
Pokeweed Antiviral Protein: Its Cytotoxicity Mechanism and Applications in Plant Disease Resistance


Abstract
:1. Introduction
2. Different Forms of PAP


3. Cytotoxicity of PAP
3.1. PAP Cytotoxicity in Plants

3.2. PAP Cytotoxicity in Yeast
3.3. Mechanism of PAP Cytotoxicity
3.3.1. N-glycosidase Activity
3.3.2. Interaction with Ribosomal Proteins
3.3.3. Inhibition of Translation
3.3.4. Other Enzymatic Activity
3.3.5. C-Terminal Involvement in PAP Processing
4. Applications of PAP in Plant Disease Resistance
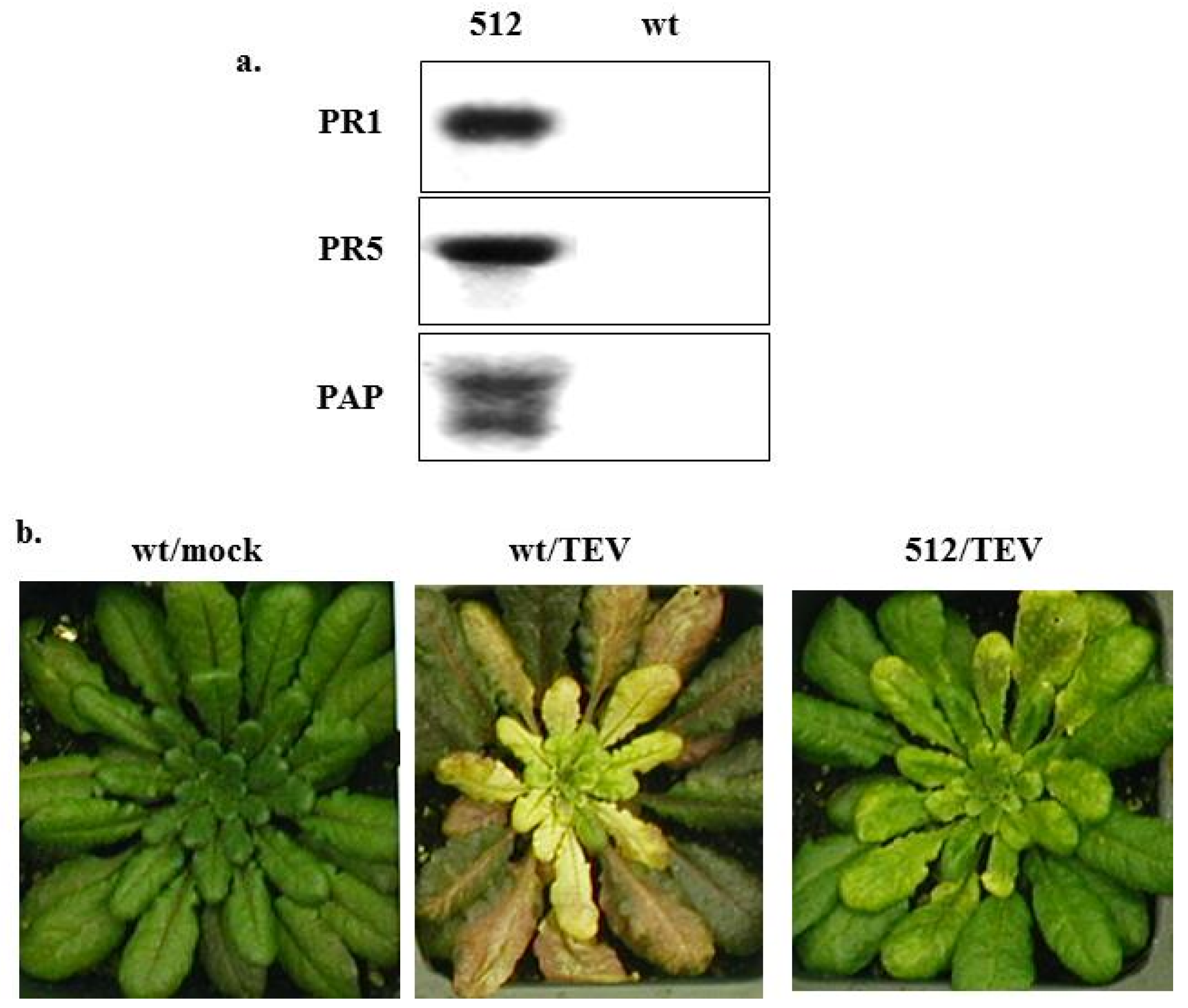
4.1. Against Plant Viruses
4.2. Against Plant Fungi
4.3. Mechanisms of PAP-Induced Disease Resistance


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene description | Fold change |
|---|---|---|
| U49076 | early auxin-induced (IAA20) mRNA | 6.1 |
| AF087819 | auxin transport protein (PIN6) mRNA | 5.7 |
| AF082176 | auxin response factor 9 (ARF9) mRNA | 5.7 |
| L15448 | auxin-responsive protein (IAA1) mRNA | 5.7 |
| AF087819 | auxin transport protein (PIN6) mRNA | 5.3 |
| AL035656 | putative auxin-induced protein | 4.9 |
| AL035656 | small auxin up RNA (SAUR-AC1) | 4.9 |
| AL035656 | auxin-induced protein-like | 4.9 |
| S70188 | small auxin up RNA | 4.9 |
| AJ249794 | lipoxygenase (lox3) | 4.6 |
| AB013301 | AtERF6 ethylene responsive element binding factor | 4.3 |
| AJ012745 | RH27 helicase | 4.3 |
| Z97341 | RNA helicase | 3.7 |
| AJ010475 | DEAD box RNA helicase, RH28 | 3.7 |
| Z97337 | RNA helicase like protein | 3.7 |
| X96600 | pathogenesis-related protein 1 (PR1) | 3.5 |
| U48698 | receptor serine/threonine kinase PR5K (PR5K) | 3.5 |
| U83490 | thaumatin-like protein | 3.2 |
| AL021687 | cytochrome P450 | 3 |
| AL049659 | cytochrome P450-like protein | 3 |
| AL021636 | cytochrome P450-like protein | 3 |
| AL049659 | cytochrome P450-like protein | 3 |
Author Contributions
Conflicts of Interest
References
- Obrig, T.G.; Irvin, J.D.; Hardesty, B. The effect of an antiviral peptide on the ribosomal reactions of the peptide elongation enzymes, EF-I and EF-II. Arch. Biochem. Biophys. 1973, 155, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, S.D.; Shepherd, R.J. Isolation and characterization of a virus inhibitor from Phytolacca americana. Phytopathology 1969, 59, 1787–1794. [Google Scholar] [PubMed]
- Tomlinson, J.A.; Walker, V.M.; Flewett, T.H.; Barclay, G.R. The inhibition of infection by cucumber mosaic virus and influenza virus by extracts from Phytolacca americana. J. Gen. Virol. 1974, 22, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Irvin, J.D. Purification and partial characterization of the antiviral protein from Phytolacca americana which inhibits eukaryotic protein synthesis. Arch. Biochem. Biophys. 1975, 169, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Tsurugi, K. The RNA N-glycosidase activity of ricin A-chain. The characteristics of the enzymatic activity of ricin A-chain with ribosomes and with rRNA. J. Biol. Chem. 1988, 263, 8735–8739. [Google Scholar] [PubMed]
- Olsnes, S.; Kozlov, J.V. Ricin. Toxicon 2001, 39, 1723–1728. [Google Scholar] [CrossRef] [PubMed]
