
Cholera Toxin B: One Subunit with Many Pharmaceutical Applications

Abstract
:1. Introduction
1.1. Cholera
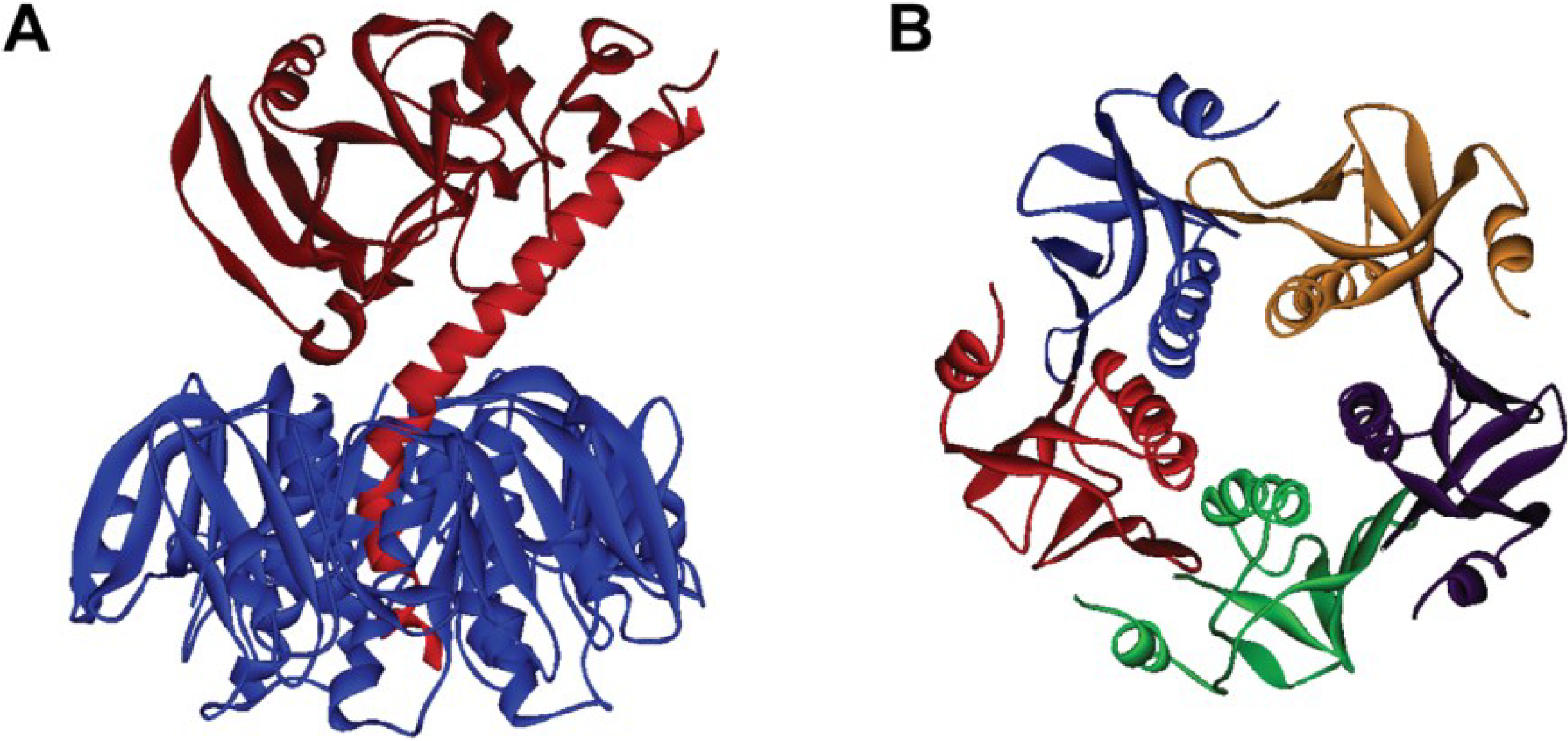
1.2. CT

1.3. Current Vaccines
2. CTB as a Vaccine Adjuvant
2.1. CTB-Based Immunogens against Bacterial Pathogens
2.2. CTB-Based Immunogens against HIV
2.3. Novel CTB-Based Vaccine Delivery and Antigen Conjugation Methods
3. CTB in Inflammation
3.1. CTB’s Anti-Inflammatory Activity in Various Inflammatory Diseases
3.2. Recombinant or Non-Recombinant CTB: Conflicting Results of CTB’s Anti-Inflammatory Activity in in Vitro Experiments
4. rCTB Production Methods


{kind=link}
{kind=link}
| System | Expression Host | Functional Evaluation | Mode of Expression | CTB Yield | Purification | Reference |
|---|---|---|---|---|---|---|
| Bacterial fermentation | V. cholerae | Affinity for GM1-ganglioside confirmed (GM1-ELISA) and immunogenic in mice | Expression plasmid: (pML-LCTBtac2) transformation | 1g/L culture | Affinity chromatography (lyso-GM1 ganglioside Spherosil column) | [102] |
| E. coli | Affinity for GM1-ganglioside confirmed (GM1-ELISA) | Expression plasmid: pQE30 transformation | 9 mg/L culture | IMAC* Purification and membrane-filtration | [103] | |
| Detected by anti-CT antibody (Western Blot) | Expression plasmid: pAE_ctxB transformation | 1.2g/L culture | Centrifugation | [104] | ||
| Affinity for GM1-ganglioside confirmed (GM1-ELISA) | Expression plasmid: pTG8148 transformation | 1 g/L culture | Cation exchange Chromatography (S-Sepharose FF column) | [105] | ||
| Detected by anti-CT antibody (Western Blot) | Expression plasmid: pGEM-T-ctxB transformation | 80 mg/L culture | Centrifugation | [106] | ||
| Lactobacilli | Affinity for GM1-ganglioside confirmed (GM1-ELISA) and immunogenic in mice | Expression plasmid: (pLDH-CTB-His-Term) transformation | 1 mg/L culture | IMAC Purification | [107] | |
| Bacillus brevis | Affinity for GM1-ganglioside confirmed (GM1-ELISA) | Expression plasmid: (pNU212-CTB) transformation | N/A | Affinity chromatography (D-galactose-agarose column) | [108] | |
| Yeast culture | Pichia pastoris | Affinity for GM1-ganglioside confirmed (GM1-ELISA) and immunogenic in mice | Expression plasmid: (pB) transformation | 50 mg/L culture | IMAC Purification | [109] |
| Insect cell culture | B. mori (silkworm larvae) | Affinity for GM1-ganglioside confirmed (GM1-ELISA) and immunogenic in mice | Baculovirus expression system | 54.4 mg/L larval hemolymph | Centrifugation | [110] |
| Plants | Solanum tubersosum (potato) | Affinity for GM1-ganglioside confirmed (GM1-ELISA) | Transgenic (Agrobacterium-mediated transformation. | 0.5% of total soluble protein | Centrifugation | [112] |
| Affinity for GM1-ganglioside confirmed (GM1-ELISA) | Transgenic (Agrobacterium-mediated transformation) | 0.3% of total soluble protein | Non-purified (edible plant vaccine) | [124] | ||
| Daucus carota (carrot) | Affinity for GM1-ganglioside confirmed (GM1-ELISA) | Transgenic (Agrobacterium-mediated transformation) | 0.48% of total soluble protein | Non-purified (edible vaccine) | [113] | |
| Oryza sativa (rice seed) | Affinity for GM1-ganglioside confirmed | Transgenic (Agrobacterium-mediated transformation) | 2.1% of total soluble protein | Non-purified (edible vaccine) | [101] | |
| Detected by anti-CTB antibody (Western Blot) | Transgenic (Agrobacterium-mediated transformation) | 3.37 mg/g rice seeds | IMAC Purification | [114] | ||
| Affinity for GM1-ganglioside confirmed | Transgenic (Agrobacterium-mediated transformation) | 2.35 mg/g of seed | Non-purified (edible vaccine) | [115] | ||
| Affinity for GM1-ganglioside confirmed | Transgenic (Expression plasmid biolistic-mediated transformation) | 2.1% of total seed | Non-purified (edible vaccine) | [116] | ||
| Latuca sativa (lettuce) | Affinity for GM1-ganglioside confirmed (GM1-ELISA) | Transgenic (Agrobacterium-mediated transformation) | 0.24% of total soluble protein | Non-purified (edible vaccine) | [117] | |
| Lycopersicon esculentum (tomato) | Affinity for GM1-ganglioside confirmed (GM1-ELISA) | Transgenic (Agrobacterium-mediated transformation) | 0.04% of total soluble protein | Non-purified (edible vaccine) | [118] | |
| Detected by anti-CTB antibody and immunogenic in mice | Transgenic (Agrobacterium-mediated transformation) | 0.081% of total soluble protein | Non-purified (edible vaccine) | [125] | ||
| Nicotiana benthamiana (a tobacco relative) | Affinity for GM1-ganglioside confirmed (GM1-ELISA) and immunogenic in mice | Transient (plant viral vectors) | 1.5 mg/g leaf material or 49.9% of total soluble protein | IMAC Purification, Hydroxyapatite Chromatography (CHT column) | [100] | |
| Affinity for GM1-ganglioside confirmed (GM1-ELISA) | Transgenic (Agrobacterium-mediated transformation) | 0.56% of total soluble protein | Centrifugation | [112] | ||
| Affinity for GM1-ganglioside confirmed (GM1-ELISA) | Transgenic (Agrobacterium-mediated transformation) | 0.095% of total soluble leaf protein | Immunoaffinity column chromatography (anti-CT IgG resin) | [119] | ||
