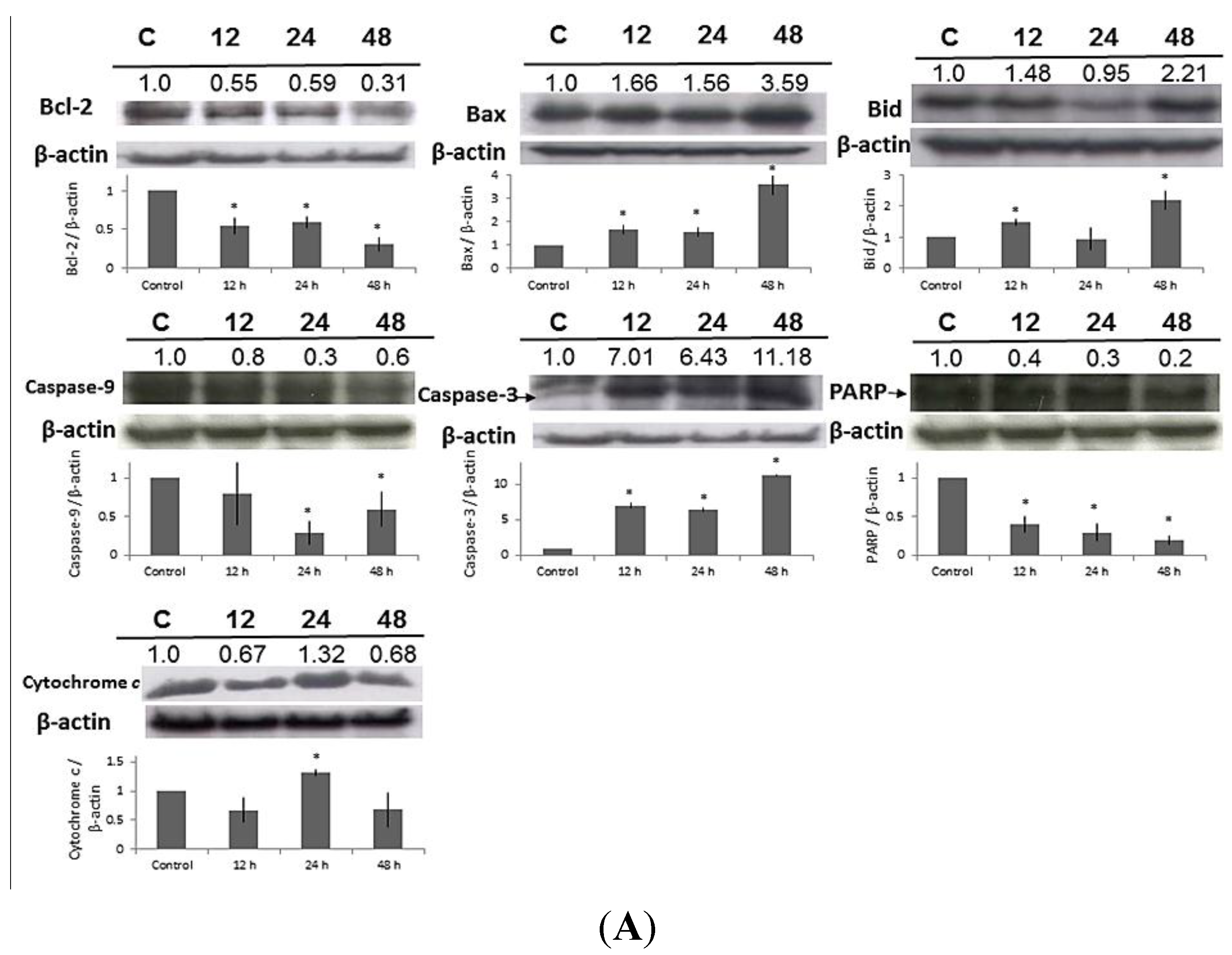
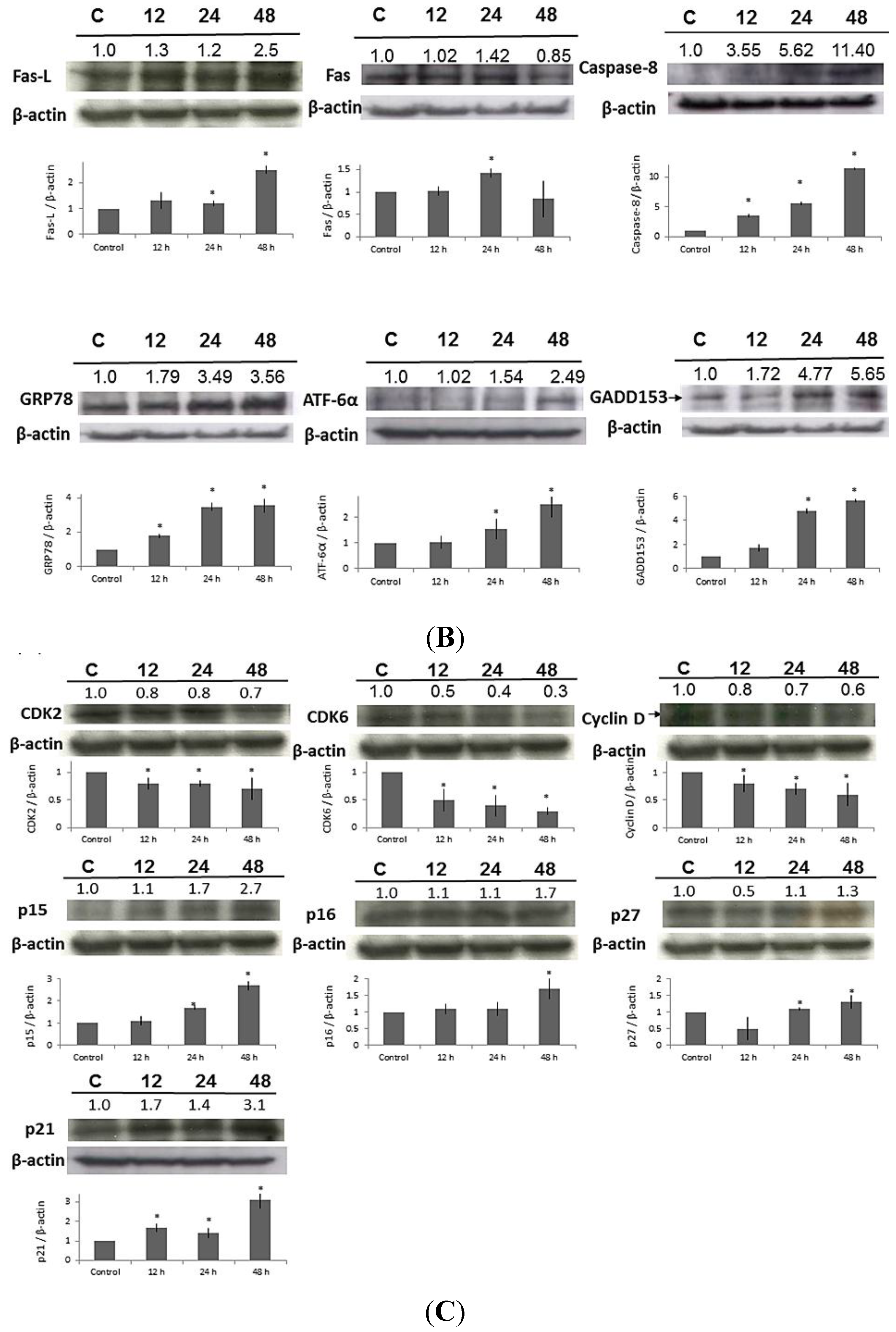
2.7. Effect of MA on the Expressions of Apoptosis-Related Proteins in HaCaT Cells
2.8. Effect of MA on the Expressions of Cell Cycle-Related Proteins in HaCaT Cells
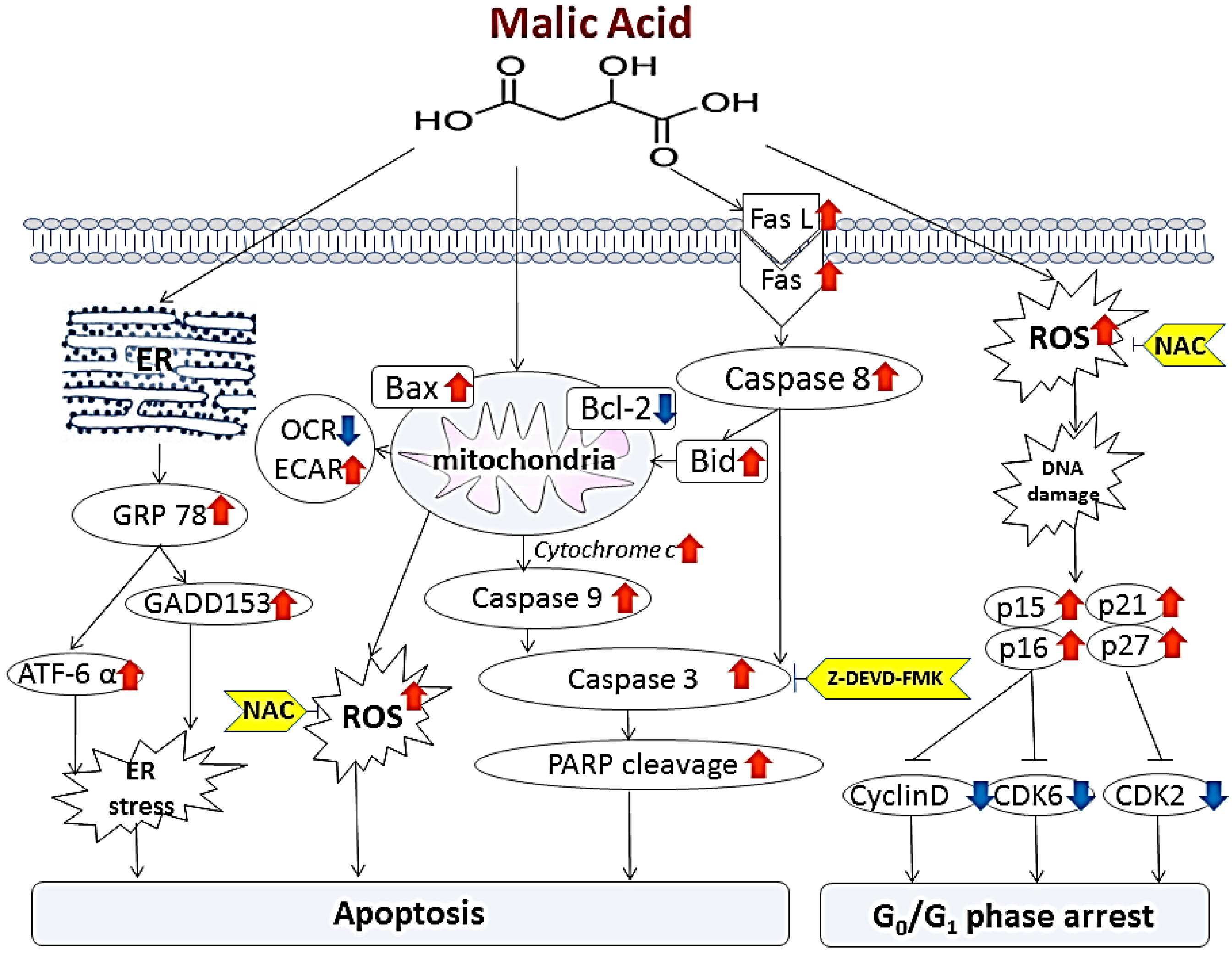
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Human Immortalized Keratinocytes (HaCaT) Cell Line and Normal Human Keratinocytes (NHEKs)
4.3. Morphological Changes and Viability of HaCaT Cells Treated with or without MA
4.4. Flow Cytometric Analysis of Cell Cycle and Apoptosis of HaCaT Cells and NHEKs Treated with Different Concentrations of MA
4.5. Examination of Apoptosis by 4,6-Diamidino-2-Phenylindole Dihydrochloride (DAPI) Staining and Determination of DNA Fragmentation by Gel Electrophoresis
4.6. Determination of Mitochondrial ROS Production and Intracellular ROS by Flow Cytometry
4.7. Measurement of Bioenergetic Parameters of Mitochondria by Seahorse XF24 Analyzer
4.8. Determination the Effect of Caspase-3 Inhibitor Z-DEVD-FMK and ROS Inhibitor NAC on Cell Viability in MA-Treated HaCaT Cells
4.9. Western Blotting for Examining the effect of MA on Expressions of Apoptosis-Related and Cell Cycle Checkpoints-Related Proteins
4.10. Statistical Analysis
5. Conclusions

Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Taylor, M.B.; Yanaki, J.S.; Draper, D.O.; Shurtz, J.C.; Coglianese, M. Successful short-term and long-term treatment of melasma and postinflammatory hyperpigmentation using vitamin C with a full-face iontophoresis mask and a mandelic/malic acid skin care regimen. J. Drugs Dermatol. 2013, 12, 45–50. [Google Scholar] [PubMed]