- Di, R.; Kyu, E.; Shete, V.; Saidasan, H.; Kahn, P.C.; Tumer, N.E. Identification of amino acids critical for the cytotoxicity of Shiga toxin 1 and 2 in Saccharomyces cerevisiae. Toxicon 2011, 57, 525–539. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.A.; Morgan, A.E.; Hey, T.D. Characterization and molecular cloning of a proenzyme form of a ribosome-inactivating protein from maize. Novel mechanism of proenzyme activation by proteolytic removal of a 2.8-kilodalton internal peptide segment. J. Biol. Chem. 1991, 266, 23422–23427. [Google Scholar] [PubMed]
- Bass, H.W.; Webster, C.; GR, O.B.; Roberts, J.K.; Boston, R.S. A maize ribosome-inactivating protein is controlled by the transcriptional activator Opaque-2. Plant Cell 1992, 4, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.T.; Ng, Y.M.; Mak, A.N.; Sze, K.H.; Wong, K.B.; Shaw, P.C. Maize ribosome-inactivating protein uses Lys158-lys161 to interact with ribosomal protein P2 and the strength of interaction is correlated to the biological activities. PLoS One 2012, 7, e49608. [Google Scholar] [CrossRef] [PubMed]
- Reinbothe, S.; Reinbothe, C.; Lehmann, J.; Becker, W.; Apel, K.; Parthier, B. JIP60, a methyl jasmonate-induced ribosome-inactivating protein involved in plant stress reactions. Proc. Natl. Acad. Sci. USA 1994, 91, 7012–7016. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, B.; Muller-Uri, F.; Cameron-Mills, V.; Gough, S.; Simpson, D.; Skriver, K.; Mundy, J. The barley 60 kDa jasmonate-induced protein (JIP60) is a novel ribosome-inactivating protein. Plant J. 1994, 6, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Rustgi, S.; Pollmann, S.; Buhr, F.; Springer, A.; Reinbothe, C.; von Wettstein, D.; Reinbothe, S. JIP60-mediated, jasmonate- and senescence-induced molecular switch in translation toward stress and defense protein synthesis. Proc. Natl. Acad. Sci. USA 2014, 111, 14181–14186. [Google Scholar] [CrossRef] [PubMed]
- Tumer, N.E.; Hudak, K.; di, R.; Coetzer, C.; Wang, P.; Zoubenko, O. Pokeweed antiviral protein and its applications. In Plant Biotechnology: New Products and Applications; Hammond, J., McGarvey, P., Yusibov, V., Eds.; Springer: New York, NY, USA, 1999; pp. 139–158. [Google Scholar]
- Parikh, B.A.; Tumer, N.E. Antiviral activity of ribosome inactivating proteins in medicine. Mini Rev. Med. Chem. 2004, 4, 523–543. [Google Scholar] [CrossRef] [PubMed]
- Irvin, J.D.; Uckun, F.M. Pokeweed antiviral protein: Ribosome inactivation and therapeutic applications. Pharmacol. Ther. 1992, 55, 279–302. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, L.; Aron, G.M.; Irvin, J.D.; Stirpe, F. Purification and partial characterization of another form of the antiviral protein from the seeds of Phytolacca americana L. (pokeweed). Biochem. J. 1982, 203, 55–59. [Google Scholar] [PubMed]
- Honjo, E.; Dong, D.; Motoshima, H.; Watanabe, K. Genomic clones encoding two isoforms of pokeweed antiviral protein in seeds (PAP-S1 and S2) and the N-glycosidase activities of their recombinant proteins on ribosomes and DNA in comparison with other isoforms. J. Biochem. 