| Affinity for GM1-ganglioside confirmed (GM1-ELISA) | Transient (plant viral vectors) | 0.14% of total soluble leaf protein | Centrifugation | [120] | ||
| Nicotiana tabacum (tobacco) | Affinity for GM1-ganglioside confirmed (GM1-ELISA) | Transplastomic (Expression plasmid [pLD-LH-CTB] microprojectile bombardment) | 4.1% of total soluble protein | Non-purified crude leaf extract | [121] | |
| Robusta sp. (banana callus) | Detected by anti-CT antibody (Western Blot) | Transgenic (Agrobacterium-mediated transformation) | 125 µg/g callus tissue | Non-purified (edible vaccine) | [122] | |
| Zea mays (maize seed) | Affinity for GM1-ganglioside confirmed and immunogenic in mice | Transgenic (Plasmid microprojectile bombardment) | 1.56 µg/g dry seed weight | Non-purified (edible vaccine) | [123] |
5. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lutz, C.; Erken, M.; Noorian, P.; Sun, S.; McDougald, D. Environmental reservoirs and mechanisms of persistence of Vibrio cholerae. Front. Microbiol. 2013, 4, 375. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Ghosh, K.; Raychoudhuri, A.; Pan, A.; Bhattacharya, M.K.; Mukhopadhyay, A.K.; Ramamurthy, T.; Bhattacharya, S.K.; Nandy, R.K. Phenotypic and genotypic traits and epidemiological implication of Vibrio cholerae O1 and O139 strains in India during 2003. J. Med. Microbiol. 2007, 56, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.B.; LaRocque, R.C.; Qadri, F.; Ryan, E.T.; Calderwood, S.B. Cholera. Lancet 2012, 379, 2466–2476. [Google Scholar] [CrossRef] [PubMed]
- Merrell, D.S.; Butler, S.M.; Qadri, F.; Dolganov, N.A.; Alam, A.; Cohen, M.B.; Calderwood, S.B.; Schoolnik, G.K.; Camilli, A. Host-induced epidemic spread of the cholera bacterium. Nature 2002, 417, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Stine, O.C.; Morris, J.G., Jr. Circulation and transmission of clones of Vibrio cholerae during cholera outbreaks. Curr. Top. Microbiol. Immunol. 2014, 379, 181–193. [Google Scholar] [PubMed]
- Leung, D.T.; Chowdhury, F.; Calderwood, S.B.; Qadri, F.; Ryan, E.T. Immune responses to cholera in children. Expert Rev. Anti Infect. Ther. 2012, 10, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Piarroux, R.; Faucher, B. Cholera epidemics in 2010: Respective roles of environment, strain changes, and human-driven dissemination. Clin. Microbiol. Infect. 2012, 18, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Orata, F.D.; Keim, P.S.; Boucher, Y. The 2010 cholera outbreak in Haiti: How science solved a controversy. PLoS Pathog. 2014, 10, e1003967. [Google Scholar] [CrossRef] [PubMed]
- WHO. Cholera 2012. Wkly. Epidemiol. Rec. 2013, 88, 321–336. [Google Scholar]
- WHO. Cholera vaccines: WHO position paper. Wkly. Epidemiol. Rec. 2010, 85, 117–128. [Google Scholar]
- Siddique, A.K.; Nair, G.B.; Alam, M.; Sack, D.A.; Huq, A.; Nizam, A.; Longini, I.M., Jr.; Qadri, F.; Faruque, S.M.; Colwell, R.R.; et al. El Tor cholera with severe disease: A new threat to Asia and beyond. Epidemiol. Infect. 2010, 138, 347–352. [Google Scholar] [CrossRef]
- Wernick, N.L.; Chinnapen, D.J.; Cho, J.A.; Lencer, W.I. Cholera toxin: An intracellular journey into the cytosol by way of the endoplasmic reticulum. Toxins 2010, 2, 310–325. [Google Scholar] [CrossRef] [PubMed]
- Lencer, W.I.; Tsai, B. The intracellular voyage of cholera toxin: Going retro. Trends Biochem. Sci. 2003, 28, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Chinnapen, D.J.; Chinnapen, H.; Saslowsky, D.; Lencer, W.I. Rafting with cholera toxin: Endocytosis and trafficking from plasma membrane to ER. FEMS Microbiol. Lett. 2007, 266, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.; Holmgren, J. Cholera toxin structure, gene regulation and pathophysiological and immunological aspects. Cell Mol. Life Sci. 2008, 65, 1347–1360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.G.; Scott, D.L.; Westbrook, M.L.; Nance, S.; Spangler, B.D.; Shipley, G.G.; Westbrook, E.M. The three-dimensional crystal structure of cholera toxin. J. Mol. Biol. 1995, 251, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.; Holmgren, J. Cholera toxin—A foe & A friend. Indian J. Med. Res. 2011, 133, 153–163. [Google Scholar] [PubMed]
- Basu, I.; Mukhopadhyay, C. Insights into Binding of Cholera Toxin to GM1 Containing Membrane. Langmuir 2014, 30, 15244–15252. [Google Scholar] [CrossRef] [PubMed]
- Saslowsky, D.E.; te Welscher, Y.M.; Chinnapen, D.J.; Wagner, J.S.; Wan, J.; Kern, E.; Lencer, W.I. Ganglioside GM1-mediated transcytosis of cholera toxin bypasses the retrograde pathway and depends on the structure of the ceramide domain. J. Biol. Chem. 2013, 288, 25804–25809. [Google Scholar] [CrossRef] [PubMed]
- Charles, R.C.; Hilaire, I.J.; Mayo-Smith, L.M.; Teng, J.E.; Jerome, J.G.; Franke, M.F.; Saha, A.; Yu, Y.; Kovac, P.; Calderwood, S.B.; et al. Immunogenicity of a killed bivalent (O1 and O139) whole cell oral cholera vaccine, Shanchol, in Haiti. PLoS Negl. Trop. Dis. 2014, 8, e2828. [Google Scholar] [CrossRef]