- Ramos-e-Silva, M.; Celem, L.R.; Ramos-e-Silva, S.; Fucci-da-Costa, A.P. Anti-aging cosmetics: Facts and controversies. Clin. Dermatol. 2013, 31, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Al-Bawab, A.; Friberg, S.E.; Bergamaschi, M.M.; Santos, O.D. Some non-equilibrium phenomena in the malic acid/water/Polysorbate 81 system. Int. J. Pharm. 2007, 332, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Fiume, Z. Final report on the safety assessment of Malic Acid and Sodium Malate. Int. J. Toxicol. 2001, 20, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Kurtzweil, P. Alpha hydroxyl acids for skin care: Smooth sailing or rough seas? FDA Consumer. 1998, 32, 30–35. [Google Scholar] [PubMed]
- Chou, T.C.; Chang, C.H. A case of facial hyperkeratosis induced by a cosmeceutical containing alpha-hydroxy acid and sunlight overexposure successfully treated using oral acitretin. Dermatol. Sinica 2013, 31, 161–162. [Google Scholar] [CrossRef]
- Hsiao, Y.P.; Huang, H.L.; Lai, W.W.; Chung, J.G.; Yang, J.H. Antiproliferative effects of lactic acid via the induction of apoptosis and cell cycle arrest in a human keratinocyte cell line (HaCaT). J. Dermatol. Sci. 2009, 54, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.W.; Hsiao, Y.P.; Chung, J.G.; Wei, Y.H.; Cheng, Y.W.; Yang, J.H. Synergistic phototoxic effects of glycolic acid in a human keratinocyte cell line (HaCaT). J. Dermatol. Sci. 2011, 64, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Green, B.A.; Yu, R.J.; van Scott, E.J. Clinical and cosmeceutical uses of hydroxyacids. Clin. Dermatol. 2009, 27, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Bilaç, C.; Şahin, M.T.; Öztürkcan, S. Chronic actinic damage of facial skin. Clin. Dermatol. 2014, in press. [Google Scholar]