2002, 131, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Chen, Z.C.; Antoniw, J.F.; White, R.F. Isolation and characterization of a cDNA clone encoding the anti-viral protein from Phytolacca americana. Plant Mol. Biol. 1991, 17, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Poyet, J.L.; Radom, J.; Hoeveler, A. Isolation and characterization of a cDNA clone encoding the pokeweed antiviral protein II from Phytolacca americana and its expression in E. coli. FEBS Lett. 1994, 347, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Poyet, J.L.; Hoeveler, A. cDNA cloning and expression of pokeweed antiviral protein from seeds in Escherichia coli and its inhibition of protein synthesis in vitro. FEBS Lett. 1997, 406, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Lodge, J.K.; Kaniewski, W.K.; Tumer, N.E. Broad-spectrum virus resistance in transgenic plants expressing pokeweed antiviral protein. Proc. Natl. Acad. Sci. USA 1993, 90, 7089–7093. [Google Scholar] [CrossRef] [PubMed]
- Ready, M.P.; Brown, D.T.; Robertus, J.D. Extracellular localization of pokeweed antiviral protein. Proc. Natl. Acad. Sci. USA 1986, 83, 5053–5056. [Google Scholar] [CrossRef] [PubMed]
- Tourlakis, M.E.; Karran, R.A.; Desouza, L.; Siu, K.W.; Hudak, K.A. Homodimerization of pokeweed antiviral protein as a mechanism to limit depurination of pokeweed ribosomes. Mol. Plant Pathol. 2010, 11, 757–767. [Google Scholar] [PubMed]
- Choi, D.S.; Kim, N.H.; Hwang, B.K. Pepper mitochondrial FORMATE DEHYDROGENASE1 regulates cell death and defnese responses against bacterial pathogens. Plant Physiol. 2014, 166, 1298–1311. [Google Scholar] [CrossRef] [PubMed]
- Marathe, R.; Anandalakshmi, R.; Liu, Y.; Dinesh-Kumar, S.P. The tobacco mosaic virus resistance gene, N. Mol. Plant Pathol. 2002, 3, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Hur, Y.; Hwang, D.J.; Zoubenko, O.; Coetzer, C.; Uckun, F.M.; Tumer, N.E. Isolation and characterization of pokeweed antiviral protein mutations in Saccharomyces cerevisiae: Identification of residues important for toxicity. Proc. Natl. Acad. Sci. USA 1995, 92, 8448–8452. [Google Scholar] [CrossRef] [PubMed]
- Schlossman, D.; Withers, D.; Welsh, P.; Alexander, A.; Robertus, J.; Frankel, A. Role of glutamic acid 177 of the ricin toxin A chain in enzymatic inactivation of ribosomes. Mol. Cell. Biol. 1989, 9, 5012–5021. [Google Scholar] [PubMed]
- Halling, K.C.; Halling, A.C.; Murray, E.E.; Ladin, B.F.; Houston, L.L.; Weaver, R.F. Genomic cloning and characterization of a ricin gene from Ricinus communis. Nucleic Acids Res. 1985, 13, 8019–8033. [Google Scholar] [CrossRef] [PubMed]
- Frankel, A.; Welsh, P.; Richardson, J.; Robertus, J.D. Role of arginine 180 and glutamic acid 177 of ricin toxin A chain in enzymatic inactivation of ribosomes. Mol. Cell. Biol. 1990, 10, 6257–6263. [Google Scholar] [PubMed]
- Hudak, K.A.; Parikh, B.A.; di, R.; Baricevic, M.; Santana, M.; Seskar, M.; Tumer, N.E. Generation of pokeweed antiviral protein mutations in Saccharomyces cerevisiae: Evidence that ribosome depurination is not sufficient for cytotoxicity. Nucleic Acids Res. 2004, 32, 4244–4256. [Google Scholar] [CrossRef] [PubMed]
- Cakir, B.; Tumer, N.E. Arabidopsis Bax inhibitor-1 inhibits cell death induced by pokeweed antiviral protein in Saccharomyces cerevisae. Microl. Cell 2015, 2, 43–56. [Google Scholar] [CrossRef]
- Endo, Y.; Tsurugi, K. RNA N-glycosidase activity of ricin A-chain. Mechanism of action of the toxic lectin ricin on eukaryotic ribosomes. J. Biol. Chem. 1987, 262, 8128–8130. [Google Scholar] [PubMed]