- Schaetti, C.; Weiss, M.G.; Ali, S.M.; Chaignat, C.L.; Khatib, A.M.; Reyburn, R.; Duintjer Tebbens, R.J.; Hutubessy, R. Costs of illness due to cholera, costs of immunization and cost-effectiveness of an oral cholera mass vaccination campaign in Zanzibar. PLoS Negl. Trop. Dis. 2012, 6, e1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lajoinie, A.; Henin, E.; Kassai, B.; Terry, D. Solid oral forms availability in children: A cost saving investigation. Br. J. Clin. Pharmacol. 2014, 78, 1080–1089. [Google Scholar] [CrossRef] [PubMed]
- Clemens, J.D.; Sack, D.A.; Harris, J.R.; Chakraborty, J.; Khan, M.R.; Stanton, B.F.; Kay, B.A.; Khan, M.U.; Yunus, M.; Atkinson, W.; et al. Field trial of oral cholera vaccines in Bangladesh. Lancet 1986, 2, 124–127. [Google Scholar] [CrossRef]
- Clemens, J.D.; Sack, D.A.; Harris, J.R.; van Loon, F.; Chakraborty, J.; Ahmed, F.; Rao, M.R.; Khan, M.R.; Yunus, M.; Huda, N.; et al. Field trial of oral cholera vaccines in Bangladesh: Results from three-year follow-up. Lancet 1990, 335, 270–273. [Google Scholar] [CrossRef]
- Alam, M.M.; Leung, D.T.; Akhtar, M.; Nazim, M.; Akter, S.; Uddin, T.; Khanam, F.; Mahbuba, D.A.; Ahmad, S.M.; Bhuiyan, T.R.; et al. Antibody avidity in humoral immune responses in Bangladeshi children and adults following administration of an oral killed cholera vaccine. Clin. Vaccine Immunol. 2013, 20, 1541–1548. [Google Scholar] [CrossRef]
- Lucas, M.E.; Deen, J.L.; von Seidlein, L.; Wang, X.Y.; Ampuero, J.; Puri, M.; Ali, M.; Ansaruzzaman, M.; Amos, J.; Macuamule, A.; et al. Effectiveness of mass oral cholera vaccination in Beira, Mozambique. N. Engl. J. Med. 2005, 352, 757–767. [Google Scholar] [CrossRef]
- Taylor, D.N.; Cardenas, V.; Sanchez, J.L.; Begue, R.E.; Gilman, R.; Bautista, C.; Perez, J.; Puga, R.; Gaillour, A.; Meza, R.; et al. Two-year study of the protective efficacy of the oral whole cell plus recombinant B subunit cholera vaccine in Peru. J. Infect. Dis. 2000, 181, 1667–1673. [Google Scholar] [CrossRef]
- Clemens, J.D.; Jertborn, M.; Sack, D.; Stanton, B.; Holmgren, J.; Khan, M.R.; Huda, S. Effect of neutralization of gastric acid on immune responses to an oral B subunit, killed whole-cell cholera vaccine. J. Infect. Dis. 1986, 154, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Chowdhury, M.I.; Khanam, F.; Bhuiyan, M.S.; Chowdhury, F.; Khan, A.I.; Khan, I.A.; Clemens, J.; Ali, M.; Cravioto, A.; et al. Safety and immunogenicity study of a killed bivalent (O1 and O139) whole-cell oral cholera vaccine Shanchol, in Bangladeshi adults and children as young as 1 year of age. Vaccine 2011, 29, 8285–8292. [Google Scholar] [CrossRef]
- Luquero, F.J.; Grout, L.; Ciglenecki, I.; Sakoba, K.; Traore, B.; Heile, M.; Diallo, A.A.; Itama, C.; Page, A.L.; Quilici, M.L.; et al. Use of Vibrio cholerae vaccine in an outbreak in Guinea. N. Engl. J. Med. 2014, 370, 2111–2120. [Google Scholar] [CrossRef]
- Kang, S.S.; Yang, J.S.; Kim, K.W.; Yun, C.H.; Holmgren, J.; Czerkinsky, C.; Han, S.H. Anti-bacterial and anti-toxic immunity induced by a killed whole-cell-cholera toxin B subunit cholera vaccine is essential for protection against lethal bacterial infection in mouse pulmonary cholera model. Mucosal. Immunol. 2013, 6, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, J.; Adamsson, J.; Anjuere, F.; Clemens, J.; Czerkinsky, C.; Eriksson, K.; Flach, C.F.; George-Chandy, A.; Harandi, A.M.; Lebens, M.; et al. Mucosal adjuvants and anti-infection and anti-immunopathology vaccines based on cholera toxin, cholera toxin B subunit and CpG DNA. Immunol. Lett. 2005, 97, 181–188. [Google Scholar] [CrossRef]
- Williams, N.A.; Hirst, T.R.; Nashar, T.O. Immune modulation by the cholera-like enterotoxins: From adjuvant to therapeutic. Immunol. Today 1999, 20, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Bharati, K.; Ganguly, N.K. Cholera toxin: A paradigm of a multifunctional protein. Indian J. Med. Res. 2011, 133, 179–187. [Google Scholar] [PubMed]
- Holmgren, J.; Lycke, N.; Czerkinsky, C. Cholera toxin and cholera B subunit as oral-mucosal adjuvant and antigen vector systems. Vaccine 1993, 11, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Bowman, C.C.; Selgrade, M.K. Utility of rodent models for evaluating protein allergenicity. Regul. Toxicol. Pharmacol. 2009, 54, S58–S61. [Google Scholar] [CrossRef] [PubMed]
- Oyoshi, M.K.; Oettgen, H.C.; Chatila, T.A.; Geha, R.S.; Bryce, P.J. Food allergy: Insights into etiology, prevention, and treatment provided by murine models. J. Allergy Clin. Immunol. 2014, 133, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Elson, C.O.; Ealding, W. Generalized systemic and mucosal immunity in mice after mucosal stimulation with cholera toxin. J. Immunol. 1984, 132, 2736–2741. [Google Scholar] [PubMed]
- Jackson, R.J.; Fujihashi, K.; Xu-Amano, J.; Kiyono, H.; Elson, C.O.; McGhee, J.R. Optimizing oral vaccines: Induction of systemic and mucosal B-cell and antibody responses to tetanus toxoid by use of cholera toxin as an adjuvant. Infect. Immun. 1993, 61, 4272–4279. [Google Scholar] [PubMed]