- Usuki, A.; Ohashi, A.; Sato, H.; Ochiai, Y.; Ichihashi, M.; Funasaka, Y. The inhibitory effect of glycolic acid and lactic acid on melanin synthesis in melanoma cells. Exp. Dermatol. 2003, 12, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Tung, R.C.; Bergfeld, W.F.; Vidimos, A.T.; Remzi, B.K. α-Hydroxy Acid–Based Cosmetic Procedures. Guidelines for patient management. Am. J. Clin. Dermatol. 2000, 1, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Kaidbey, K.; Sutherland, B.; Bennett, P.; Wamer, W.G.; Barton, C.; Dennis, D.; Kornhauser, A. Topical glycolic acid enhances photodamage by ultraviolet light. Photodermatol. Photoimmunol. Photomed. 2003, 19, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.T.; Kim, E.J.; Ahn, K.S.; Jung, K.M.; Yun, Y.P.; Park, Y.K.; Lee, S.H. Inhibitory effect of glycolic acid on ultraviolet-induced skin tumorigenesis in SKH-1 hairless mice and its mechanism of action. Mol. Carcinog. 2001, 31, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.S.; Park, K.S.; Jung, K.M.; Jung, H.K.; Lee, S.H.; Chung, S.Y.; Yang, K.H.; Yun, Y.P.; Pyo, H.B.; Park, Y.K.; et al. Inhibitory effect of glycolic acid on ultraviolet B-induced c-fos expression, AP-1 activation and p53-p21 response in a human keratinocyte cell line. Cancer Lett. 2002, 186, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Ying, T.H.; Chen, C.W.; Hsiao, Y.P.; Hung, S.J.; Chung, J.G.; Yang, J.H. Citric acid induces cell-cycle arrest and apoptosis of human immortalized keratinocyte cell line (HaCaT) via caspase- and mitochondrial-dependent signaling pathways. Anticancer Res. 2013, 33, 4411–4420. [Google Scholar] [PubMed]