- Monzingo, A.F.; Robertus, J.D. X-ray analysis of substrate analogs in the ricin A-chain active site. J. Mol. Biol. 1992, 227, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Rajamohan, F.; Mao, C.; Uckun, F.M. Binding interactions between the active center cleft of recombinant pokeweed antiviral protein and the alpha-sarcin/ricin stem loop of ribosomal RNA. J. Biol. Chem. 2001, 276, 24075–24081. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, H.; Fukunaga, Y.; Ueno, R.; Nishimoto, E. Sugar binding effects on the enzymatic reaction and conformation near the active site of pokeweed antiviral protein revealed by fluorescence spectroscopy. J. Fluoresc. 2014, 24, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Hartley, M.R.; Legname, G.; Osborn, R.; Chen, Z.; Lord, J.M. Single-chain ribosome inactivating proteins from plants depurinate Escherichia coli 23S ribosomal RNA. FEBS Lett. 1991, 290, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Monzingo, A.F.; Collins, E.J.; Ernst, S.R.; Irvin, J.D.; Robertus, J.D. The 2.5 A structure of pokeweed antiviral protein. J. Mol. Biol. 1993, 233, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Kurinov, I.V.; Myers, D.E.; Irvin, J.D.; Uckun, F.M. X-ray crystallographic analysis of the structural basis for the interactions of pokeweed antiviral protein with its active site inhibitor and ribosomal RNA substrate analogs. Protein Sci. 1999, 8, 1765–1772. [Google Scholar] [CrossRef] [PubMed]
- Kurinov, I.V.; Rajamohan, F.; Venkatachalam, T.K.; Uckun, F.M. X-ray crystallographic analysis of the structural basis for the interaction of pokeweed antiviral protein with guanine residues of ribosomal RNA. Protein Sci. 1999, 8, 2399–2405. [Google Scholar] [CrossRef] [PubMed]
- Muralikrishna, P.; Alexander, R.W.; Cooperman, B.S. Placement of the alpha-sarcin loop within the 50S subunit: Evidence derived using a photolabile oligodeoxynucleotide probe. Nucleic Acids Res. 1997, 25, 4562–4569. [Google Scholar] [CrossRef] [PubMed]
- Hudak, K.A.; Dinman, J.D.; Tumer, N.E. Pokeweed antiviral protein accesses ribosomes by binding to L3. J. Biol. Chem. 1999, 274, 3859–3864. [Google Scholar] [CrossRef] [PubMed]
- Hampl, H.; Schulze, H.; Nierhaus, K.H. Ribosomal components from Escherichia coli 50 S subunits involved in the reconstitution of peptidyltransferase activity. J. Biol. Chem. 1981, 256, 2284–2288. [Google Scholar] [PubMed]
- Noller, H.F. Peptidyl transferase: Protein, ribonucleoprotein, or RNA? J. Bacteriol. 1993, 175, 5297–5300. [Google Scholar] [PubMed]
- Green, R.; Noller, H.F. Ribosomes and translation. Annu. Rev. Biochem. 1997, 66, 679–716. [Google Scholar] [CrossRef] [PubMed]
- Rajamohan, F.; Ozer, Z.; Mao, C.; Uckun, F.M. Active center cleft residues of pokeweed antiviral protein mediate its high-affinity binding to the ribosomal protein L3. Biochemistry 2001, 40, 9104–9114. [Google Scholar] [CrossRef] [PubMed]
- Di, R.; Tumer, N.E. Expression of a truncated form of ribosomal protein L3 confers resistance to pokeweed antiviral protein and the Fusarium mycotoxin deoxynivalenol. Mol. Plant Microbe Interact. 2005, 18, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Di, R.; Tumer, N.E. An N-terminal fragment of yeast ribosomal protein L3 inhibits the cytotoxicity of pokeweed antiviral protein in Saccharomyces cerevisiae. Toxins (Basel) 2014, 6, 1349–1361. [Google Scholar] [CrossRef]