- Bourguin, I.; Chardes, T.; Bout, D. Oral immunization with Toxoplasma gondii antigens in association with cholera toxin induces enhanced protective and cell-mediated immunity in C57BL/6 mice. Infect. Immun. 1993, 61, 2082–2088. [Google Scholar] [PubMed]
- Holmgren, J.; Czerkinsky, C.; Lycke, N.; Svennerholm, A.M. Strategies for the induction of immune responses at mucosal surfaces making use of cholera toxin B subunit as immunogen, carrier, and adjuvant. Am. J. Trop. Med. Hyg. 1994, 50, 42–54. [Google Scholar] [PubMed]
- Blanchard, T.G.; Lycke, N.; Czinn, S.J.; Nedrud, J.G. Recombinant cholera toxin B subunit is not an effective mucosal adjuvant for oral immunization of mice against Helicobacter felis. Immunology 1998, 94, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Kubota, E.; Joh, T.; Tanida, S.; Sasaki, M.; Kataoka, H.; Watanabe, K.; Itoh, K.; Oshima, T.; Ogasawara, N.; Togawa, S.; et al. Oral vaccination against Helicobacter pylori with recombinant cholera toxin B-subunit. Helicobacter 2005, 10, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Mutsch, M.; Zhou, W.; Rhodes, P.; Bopp, M.; Chen, R.T.; Linder, T.; Spyr, C.; Steffen, R. Use of the inactivated intranasal influenza vaccine and the risk of Bell’s palsy in Switzerland. N. Engl. J. Med. 2004, 350, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Couch, R.B. Nasal vaccination, Escherichia coli enterotoxin, and Bell’s palsy. N. Engl. J. Med. 2004, 350, 860–861. [Google Scholar] [CrossRef] [PubMed]
- Rath, B.; Linder, T.; Cornblath, D.; Hudson, M.; Fernandopulle, R.; Hartmann, K.; Heininger, U.; Izurieta, H.; Killion, L.; Kokotis, P.; et al. All that palsies is not Bell’s -the need to define Bell’s palsy as an adverse event following immunization. Vaccine 2007, 26, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Liu, K.; Xu, G.; Li, X.; Tu, J.; Tang, F.; Xing, Y.; Xi, T. Prophylactic and therapeutic efficacy of the epitope vaccine CTB-UA against Helicobacter pylori infection in a BALB/c mice model. Appl. Microbiol. Biotechnol. 2012, 95, 1437–1444. [Google Scholar] [CrossRef] [PubMed]
- De Geus, E.D.; van Haarlem, D.A.; Poetri, O.N.; de Wit, J.J.; Vervelde, L. A lack of antibody formation against inactivated influenza virus after aerosol vaccination in presence or absence of adjuvantia. Vet. Immunol. Immunopathol. 2011, 143, 143–147. [Google Scholar]
- Boustanshenas, M.; Bakhshi, B.; Ghorbani, M. Investigation into immunological responses against a native recombinant CTB whole-cell Vibrio cholerae vaccine in a rabbit model. J. Appl. Microbiol. 2013, 114, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Baptista, A.A.; Donato, T.C.; Garcia, K.C.; Goncalves, G.A.; Coppola, M.P.; Okamoto, A.S.; Sequeira, J.L.; Andreatti Filho, R.L. Immune response of broiler chickens immunized orally with the recombinant proteins flagellin and the subunit B of cholera toxin associated with Lactobacillus spp. Poult. Sci. 2014, 93, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yin, R.; Liu, K.; Lv, X.; Li, Y.; Duan, X.; Chu, Y.; Xi, T.; Xing, Y. Immunological features and efficacy of a multi-epitope vaccine CTB-UE against H. pylori in BALB/c mice model. Appl. Microbiol. Biotechnol. 2014, 98, 3495–3507. [Google Scholar] [CrossRef] [PubMed]
- Tinker, J.K.; Yan, J.; Knippel, R.J.; Panayiotou, P.; Cornell, K.A. Immunogenicity of a West Nile virus DIII-cholera toxin A2/B chimera after intranasal delivery. Toxins 2014, 6, 1397–1418. [Google Scholar] [CrossRef] [PubMed]
- Czerkinsky, C.; Sun, J.B.; Lebens, M.; Li, B.L.; Rask, C.; Lindblad, M.; Holmgren, J. Cholera toxin B subunit as transmucosal carrier-delivery and immunomodulating system for induction of antiinfectious and antipathological immunity. Ann. N. Y. Acad. Sci. 1996, 778, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Martell, B.A.; Orson, F.M.; Poling, J.; Mitchell, E.; Rossen, R.D.; Gardner, T.; Kosten, T.R. Cocaine vaccine for the treatment of cocaine dependence in methadone-maintained patients: A randomized, double-blind, placebo-controlled efficacy trial. Arch. Gen. Psychiatry 2009, 66, 1116–1123. [Google Scholar] [CrossRef] [PubMed]
- Orson, F.M.; Rossen, R.D.; Shen, X.; Lopez, A.Y.; Wu, Y.; Kosten, T.R. Spontaneous development of IgM anti-cocaine antibodies in habitual cocaine users: Effect on IgG antibody responses to a cocaine cholera toxin B conjugate vaccine. Am. J. Addict. 2013, 22, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Lebens, M.; Holmgren, J. Mucosal vaccines based on the use of cholera toxin B subunit as immunogen and antigen carrier. Dev. Biol. Stand. 1994, 82, 215–227. [Google Scholar] [PubMed]