- Taiwan Ministry of Health and Welfare. Commercially Available Alpha-Hydroxy Acids-Containing Cosmetic Ingredients Have complete Management Practices. Available online: http://cosmetic-recall.fda.gov.tw (accessed on 9 September 2014).
- Yang, J.H.; Chou, C.C.; Cheng, Y.W.; Sheen, L.Y.; Chou, M.C.; Yu, H.S.; Wei, Y.H.; Chung, J.G. Effects of glycolic acid on the induction of apoptosis via caspase-3 activation in human leukemia cell line (HL-60). Food Chem. Toxicol. 2004, 42, 1777–1784. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, Y.P.; Yu, C.S.; Yu, C.C.; Yang, J.S.; Chiang, J.H.; Lu, C.C.; Huang, H.Y.; Tang, N.Y.; Yang, J.H.; Huang, A.C.; et al. Triggering apoptotic death of human malignant melanoma a375.s2 cells by bufalin: involvement of caspase cascade-dependent and independent mitochondrial signaling pathways. Evid. Based Complement. Altern. Med. 2012. [Google Scholar] [CrossRef]
- Raleigh, D.R.; Haas-Kogan, D.A. Molecular targets and mechanisms of radiosensitization using DNA damage response pathways. Future Oncol. 2013, 9, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R.; Amarante-Mendes, G.P. The point-of-no-return: Mitochondria, caspases, and the commitment to cell death. Results Probl. Cell Differ. 1998, 24, 45–61. [Google Scholar] [PubMed]
- Julian, D.; April, K.L.; Patel, S.; Stein, J.R.; Wohlgemuth, S.E. Mitochondrial depolarization following hydrogen sulfide exposure in erythrocytes from a sulfide-tolerant marine invertebrate. J. Exp. Biol. 2005, 208, 4109–4122. [Google Scholar] [CrossRef] [PubMed]