- Chiou, J.C.; Li, X.P.; Remacha, M.; Ballesta, J.P.; Tumer, N.E. The ribosomal stalk is required for ribosome binding, depurination of the rRNA and cytotoxicity of ricin A chain in Saccharomyces cerevisiae. Mol. Microbiol. 2008, 70, 1441–1452. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.S.; Chu, L.O.; Lee, K.M.; Too, P.H.; Ma, K.W.; Sze, K.H.; Zhu, G.; Shaw, P.C.; Wong, K.B. Interaction between trichosanthin, a ribosome-inactivating protein, and the ribosomal stalk protein P2 by chemical shift perturbation and mutagenesis analyses. Nucleic Acids Res. 2007, 35, 1660–1672. [Google Scholar] [CrossRef] [PubMed]
- Ayub, M.J.; Smulski, C.R.; Ma, K.W.; Levin, M.J.; Shaw, P.C.; Wong, K.B. The C-terminal end of P proteins mediates ribosome inactivation by trichosanthin but does not affect the pokeweed antiviral protein activity. Biochem. Biophys. Res. Commun. 2008, 369, 314–319. [Google Scholar] [CrossRef] [PubMed]
- McCluskey, A.J.; Poon, G.M.; Bolewska-Pedyczak, E.; Srikumar, T.; Jeram, S.M.; Raught, B.; Gariepy, J. The catalytic subunit of shiga-like toxin 1 interacts with ribosomal stalk proteins and is inhibited by their conserved C-terminal domain. J. Mol. Biol. 2008, 378, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Mak, A.N.; Shaw, P.C.; Sze, K.H. Solution structure of an active mutant of maize ribosome-inactivating protein (MOD) and its interaction with the ribosomal stalk protein P2. J. Mol. Biol. 2010, 395, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, Y.; Fujii, K.; Yoshikawa, T.; Kobayashi, Y.; Kondo, T. Pokeweed antiviral protein region Gly209-Lys225 is critical for RNA N-glycosidase activity of the prokaryotic ribosome. Phytochemistry 2008, 69, 1653–1660. [Google Scholar] [CrossRef] [PubMed]
- Lapadula, W.J.; Sanchez-Puerta, M.V.; Ayub, M.J. Convergent evolution led ribosome inactivating proteins to interact with ribosomal stalk. Toxicon 2012, 59, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Sperti, S.; Montanaro, L.; Mattioli, A.; Testoni, G.; Stirpe, F. Inhibition of protein synthesis in vitro by crotins and ricin. Effect on the steps of peptide chain elongation. Biochem. J. 1976, 156, 7–13. [Google Scholar] [PubMed]
- Ramakrishnan, V. Ribosome structure and the mechanism of translation. Cell 2002, 108, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Andersen, G.R.; Nissen, P.; Nyborg, J. Elongation factors in protein biosynthesis. Trends Biochem. Sci. 2003, 28, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S. After the ribosome structure: How does translocation work? RNA 2003, 9, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Brigotti, M.; Rambelli, F.; Zamboni, M.; Montanaro, L.; Sperti, S. Effect of alpha-sarcin and ribosome-inactivating proteins on the interaction of elongation factors with ribosomes. Biochem. J. 1989, 257, 723–727. [Google Scholar] [PubMed]
- Mansouri, S.; Nourollahzadeh, E.; Hudak, K.A. Pokeweed antiviral protein depurinates the sarcin/ricin loop of the rRNA prior to binding of aminoacyl-tRNA to the ribosomal A-site. RNA 2006, 12, 1683–1692. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, L.; Valbonesi, P.; Gorini, P.; Pession, A.; Stirpe, F. Polynucleotide: Adenosine glycosidase activity of saporin-L1: Effect on DNA, RNA and poly(A). Biochem. J. 1996, 319 (Pt 2), 507–513. [Google Scholar] [PubMed]
