
Chronic Exposure to Deoxynivalenol Has No Influence on the Oral Bioavailability of Fumonisin B1 in Broiler Chickens

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results and Discussion


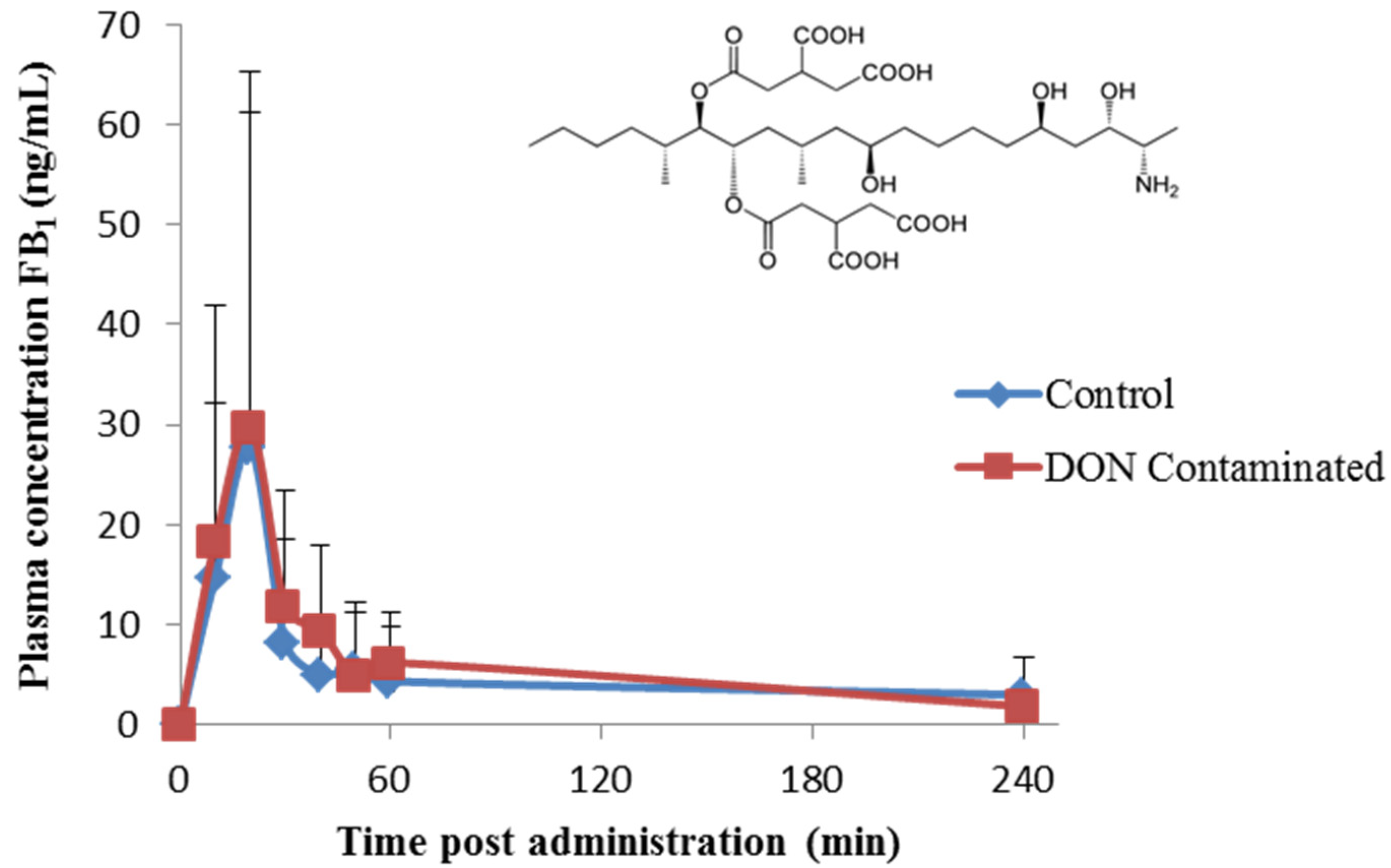{kind=link}
| Toxicokinetic Parameter of FB1 | DON Contaminated | Control |
|---|---|---|
| Cmax (µg/mL) | 0.035 ± 0.0248 | 0.033 ± 0.0213 |
| Tmax (min) | 20 ± 5.0 | 20 ± 5.0 |
| AUC0-t (µg/mL·min) | 83.5 ± 40.21 | 90.6 ± 54.07 |
| kel (min−1) | 0.0075 ± 0.00155 | 0.0078 ± 0.00052 |
| T1/2el (min) | 98.4 ± 22.74 | 106.2 ± 8.34 |
| MRT (min) | 150.8 ± 35.52 | 165.5 ± 48.81 |
| Vd/F (L/kg) | 206.7 ± 92.37 | 234.3 ± 25.03 |
| Cl/F (mL/min·kg) | 1544.3 ± 807.33 | 944.5 ± 387.33 |
| Rel F (%) | 92.2 | 100 |
3. Experimental Section
3.1. Chemicals, Products and Reagents
3.2. Feed Preparation and Experimental Diets
3.3. Animal Experiment
3.4. Quantification of DON, DOM-1 and FB1 in Plasma
3.5. Toxicokinetic and Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Binder, E.M. Managing the risk of mycotoxins in modern feed production. Anim. Feed Sci. Technol. 2007, 133, 149–166. [Google Scholar] [CrossRef]
- Edwards, S.G. Zearalenone risk in european wheat. World Mycotoxin J. 2011, 4, 433–438. [Google Scholar] [CrossRef]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin occurrence in feed and feed raw materials worldwide: Long-term analysis with special focus on Europe and Asia. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O. Current situation of mycotoxin contamination and co-occurrence in animal feed—focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [PubMed]
- Bouhet, S.; Oswald, I.P. The effects of mycotoxins, fungal food contaminants, on the intestinal epithelial cell-derived innate immune response. Vet. Immunol. Immunopathol. 2005, 108, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Bracarense, A.P.F.; Lucioli, J.; Grenier, B.; Drociunas Pacheco, G.; Moll, W.D.; Schatzmayr, G.; Oswald, I.P. Chronic ingestion of deoxynivalenol and fumonisin, alone or in interaction, induces morphological and immunological changes in the intestine of piglets. Br. J. Nutr. 2012, 107, 1776–1786. [Google Scholar] [CrossRef] [PubMed]
- Rotter, B.A. Invited review: Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health A 1996, 48, 1–34. [Google Scholar] [CrossRef]
- Osselaere, A.; Santos, R.; Hautekiet, V.; de Backer, P.; Chiers, K.; Ducatelle, R.; Croubels, S. Deoxynivalenol impairs hepatic and intestinal gene expression of selected oxidative stress, tight junction and inflammation proteins in broiler chickens, but addition of an adsorbing agent shifts the effects to the distal parts of the small intestine. PLoS One 2013, 8, e69014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonissen, G.; van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Haesebrouck, F.; Timbermont, L.; Verlinden, M.; Janssens, G.P.; Eeckhaut, V.; Eeckhout, M.; et al. The mycotoxin deoxynivalenol predisposes for the development of Clostridium perfringens-induced necrotic enteritis in broiler chickens. PLoS One 2014, 9, e108775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yunus, A.W.; Blajet-Kosicka, A.; Kosicki, R.; Khan, M.Z.; Rehman, H.; Bohm, J. Deoxynivalenol as a contaminant of broiler feed: Intestinal development, absorptive functionality, and metabolism of the mycotoxin. Poult. Sci. 2012, 91, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Devreese, M.; Girgis, G.N.; Tran, S.T.; de Baere, S.; de Backer, P.; Croubels, S.; Smith, T.K. The effects of feed-borne Fusarium mycotoxins and glucomannan in turkey poults based on specific and non-specific parameters. Food Chem. Toxicol. 