- Yehia, H.N.; Draper, R.K.; Mikoryak, C.; Walker, E.K.; Bajaj, P.; Musselman, I.H.; Daigrepont, M.C.; Dieckmann, G.R.; Pantano, P. Single-walled carbon nanotube interactions with HeLa cells. J. Nanobiotechnol. 2007, 5, 8. [Google Scholar] [CrossRef]
- Ferrick, D.A.; Neilson, A.; Beeson, C. Advances in measuring cellular bioenergetics using extracellular flux. Drug Discov. Today 2008, 13, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.B.; Wei, Y.H. AMPK-mediated increase of glycolysis as an adaptive response to oxidative stress in human cells: Implication of the cell survival in mitochondrial diseases. Biochim. Biophys. Acta 2012, 1822, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, D.G.; Darley-Usmar, V.M.; Wu, M.; Jensen, P.B.; Rogers, G.W.; Ferrick, D.A. Bioenergetic profile experiment using C2C12 myoblast cells. J. Vis. Exp. 2010, 46, 2511. [Google Scholar] [PubMed]
- Hail, N., Jr.; Chen, P.; Kepa, J.J.; Bushman, L.R. Evidence supporting a role for dihydroorotate dehydrogenase, bioenergetics, and p53 in selective teriflunomide-induced apoptosis in transformed versus normal human keratinocytes. Apoptosis 2012, 17, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xie, H.; Chen, X.; Shi, W.; Xiao, X.; Lei, D.; Li, J. Differential response of normal human epidermal keratinocytes and HaCaT cells to hydrogen peroxide-induced oxidative stress. Clin. Exp. Dermatol. 2012, 37, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Fusenig, N.E.; Boukamp, P. Multiple stages and genetic alterations in immortalization, malignant transformation, and tumor progression of human skin keratinocytes. Mol. Carcinog. 1998, 23, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Spörl, F.; Schellenberg, K.; Blatt, T.; Wenck, H.; Wittern, K.P.; Schrader, A.; Kramer, A. A circadian clock in HaCaT keratinocytes. J. Investig. Dermatol. 