- Barbieri, L.; Valbonesi, P.; Bonora, E.; Gorini, P.; Bolognesi, A.; Stirpe, F. Polynucleotide: Adenosine glycosidase activity of ribosome-inactivating proteins: Effect on DNA, RNA and poly(A). Nucleic Acids Res. 1997, 25, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Hudak, K.A.; Wang, P.; Tumer, N.E. A novel mechanism for inhibition of translation by pokeweed antiviral protein: Depurination of the capped RNA template. RNA 2000, 6, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Hudak, K.A.; Bauman, J.D.; Tumer, N.E. Pokeweed antiviral protein binds to the cap structure of eukaryotic mRNA and depurinates the mRNA downstream of the cap. RNA 2002, 8, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Hudak, K.A. A novel interaction of pokeweed antiviral protein with translation initiation factors 4G and iso4G: A potential indirect mechanism to access viral RNAs. Nucleic Acids Res. 2006, 34, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, A.E.; Khan, M.A.; Tumer, N.E.; Goss, D.J.; Friedland, D.E. Characterization of pokeweed antiviral protein binding to mRNA cap analogs: Competition with nucleotides and enhancement by translation initiation factor iso4G. Biochim. Biophys. Acta 2009, 1789, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Parikh, B.A.; Coetzer, C.; Tumer, N.E. Pokeweed antiviral protein regulates the stability of its own mRNA by a mechanism that requires depurination but can be separated from depurination of the alpha-sarcin/ricin loop of rRNA. J. Biol. Chem. 2002, 277, 41428–41437. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Tumer, N.E. Pokeweed antiviral protein cleaves double-stranded supercoiled DNA using the same active site required to depurinate rRNA. Nucleic Acids Res. 1999, 27, 1900–1905. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, L.; Valbonesi, P.; Righi, F.; Zuccheri, G.; Monti, F.; Gorini, P.; Samori, B.; Stirpe, F. Polynucleotide: Adenosine glycosidase is the sole activity of ribosome-inactivating proteins on DNA. J. Biochem. 2000, 128, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, K.; Garred, O.; Prydz, K.; Kozlov, J.V.; Hansen, S.H.; van Deurs, B. Retrograde transport of endocytosed Shiga toxin to the endoplasmic reticulum. Nature 1992, 358, 510–512. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.C.; Roberts, L.M.; Romisch, K.; Davey, J.; Wolf, D.H.; Lord, J.M. Ricin A chain utilises the endoplasmic reticulum-associated protein degradation pathway to enter the cytosol of yeast. FEBS Lett. 1999, 459, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Rapak, A.; Falnes, P.O.; Olsnes, S. Retrograde transport of mutant ricin to the endoplasmic reticulum with subsequent translocation to cytosol. Proc. Natl. Acad. Sci. USA 1997, 94, 3783–3788. [Google Scholar] [CrossRef] [PubMed]
- Di Cola, A.; Frigerio, L.; Lord, J.M.; Ceriotti, A.; Roberts, L.M. Ricin A chain without its partner B chain is degraded after retrotranslocation from the endoplasmic reticulum to the cytosol in plant cells. Proc. Natl. Acad. Sci. USA 2001, 98, 14726–14731. [Google Scholar] [CrossRef] [PubMed]