2014, 63, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Nougayrède, J.P.; del Rio, J.C.; Moreno, C.; Marin, D.E.; Ferrier, L.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicol. Appl. Pharm. 2009, 237, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Goossens, J.; Pasmans, F.; Verbrugghe, E.; Vandenbroucke, V.; de Baere, S.; Meyer, E.; Haesebrouck, F.; de Backer, P.; Croubels, S. Porcine intestinal epithelial barrier disruption by the fusarium mycotoxins deoxynivalenol and T-2 toxin promotes transepithelial passage of doxycycline and paromomycin. BMC Vet. Res. 2012, 8, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voss, K.A.; Smith, G.W.; Haschek, W.M. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- Devreese, M.; de Backer, P.; Croubels, S. Overview of the most important mycotoxins for the pig and poultry husbandry. Vlaams Diergen. Tijds. 2013, 82, 171–180. [Google Scholar]
- Riley, R.T.; Enongene, E.; Voss, K.A.; Norred, W.P.; Meredith, F.I.; Sharma, R.P.; Spitsbergen, J.; Williams, D.E.; Carlson, D.B.; Merrill, A.H. Sphingolipid perturbations as mechanisms for fumonisin carcinogenesis. Environ. Health Persp. 2001, 109, 301–308. [Google Scholar] [CrossRef]
- Bouhet, S.; Oswald, I.P. The intestine as a possible target for fumonisin toxicity. Mol. Nutr. Food Res. 2007, 51, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed]
- Javed, T.; Bennett, G.A.; Richard, J.L.; Dombrinkkurtzman, M.A.; Cote, L.M.; Buck, W.B. Mortality in broiler chicks on feed amended with Fusarium proliferatum culture material or with purified fumonisin B1 and moniliformin. Mycopathologia 1993, 123, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Ledoux, D.R.; Brown, T.P.; Weibking, T.S.; Rottinghaus, G.E. Fumonisin toxicity in broiler chicks. J. Vet. Diagn. Investig. 1992, 4, 330–333. [Google Scholar] [CrossRef]
- Weibking, T.S.; Ledoux, D.R.; Bermudez, A.J.; Turk, J.R.; Rottinghaus, G.E.; Wang, E.; Merrill, A.H. Effects of feeding Fusarium moniliforme culture material, containing known levels of fumonisin B1, on the young broiler chick. Poult. Sci. 1993, 72, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, D.; Bailly, J.D.; Benard, G.; Tran, T.S.; Guerre, P. Toxicity of maize containing known levels of fumonisin B1 during force-feeding of ducks. Poult. Sci. 2004, 83, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Kubena, L.F.; Harvey, R.B.; Buckley, S.A.; Bailey, R.H.; Rottinghaus, G.E. Effects of long-term feeding of diets containing moniliformin, supplied by Fusarium fujikuroi culture material, and fumonisin, supplied by Fusarium moniliforme culture material, to laying hens. Poult. Sci. 1999, 78, 1499–1505. [Google Scholar] [CrossRef] [PubMed]
- Tran, S.T.; Auvergne, A.; Benard, G.; Bailly, J.D.; Tardieu, D.; Babile, R.; Guerre, P. Chronic effects of fumonisin B1 on ducks. Poult. Sci. 2005, 84, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Ledoux, D.R.; Bermudez, A.J.; Rottinghaus, G.E. Effects of feeding Fusarium moniliforme culture material, containing known levels of fumonisin B1, in the young turkey poult. Poult. Sci. 1996, 75, 1472–1478. [Google Scholar] [CrossRef] [PubMed]