2011, 131, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Boukamp, P.; Petrussevska, R.T.; Breitkreutz, D.; Hornung, J.; Markham, A.; Fusenig, N.E. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 1988, 106, 761–771. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsiao, Y.-P.; Lai, W.-W.; Wu, S.-B.; Tsai, C.-H.; Tang, S.-C.; Chung, J.-G.; Yang, J.-H. Triggering Apoptotic Death of Human Epidermal Keratinocytes by Malic Acid: Involvement of Endoplasmic Reticulum Stress- and Mitochondria-Dependent Signaling Pathways. Toxins 2015, 7, 81-96. https://doi.org/10.3390/toxins7010081
Hsiao Y-P, Lai W-W, Wu S-B, Tsai C-H, Tang S-C, Chung J-G, Yang J-H. Triggering Apoptotic Death of Human Epidermal Keratinocytes by Malic Acid: Involvement of Endoplasmic Reticulum Stress- and Mitochondria-Dependent Signaling Pathways. Toxins. 2015; 7(1):81-96. https://doi.org/10.3390/toxins7010081
Chicago/Turabian StyleHsiao, Yu-Ping, Wan-Wen Lai, Shi-Bei Wu, Chung-Hung Tsai, Sheau-Chung Tang, Jing-Gung Chung, and Jen-Hung Yang. 2015. "Triggering Apoptotic Death of Human Epidermal Keratinocytes by Malic Acid: Involvement of Endoplasmic Reticulum Stress- and Mitochondria-Dependent Signaling Pathways" Toxins 7, no. 1: 81-96. https://doi.org/10.3390/toxins7010081
APA StyleHsiao, Y. -P., Lai, W. -W., Wu, S. -B., Tsai, C. -H., Tang, S. -C., Chung, J. -G., & Yang, J. -H. (2015). Triggering Apoptotic Death of Human Epidermal Keratinocytes by Malic Acid: Involvement of Endoplasmic Reticulum Stress- and Mitochondria-Dependent Signaling Pathways. Toxins, 7(1), 81-96. https://doi.org/10.3390/toxins7010081