- Baykal, U.; Tumer, N.E. The C-terminus of pokeweed antiviral protein has distinct roles in transport to the cytosol, ribosome depurination and cytotoxicity. Plant J. 2007, 49, 995–1007. [Google Scholar] [CrossRef] [PubMed]
- Parikh, B.A.; Baykal, U.; di, R.; Tumer, N.E. Evidence for retro-translocation of pokeweed antiviral protein from endoplasmic reticulum into cytosol and separation of its activity on ribosomes from its activity on capped RNA. Biochemistry 2005, 44, 2478–2490. [Google Scholar] [CrossRef] [PubMed]
- Duggar, B.M.; Armstrong, J.K. The effect of treatng the virus of tobacco mosaic with the juice of various plants. Ann. Missouri Bot. Gard. 1925, 12, 359–366. [Google Scholar] [CrossRef]
- Gasso, S.; Shepherd, R.J. Isolation and partial characterization of virus inhibitors from plant species taxonomically related to Phytolacca. Phytopathology 1978, 68, 199–205. [Google Scholar] [CrossRef]
- Chen, Z.-C.; Antoniw, J.F.; White, R.F. A possible mechanism for teh antiviral activity of pokeweed antiviral protein. Physiol. Mol. Plant Pathol. 1993, 42, 249–258. [Google Scholar] [CrossRef]
- Tumer, N.E.; Hwang, D.J.; Bonness, M. C-terminal deletion mutant of pokeweed antiviral protein inhibits viral infection but does not depurinate host ribosomes. Proc. Natl. Acad. Sci. USA 1997, 94, 3866–3871. [Google Scholar] [CrossRef] [PubMed]
- Vivanco, J.M.; Tumer, N.E. Translation inhibition of capped and uncapped viral RNAs mediated by ribosome-inactivating proteins. Phytopathology 2003, 93, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Picard, D.; Kao, C.C.; Hudak, K.A. Pokeweed antiviral protein inhibits brome mosaic virus replication in plant cells. J. Biol. Chem. 2005, 280, 20069–20075. [Google Scholar] [CrossRef] [PubMed]
- Karran, R.A.; Hudak, K.A. Depurination within the intergenic region of Brome mosaic virus RNA3 inhibits viral replication in vitro and in vivo. Nucleic Acids Res. 2008, 36, 7230–7239. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.; Massiah, A.; Lomonossoff, G.; Roberts, L.M.; Lord, J.M.; Hartley, M. Correlation between the activities of five ribosome-inactivating proteins in depurination of tobacco ribosomes and inhibition of tobacco mosaic virus infection. Plant J. 1994, 5, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Roberts, W.K.; Selitrennikoff, C.P. Isolation and partial characterization of two antifungal proteins from barley. Biochim. Biophys. Acta 1986, 880, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Logemann, J.; Jach, G.; Tommerup, H.; Mundy, J.; Schell, J. Expression of a barley ribosome-inactivating protein leads to increased fungal protection in transgenic tobacco plants. Nat. Biotechnol. 1992, 10, 305–308. [Google Scholar] [CrossRef]