- Broomhead, J.N.; Ledoux, D.R.; Bermudez, A.J.; Rottinghaus, G.E. Chronic effects of fumonisin B1 in broilers and turkeys fed dietary treatments to market age. Poult. Sci. 2002, 81, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, D.; Bailly, J.D.; Benlashehr, I.; Auby, A.; Jouglar, J.Y.; Guerre, P. Tissue persistence of fumonisin B1 in ducks and after exposure to a diet containing the maximum European tolerance for fumonisins in avian feeds. Chem. Biol. Interact. 2009, 182, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, D.; Bailly, J.D.; Skiba, F.; Grosjean, F.; Guerre, P. Toxicokinetics of fumonisin B1 in turkey poults and tissue persistence after exposure to a diet containing the maximum European tolerance for fumonisins in avian feeds. Food Chem. Toxicol. 2008, 46, 3213–3218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vudathala, D.K.; Prelusky, D.B.; Ayroud, M.; Trenholm, H.L.; Miller, J.D. Pharmacokinetic fate and pathological effects of 14C-fumonisin B1 in laying hens. Nat. Toxins 1994, 2, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Benlashehr, I.; Repussard, C.; Jouglar, J.Y.; Tardieu, D.; Guerre, P. Toxicokinetics of fumonisin B2 in ducks and turkeys. Poult. Sci. 2011, 90, 1671–1675. [Google Scholar] [CrossRef] [PubMed]
- Prelusky, D.B.; Trenholm, H.L.; Savard, M.E. Pharmacokinetic fate of 14C-labelled fumonisin B1 in swine. Nat. Toxins 1994, 2, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Larranaga, M.R.; Anadon, A.; Diaz, M.J.; Fernandez-Cruz, M.L.; Martinez, M.A.; Frejo, M.T.; Martinez, M.; Fernandez, R.; Anton, R.M.; Morales, M.E.; et al. Toxicokinetics and oral bioavailability of fumonisin B1. Vet. Hum. Toxicol. 1999, 41, 357–362. [Google Scholar] [PubMed]
- Shephard, G.S.; Thiel, P.G.; Sydenham, E.W.; Savard, M.E. Fate of a single dose of 14C-labelled fumonisin B1 in vervet monkeys. Nat. Toxins 1995, 3, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Shier, W.T. The fumonisin paradox: A review of research on oral bioavailability of fumonisin B1, a mycotoxin produced by fusarium moniliforme. J. Toxicol. Toxin Rev. 2000, 19, 161–187. [Google Scholar] [CrossRef]
- European Commission. Commission recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, 229, 7–9. [Google Scholar]
- Dänicke, S.; Valenta, H.; Doll, S. On the toxicokinetics and the metabolism of deoxynivalenol (DON) in the pig. Arch. Anim. Nutr. 2004, 58, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Devreese, M.; Antonissen, G.; de Backer, P.; Croubels, S. Efficacy of active carbon towards the absorption of deoxynivalenol in pigs. Toxins 2014, 6, 2998–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maresca, M. From the gut to the brain: Journey and pathophysiological effects of the food-associated trichothecene mycotoxin deoxynivalenol. Toxins 2013, 5, 784–820. [Google Scholar] [CrossRef] [PubMed]
- Mangia, M. Free and Hidden Fumonisins in Maize and Gluten-Free Products. Ph.D. Thesis, University of Parma, Parma, Italy, 2009. [Google Scholar]
- Awad, W.A.; Rehman, H.; Bohm, J.; Razzazi-Fazeli, E.; Zentek, J. Effects of luminal deoxynivalenol and L-proline on electrophysiological parameters in the jejunums of laying hens. Poult. Sci. 2005, 84, 928–932. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M.; Mahfoud, R.; Garmy, N.; Fantini, J. The mycotoxin deoxynivalenol affects nutrient absorption in human intestinal epithelial cells. J. Nutr. 2002, 132, 2723–2731. [Google Scholar] [PubMed]