- Sun, J.B.; Czerkinsky, C.; Holmgren, J. Mucosally induced immunological tolerance, regulatory T cells and the adjuvant effect by cholera toxin B subunit. Scand. J. Immunol. 2010, 71, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Li, X.; Tang, F.; He, Y.; Xing, Y.; Deng, X.; Xi, T. Immunological features and the ability of inhibitory effects on enzymatic activity of an epitope vaccine composed of cholera toxin B subunit and B cell epitope from Helicobacter pylori urease A subunit. Appl. Microbiol. Biotechnol. 2012, 93, 1937–1945. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Yang, J.; Song, H.; Li, T.; Guo, L.; Xing, Y.; Xi, T. Therapeutic efficacy of the multi-epitope vaccine CTB-UE against Helicobacter pylori infection in a Mongolian gerbil model and its microRNA-155-associated immuno-protective mechanism. Vaccine 2014, 32, 5343–5352. [Google Scholar] [CrossRef] [PubMed]
- Kono, M.; Hotomi, M.; Hollingshead, S.K.; Briles, D.E.; Yamanaka, N. Maternal immunization with pneumococcal surface protein A protects against pneumococcal infections among derived offspring. PLoS One 2011, 6, e27102. [Google Scholar] [CrossRef] [PubMed]
- Matoba, N.; Magerus, A.; Geyer, B.C.; Zhang, Y.; Muralidharan, M.; Alfsen, A.; Arntzen, C.J.; Bomsel, M.; Mor, T.S. A mucosally targeted subunit vaccine candidate eliciting HIV-1 transcytosis-blocking Abs. Proc. Natl. Acad. Sci. USA 2004, 101, 13584–13589. [Google Scholar] [CrossRef] [PubMed]
- Nowroozalizadeh, S.; Jansson, M.; Adamsson, J.; Lindblad, M.; Fenyo, E.M.; Holmgren, J.; Harandi, A.M. Suppression of HIV replication in vitro by CpG and CpG conjugated to the non toxic B subunit of cholera toxin. Curr. HIV Res. 2008, 6, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Maeto, C.; Rodriguez, A.M.; Holgado, M.P.; Falivene, J.; Gherardi, M.M. Novel mucosal DNA-MVA HIV vaccination in which DNA-IL-12 plus cholera toxin B subunit (CTB) cooperates to enhance cellular systemic and mucosal genital tract immunity. PLoS One 2014, 9, e107524. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Liu, Y.; Hsi, J.; Wang, H.; Tao, R.; Shao, Y. Cholera toxin B subunit acts as a potent systemic adjuvant for HIV-1 DNA vaccination intramuscularly in mice. Hum. Vaccin Immunother. 2014, 10, 1274–1283. [Google Scholar] [CrossRef] [PubMed]
- Hervouet, C.; Luci, C.; Cuburu, N.; Cremel, M.; Bekri, S.; Vimeux, L.; Maranon, C.; Czerkinsky, C.; Hosmalin, A.; Anjuere, F. Sublingual immunization with an HIV subunit vaccine induces antibodies and cytotoxic T cells in the mouse female genital tract. Vaccine 2010, 28, 5582–5590. [Google Scholar] [CrossRef] [PubMed]
- Boberg, A.; Gaunitz, S.; Brave, A.; Wahren, B.; Carlin, N. Enhancement of epitope-specific cellular immune responses by immunization with HIV-1 peptides genetically conjugated to the B-subunit of recombinant cholera toxin. Vaccine 2008, 26, 5079–5082. [Google Scholar] [CrossRef] [PubMed]
- Zolla-Pazner, S.; Kong, X.P.; Jiang, X.; Cardozo, T.; Nadas, A.; Cohen, S.; Totrov, M.; Seaman, M.S.; Wang, S.; Lu, S. Cross-clade HIV-1 neutralizing antibodies induced with V3-scaffold protein immunogens following priming with gp120 DNA. J. Virol. 2011, 85, 9887–9898. [Google Scholar] [CrossRef] [PubMed]
- Matoba, N.; Kajiura, H.; Cherni, I.; Doran, J.D.; Bomsel, M.; Fujiyama, K.; Mor, T.S. Biochemical and immunological characterization of the plant-derived candidate human immunodeficiency virus type 1 mucosal vaccine CTB-MPR(649–684). Plant Biotechnol. J. 2009, 7, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Matoba, N.; Griffin, T.A.; Mittman, M.; Doran, J.D.; Hanson, C.V.; Montefiori, D.; Alfsen, A.; Bomsel, M.; Mor, T.S. Transcytosis-blocking Abs elicited by an oligomeric immunogen based on the membrane proximal region of HIV-1 gp41 target non-neutralizing epitopes. Curr. HIV Res. 2008, 6, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Matoba, N.; Geyer, B.C.; Kilbourne, J.; Alfsen, A.; Bomsel, M.; Mor, T.S. Humoral immune responses by prime-boost heterologous route immunizations with CTB-MPR(649–684), a mucosal subunit HIV/AIDS vaccine candidate. Vaccine 2006, 24, 5047–5055. [Google Scholar] [CrossRef] [PubMed]
- Matoba, N.; Shah, N.R.; Mor, T.S. Humoral immunogenicity of an HIV-1 envelope residue 649–684 membrane-proximal region peptide fused to the plague antigen F1-V. Vaccine 2011, 29, 5584–5590. [Google Scholar] [CrossRef] [PubMed]
- Arulanandam, B.P.; Metzger, D.W. Modulation of mucosal and systemic immunity by intranasal interleukin 12 delivery. Vaccine 1999, 17, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Li, C.; Lu, H.; Zhu, Z.; Du, S.; Ye, M.; Tan, L.; Ren, D.; Han, J.; Kan, S.; et al. Immune responses to the oral administration of recombinant Bacillus subtilis expressing multi-epitopes of foot-and-mouth disease virus and a cholera toxin B subunit. J. Virol. Methods 2011, 171, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Qiu, Y.; Chen, J.; Zhang, S.; Xu, B.; Gao, Y. Effective transcutaneous immunization against hepatitis B virus by a combined approach of hydrogel patch formulation and microneedle arrays. Biomed. Microdevices 2013, 15, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Anjuere, F.; George-Chandy, A.; Audant, F.; Rousseau, D.; Holmgren, J.; Czerkinsky, C. Transcutaneous immunization with cholera toxin B subunit adjuvant suppresses IgE antibody responses via selective induction of Th1 immune responses. J. Immunol. 2003, 170, 1586–1592. [Google Scholar] [CrossRef] [PubMed]