- Jach, G.; Gornhardt, B.; Mundy, J.; Logemann, J.; Pinsdorf, E.; Leah, R.; Schell, J.; Maas, C. Enhanced quantitative resistance against fungal disease by combinatorial expression of different barley antifungal proteins in transgenic tobacco. Plant J. 1995, 8, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.; Payne, G.A.; Boston, R.S. Maize ribosome-inactivating protein inhibits normal development of Aspergillus nidulans and Aspergillus flavus. Mol. Plant Microbe Interact. 2001, 14, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Zoubenko, O.; Uckun, F.; Hur, Y.; Chet, I.; Tumer, N. Plant resistance to fungal infection induced by nontoxic pokeweed antiviral protein mutants. Nat. Biotechnol. 1997, 15, 992–996. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, S.; Shulaev, V.; Tumer, N.E. Expression of pokeweed antiviral protein in transgenic plants induces virus resistance in grafted wild-type plants independently of salicylic acid accumulation and pathogenesis-related protein synthesis. Plant Physiol. 1997, 114, 1113–1121. [Google Scholar] [PubMed]
- Zoubenko, O.; Hudak, K.; Tumer, N.E. A non-toxic pokeweed antiviral protein mutant inhibits pathogen infection via a novel salicylic acid-independent pathway. Plant Mol. Biol. 2000, 44, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Gozzo, F.; Faoro, F. Systemic acquired resistance (50 years after discovery): Moving from the lab to the field. J. Agric. Food Chem. 2013, 61, 12473–12491. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Reddy, A.S. Cloning and expression of a PR5-like protein from Arabidopsis: Inhibition of fungal growth by bacterially expressed protein. Plant Mol. Biol. 1997, 34, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Schaad, M.C.; Anderberg, R.J.; Carrington, J.C. Strain-specific interaction of the tobacco etch virus NIa protein with the translation initiation factor eIF4E in the yeast two-hybrid system. Virology 2000, 273, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Sewelam, N.; Kazan, K.; Thomas-Hall, S.R.; Kidd, B.N.; Manners, J.M.; Schenk, P.M. Ethylene response factor 6 is a regulator of reactive oxygen species signaling in Arabidopsis. PLoS One 2013, 8, e70289. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zafian, P.; Choudhary, M.; Lawton, M. The PR5K receptor protein kinase from Arabidopsis thaliana is structurally related to a family of plant defense proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 2598–2602. [Google Scholar] [CrossRef] [PubMed]
- Deihimi, T.; Niazi, A.; Ebrahimi, M.; Kajbaf, K.; Fanaee, S.; Bakhtiarizadeh, M.R.; Ebrahimie, E. Finding the undiscovered roles of genes: An approach using mutual ranking of coexpressed genes and promoter architecture-case study: Dual roles of thaumatin like proteins in biotic and abiotic stresses. Springerplus 2012, 1, 30. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di, R.; Tumer, N.E. Pokeweed Antiviral Protein: Its Cytotoxicity Mechanism and Applications in Plant Disease Resistance. Toxins 2015, 7, 755-772. https://doi.org/10.3390/toxins7030755
Di R, Tumer NE. Pokeweed Antiviral Protein: Its Cytotoxicity Mechanism and Applications in Plant Disease Resistance. Toxins. 2015; 7(3):755-772. https://doi.org/10.3390/toxins7030755
Chicago/Turabian StyleDi, Rong, and Nilgun E. Tumer. 2015. "Pokeweed Antiviral Protein: Its Cytotoxicity Mechanism and Applications in Plant Disease Resistance" Toxins 7, no. 3: 755-772. https://doi.org/10.3390/toxins7030755
APA StyleDi, R., & Tumer, N. E. (2015). Pokeweed Antiviral Protein: Its Cytotoxicity Mechanism and Applications in Plant Disease Resistance. Toxins, 7(3), 755-772. https://doi.org/10.3390/toxins7030755