- Dietrich, B.; Neuenschwander, S.; Bucher, B.; Wenk, C. Fusarium mycotoxin-contaminated wheat containing deoxynivalenol alters the gene expression in the liver and the jejunum of broilers. Animal 2011, 6, 278–291. [Google Scholar] [CrossRef]
- Leslie, J.F.; Plattner, R.D.; Desjardins, A.E.; Klittich, C.J.R. Fumonisin B1 production by strains from different mating populations of Gibberella fujikuroi (Fusarium section Liseola). Phytopathology 1992, 82, 341–345. [Google Scholar] [CrossRef]
- Altpeter, F.; Posselt, U.K. Production of high quantities of 3-acetyldeoxynivalenol and deoxynivalenol. Appl. Microbiol. Biotechnol. 1994, 41, 384–387. [Google Scholar] [CrossRef]
- Hahn, I.; Nagl, V.; Schwartz-Zimmermann, H.E.; Varga, E.; Schwarz, C.; Slavik, V.; Reisinger, N.; Malachová, A.; Cirlini, M.; Generotti, S. Effects of orally administered fumonisin B1 (FB1), partially hydrolysed FB1, hydrolysed FB1 and n-(1-deoxy-d-fructos-1-yl) FB1 on the sphingolipid metabolism in rats. Food Chem. Toxicol. 2015, 76, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Gholamiandehkordi, A.R.; Timbermont, L.; Lanckriet, A.; van Den Broeck, W.; Pedersen, K.; Dewulf, J.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R.; van Immerseel, F. Quantification of gut lesions in a subclinical necrotic enteritis model. Avian Pathol. 2007, 36, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Monbaliu, S.; van Poucke, C.; Detavernier, C.; Dumoulin, F.; van de Velde, M.; Schoeters, E.; van Dyck, S.; Averkieva, O.; van Peteghem, C.; de Saeger, S. Occurrence of mycotoxins in feed as analyzed by a multi-mycotoxin lc-ms/ms method. J. Agric. Food Chem. 2010, 58, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Devreese, M.; de Baere, S.; de Backer, P.; Croubels, S. Quantitative determination of several toxicological important mycotoxins in pig plasma using multi-mycotoxin and analyte-specific high performance liquid chromatography-tandem mass spectrometric methods. J. Chromatogr. A 2012, 1257, 74–80. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antonissen, G.; Devreese, M.; Van Immerseel, F.; De Baere, S.; Hessenberger, S.; Martel, A.; Croubels, S. Chronic Exposure to Deoxynivalenol Has No Influence on the Oral Bioavailability of Fumonisin B1 in Broiler Chickens. Toxins 2015, 7, 560-571. https://doi.org/10.3390/toxins7020560
Antonissen G, Devreese M, Van Immerseel F, De Baere S, Hessenberger S, Martel A, Croubels S. Chronic Exposure to Deoxynivalenol Has No Influence on the Oral Bioavailability of Fumonisin B1 in Broiler Chickens. Toxins. 2015; 7(2):560-571. https://doi.org/10.3390/toxins7020560
Chicago/Turabian StyleAntonissen, Gunther, Mathias Devreese, Filip Van Immerseel, Siegrid De Baere, Sabine Hessenberger, An Martel, and Siska Croubels. 2015. "Chronic Exposure to Deoxynivalenol Has No Influence on the Oral Bioavailability of Fumonisin B1 in Broiler Chickens" Toxins 7, no. 2: 560-571. https://doi.org/10.3390/toxins7020560
APA StyleAntonissen, G., Devreese, M., Van Immerseel, F., De Baere, S., Hessenberger, S., Martel, A., & Croubels, S. (2015). Chronic Exposure to Deoxynivalenol Has No Influence on the Oral Bioavailability of Fumonisin B1 in Broiler Chickens. Toxins, 7(2), 560-571. https://doi.org/10.3390/toxins7020560