- Glenn, G.M.; Scharton-Kersten, T.; Vassell, R.; Matyas, G.R.; Alving, C.R. Transcutaneous immunization with bacterial ADP-ribosylating exotoxins as antigens and adjuvants. Infect. Immun. 1999, 67, 1100–1106. [Google Scholar] [PubMed]
- Maheshwari, C.; Pandey, R.S.; Chaurasiya, A.; Kumar, A.; Selvam, D.T.; Prasad, G.B.; Dixit, V.K. Non-ionic surfactant vesicles mediated transcutaneous immunization against hepatitis B. Int. Immunopharmacol. 2011, 11, 1516–1522. [Google Scholar] [CrossRef] [PubMed]
- Harakuni, T.; Kohama, H.; Tadano, M.; Uechi, G.; Tsuji, N.; Matsumoto, Y.; Miyata, T.; Tsuboi, T.; Oku, H.; Arakawa, T. Mucosal vaccination approach against mosquito-borne Japanese encephalitis virus. Jpn. J. Infect. Dis. 2009, 62, 37–45. [Google Scholar] [PubMed]
- Harokopakis, E.; Hajishengallis, G.; Greenway, T.E.; Russell, M.W.; Michalek, S.M. Mucosal immunogenicity of a recombinant Salmonella typhimurium-cloned heterologous antigen in the absence or presence of coexpressed cholera toxin A2 and B subunits. Infect. Immun. 1997, 65, 1445–1454. [Google Scholar] [PubMed]
- Martin, M.; Hajishengallis, G.; Metzger, D.J.; Michalek, S.M.; Connell, T.D.; Russell, M.W. Recombinant antigen-enterotoxin A2/B chimeric mucosal immunogens differentially enhance antibody responses and B7-dependent costimulation of CD4(+) T cells. Infect. Immun. 2001, 69, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Harokopakis, E.; Childers, N.K.; Michalek, S.M.; Zhang, S.S.; Tomasi, M. Conjugation of cholera toxin or its B subunit to liposomes for targeted delivery of antigens. J. Immunol. Methods 1995, 185, 31–42. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, D.T.; McGee, J.P.; Lindblad, M.; Holmgren, J. Cholera toxin B Subunit (CTB) entrapped in microparticles shows comparable immunogenicity to CTB mixed with whole cholera toxin following oral immunization. Int. J. Pharm. 1995, 119, 251–255. [Google Scholar] [CrossRef]
- Rosli, R.; Nograles, N.; Hanafi, A.; Nor Shamsudin, M.; Abdullah, S. Mucosal genetic immunization through microsphere-based oral carriers. Hum. Vaccin Immunother. 2013, 9, 2222–2227. [Google Scholar] [CrossRef] [PubMed]
- Smits, H.H.; Gloudemans, A.K.; van Nimwegen, M.; Willart, M.A.; Soullie, T.; Muskens, F.; de Jong, E.C.; Boon, L.; Pilette, C.; Johansen, F.E.; et al. Cholera toxin B suppresses allergic inflammation through induction of secretory IgA. Mucosal. Immunol. 2009, 2, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Stanford, M.; Whittall, T.; Bergmeier, L.A.; Lindblad, M.; Lundin, S.; Shinnick, T.; Mizushima, Y.; Holmgren, J.; Lehner, T. Oral tolerization with peptide 336–351 linked to cholera toxin B subunit in preventing relapses of uveitis in Behcet’s disease. Clin. Exp. Immunol. 2004, 137, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Nathan, D.M.; Abraham, K.; Brunzell, J.D.; Fradkin, J.E.; Haffner, S.M.; Hsueh, W.; Rewers, M.; Roberts, B.T.; Savage, P.J.; et al. Report of the National Heart, Lung, and Blood Institute-National Institute of Diabetes and Digestive and Kidney Diseases Working Group on Cardiovascular Complications of Type 1 Diabetes Mellitus. Circulation 2005, 111, 3489–3493. [Google Scholar] [CrossRef] [PubMed]
- Odumosu, O.; Nicholas, D.; Payne, K.; Langridge, W. Cholera toxin B subunit linked to glutamic acid decarboxylase suppresses dendritic cell maturation and function. Vaccine 2011, 29, 8451–8458. [Google Scholar] [CrossRef] [PubMed]
- Denes, B.; Fodor, I.; Langridge, W.H. Persistent suppression of type 1 diabetes by a multicomponent vaccine containing a cholera toxin B subunit-autoantigen fusion protein and complete Freund’s adjuvant. Clin. Dev. Immunol. 2013, 2013, 578786. [Google Scholar] [CrossRef] [PubMed]
- Shil, P.K.; Kwon, K.C.; Zhu, P.; Verma, A.; Daniell, H.; Li, Q. Oral delivery of ACE2/Ang-(1–7) bioencapsulated in plant cells protects against experimental uveitis and autoimmune uveoretinitis. Mol. Ther. 2014, 22, 2069–2082. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.; Li, J.; Jin, L.; Liu, J.; Li, T. Nasal immunization with heat shock protein 65 attenuates atherosclerosis and reduces serum lipids in cholesterol-fed wild-type rabbits probably through different mechanisms. Immunol. Lett. 2009, 125, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, R.; Lebens, M.; Hermansson, A.; Fredrikson, G.N.; Strodthoff, D.; Rudling, M.; Ketelhuth, D.F.; Gerdes, N.; Holmgren, J.; Nilsson, J.; et al. Intranasal immunization with an apolipoprotein B-100 fusion protein induces antigen-specific regulatory T cells and reduces atherosclerosis. Arterioscler Thromb. Vasc. Biol. 2010, 30, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Gonzalez, J.A.; Rosales-Mendoza, S.; Romero-Maldonado, A.; Monreal-Escalante, E.; Uresti-Rivera, E.E.; Banuelos-Hernandez, B. Production of a Plant-Derived Immunogenic Protein Targeting ApoB100 and CETP: Toward a Plant-Based Atherosclerosis Vaccine. Mol. Biotechnol. 2014, 56, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Weiner, H.L. The mucosal milieu creates tolerogenic dendritic cells and T(R)1 and T(H)3 regulatory cells. Nat. Immunol. 2001, 2, 671–672. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, H.J.; Rutitzky, L.I.; Lebens, M.; Holmgren, J.; Stadecker, M.J. Diminished immunopathology in Schistosoma mansoni infection following intranasal administration of cholera toxin B-immunodominant peptide conjugate correlates with enhanced transforming growth factor-beta production by CD4 T cells. Parasite Immunol. 2002, 24, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Song, B.; Huang, C.; Xiao, Y.; Fang, M.; Feng, J.; Wang, P.; Zhang, G. Prolonged survival time of allografts by the oral administration of RDP58 linked to the cholera toxin B subunit. Transpl. Immunol. 2012, 27, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Boirivant, M.; Fuss, I.J.; Ferroni, L.; de Pascale, M.; Strober, W. Oral administration of recombinant cholera toxin subunit B inhibits IL-12-mediated murine experimental (trinitrobenzene sulfonic acid) colitis. J. Immunol. 2001, 166, 3522–3532. [Google Scholar] [CrossRef] [PubMed]
- Coccia, E.M.; Remoli, M.E.; Di Giacinto, C.; Del Zotto, B.; Giacomini, E.; Monteleone, G.; Boirivant, M. Cholera toxin subunit B inhibits IL-12 and IFN-γ production and signaling in experimental colitis and Crohn’s disease. Gut 2005, 54, 1558–1564. [Google Scholar] [CrossRef] [PubMed]
- Stal, P.; Befrits, R.; Ronnblom, A.; Danielsson, A.; Suhr, O.; Stahlberg, D.; Brinkberg Lapidus, A.; Lofberg, R. Clinical trial: The safety and short-term efficacy of recombinant cholera toxin B subunit in the treatment of active Crohn’s disease. Aliment. Pharmacol. Ther. 2010, 31, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Tamura, S.; Yamanaka, A.; Shimohara, M.; Tomita, T.; Komase, K.; Tsuda, Y.; Suzuki, Y.; Nagamine, T.; Kawahara, K.; Danbara, H.; et al. Synergistic action of cholera toxin B subunit (and Escherichia coli heat-labile toxin B subunit) and a trace amount of cholera whole toxin as an adjuvant for nasal influenza vaccine. Vaccine 1994, 12, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Hamorsky, K.T.; Kouokam, J.C.; Bennett, L.J.; Baldauf, K.J.; Kajiura, H.; Fujiyama, K.; Matoba, N. Rapid and scalable plant-based production of a cholera toxin B subunit variant to aid in mass vaccination against cholera outbreaks. PLoS Negl. Trop. Dis. 2013, 7, e2046. [Google Scholar] [CrossRef] [PubMed]
- Nochi, T.; Takagi, H.; Yuki, Y.; Yang, L.; Masumura, T.; Mejima, M.; Nakanishi, U.; Matsumura, A.; Uozumi, A.; Hiroi, T.; et al. Rice-based mucosal vaccine as a global strategy for cold-chain- and needle-free vaccination. Proc. Natl. Acad. Sci. USA 2007, 104, 10986–10991. [Google Scholar] [CrossRef] [PubMed]
- Lebens, M.; Johansson, S.; Osek, J.; Lindblad, M.; Holmgren, J. Large-scale production of Vibrio cholerae toxin B subunit for use in oral vaccines. Biotechnology 1993, 11, 1574–1578. [Google Scholar] [CrossRef] [PubMed]
- Dakterzada, F.; Mobarez, A.M.; Roudkenar, M.H.; Forouzandeh, M. Production of Pentameric Cholera Toxin B Subunit in Escherichia coli. Avicenna J. Med. Biotechnol. 2012, 4, 89–94. [Google Scholar] [PubMed]
- Boustanshenas, M.; Bakhshi, B.; Ghorbani, M.; Norouzian, D. Comparison of two recombinant systems for expression of cholera toxin B subunit from Vibrio cholerae. Indian J. Med. Microbiol. 2013, 31, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Slos, P.; Speck, D.; Accart, N.; Kolbe, H.V.; Schubnel, D.; Bouchon, B.; Bischoff, R.; Kieny, M.P. Recombinant cholera toxin B subunit in Escherichia coli: High-level secretion, purification, and characterization. Protein Exp. Purif. 1994, 5, 518–526. [Google Scholar] [CrossRef]
- Bakhshi, B.; Boustanshenas, M.; Ghorbani, M. A single point mutation within the coding sequence of cholera toxin B subunit will increase its expression yield. Iran Biomed. J. 2014, 18, 130–135. [Google Scholar] [PubMed]
- Okuno, T.; Kashige, N.; Satho, T.; Irie, K.; Hiramatsu, Y.; Sharmin, T.; Fukumitsu, Y.; Uyeda, S.; Yamada, S.; Harakuni, T.; et al. Expression and secretion of cholera toxin B subunit in Lactobacilli. Biol. Pharm. Bull. 2013, 36, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, Y.; Matano, K.; Asai, T.; Tochikubo, K. Affinity purification of recombinant cholera toxin B subunit oligomer expressed in Bacillus brevis for potential human use as a mucosal adjuvant. FEMS Immunol. Med. Microbiol. 1998, 20, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Miyata, T.; Harakuni, T.; Taira, T.; Matsuzaki, G.; Arakawa, T. Merozoite surface protein-1 of Plasmodium yoelii fused via an oligosaccharide moiety of cholera toxin B subunit glycoprotein expressed in yeast induced protective immunity against lethal malaria infection in mice. Vaccine 2012, 30, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.H.; Jin, H.Q.; Jin, Y.F.; Zhang, Y.Z. Expression of cholera toxin B subunit and assembly as functional oligomers in silkworm. J. Biochem. Mol. Biol. 2005, 38, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, T.; Chong, D.K.; Langridge, W.H. Efficacy of a food plant-based oral cholera toxin B subunit vaccine. Nat. Biotechnol. 1998, 16, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Mikschofsky, H.; König, P.; Keil, G.M.; Hammer, M.; Schirrmeier, H.; Broer, I. Cholera toxin B (CTB) is functional as an adjuvant for cytoplasmatic proteins if directed to the endoplasmatic reticulum (ER), but not to the cytoplasm of plants. Plant Sci. 2009, 177, 35–42. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, M.Y.; Kim, T.G.; Yang, M.S. Expression and assembly of cholera toxin B subunit (CTB) in transgenic carrot (Daucus carota L.). Mol. Biotechnol. 2009, 41, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Kajiura, H.; Wasai, M.; Kasahara, S.; Takaiwa, F.; Fujiyama, K. N-glycosylation and N-glycan moieties of CTB expressed in rice seeds. Mol. Biotechnol. 2013, 54, 784–794. [Google Scholar] [CrossRef] [PubMed]
- Yuki, Y.; Mejima, M.; Kurokawa, S.; Hiroiwa, T.; Takahashi, Y.; Tokuhara, D.; Nochi, T.; Katakai, Y.; Kuroda, M.; Takeyama, N.; et al. Induction of toxin-specific neutralizing immunity by molecularly uniform rice-based oral cholera toxin B subunit vaccine without plant-associated sugar modification. Plant Biotechnol. J. 2013, 11, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Oszvald, M.; Kang, T.J.; Tomoskozi, S.; Jenes, B.; Kim, T.G.; Cha, Y.S.; Tamas, L.; Yang, M.S. Expression of cholera toxin B subunit in transgenic rice endosperm. Mol. Biotechnol. 2008, 40, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, B.G.; Kim, T.G.; Kang, T.J.; Yang, M.S. Expression of a cholera toxin B subunit in transgenic lettuce (Lactuca sativa L.) using Agrobacterium-mediated transformation system. Plant Cell Tiss. Organ. Cult. 2006, 87, 203–210. [Google Scholar] [CrossRef]
- Jani, D.; Meena, L.S.; Rizwan-ul-Haq, Q.M.; Singh, Y.; Sharma, A.K.; Tyagi, A.K. Expression of cholera toxin B subunit in transgenic tomato plants. Transgenic Res. 2002, 11, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.G.; Zhang, G.H.; Liu, C.X.; Zhang, Y.H.; Xiao, C.Z.; Fang, R.X. Purified cholera toxin B subunit from transgenic tobacco plants possesses authentic antigenicity. Biotechnol. Bioeng. 2001, 72, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Rattanapisit, K.; Bhoo, S.H.; Hahn, T.R.; Mason, H.S.; Phoolcharoen, W. Rapid transient expression of cholera toxin B subunit (CTB) in Nicotiana benthamiana. In Vitro Cell Dev. Biol. Plant 2013, 49, 107–113. [Google Scholar] [CrossRef]
- Daniell, H.; Lee, S.B.; Panchal, T.; Wiebe, P.O. Expression of the native cholera toxin B subunit gene and assembly as functional oligomers in transgenic tobacco chloroplasts. J. Mol. Biol. 2001, 311, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Renuga, R.S.; Babu Thandapani, A.; Arumugam, K.R. Expression of Cholera toxin B subunit in Banana callus culture. J. Pharm. Sci. Res. 2010, 2, 26–33. [Google Scholar]
- Karaman, S.; Cunnick, J.; Wang, K. Expression of the cholera toxin B subunit (CT-B) in maize seeds and a combined mucosal treatment against cholera and traveler’s diarrhea. Plant Cell Rep. 2012, 31, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, T.; Chong, D.K.; Merritt, J.L.; Langridge, W.H. Expression of cholera toxin B subunit oligomers in transgenic potato plants. Transgenic Res. 1997, 6, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.L.; He, Z.M.; Peng, Z.Q.; Qi, Y.; Chen, Q.; Yu, S.Y. Cholera toxin B protein in transgenic tomato fruit induces systemic immune response in mice. Transgenic Res. 2007, 16, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Nochi, T.; Yuki, Y.; Katakai, Y.; Shibata, H.; Tokuhara, D.; Mejima, M.; Kurokawa, S.; Takahashi, Y.; Nakanishi, U.; Ono, F.; et al. A rice-based oral cholera vaccine induces macaque-specific systemic neutralizing antibodies but does not influence pre-existing intestinal immunity. J. Immunol. 2009, 183, 6538–6544. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldauf, K.J.; Royal, J.M.; Hamorsky, K.T.; Matoba, N. Cholera Toxin B: One Subunit with Many Pharmaceutical Applications. Toxins 2015, 7, 974-996. https://doi.org/10.3390/toxins7030974
Baldauf KJ, Royal JM, Hamorsky KT, Matoba N. Cholera Toxin B: One Subunit with Many Pharmaceutical Applications. Toxins. 2015; 7(3):974-996. https://doi.org/10.3390/toxins7030974
Chicago/Turabian StyleBaldauf, Keegan J., Joshua M. Royal, Krystal Teasley Hamorsky, and Nobuyuki Matoba. 2015. "Cholera Toxin B: One Subunit with Many Pharmaceutical Applications" Toxins 7, no. 3: 974-996. https://doi.org/10.3390/toxins7030974
APA StyleBaldauf, K. J., Royal, J. M., Hamorsky, K. T., & Matoba, N. (2015). Cholera Toxin B: One Subunit with Many Pharmaceutical Applications. Toxins, 7(3), 974-996. https://doi.org/10.3390/toxins7030974