
Prevention of Aflatoxin B1-Induced DNA Breaks by β-D-Glucan


 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Antigenotoxic Effect of β-d-Glucan

2.2. Degrees of Damage Assessed by the Comet Assay

{kind=link}
{kind=link}
{kind=link}
| Agent/Dose | Time (h) | Grades of Damage (%) | |||
|---|---|---|---|---|---|
| G0 | G1 | G2 | G3 | ||
| Corn oil 0.02 mL | 4 | 88 | 7 | 5 | 1 |
| 10 | 91 | 7 | 1 | 1 | |
| 16 | 92 | 4 | 3 | 1 | |
| DMSO 0.02 mL | 4 | 89 | 7 | 3 | 1 |
| 10 | 86 | 11 | 2 | 1 | |
| 16 | 92 | 5 | 3 | 0 | |
| Glu 700 mg/kg | 4 | 90 | 5 | 4 | 1 |
| 10 | 91 | 5 | 4 | 0 | |
| 16 | 93 | 3 | 3 | 1 | |
| AFB1 1.0 mg/kg | 4 | 69 | 11 | 10 | 10 |
| 10 | 7 | 6 | 26 | 61 | |
| 16 | 18 | 16 | 28 | 38 | |
| Glu + AFB1 100 + 1.0 mg/kg | 4 | 69 | 15 | 8 | 8 |
| 10 | 11 | 16 | 24 | 49 | |
| 16 | 30 | 14 | 23 | 33 | |
| Glu + AFB1 400 + 1.0 mg/kg | 4 | 76 | 9 | 6 | 9 |
| 10 | 21 | 19 | 21 | 39 | |
| 16 | 33 | 18 | 23 | 26 | |
| Glu + AFB1 700 + 1.0 mg/kg | 4 | 80 | 7 | 8 | 5 |
| 10 | 19 | 22 | 27 | 32 | |
| 16 | 26 | 32 | 20 | 22 | |
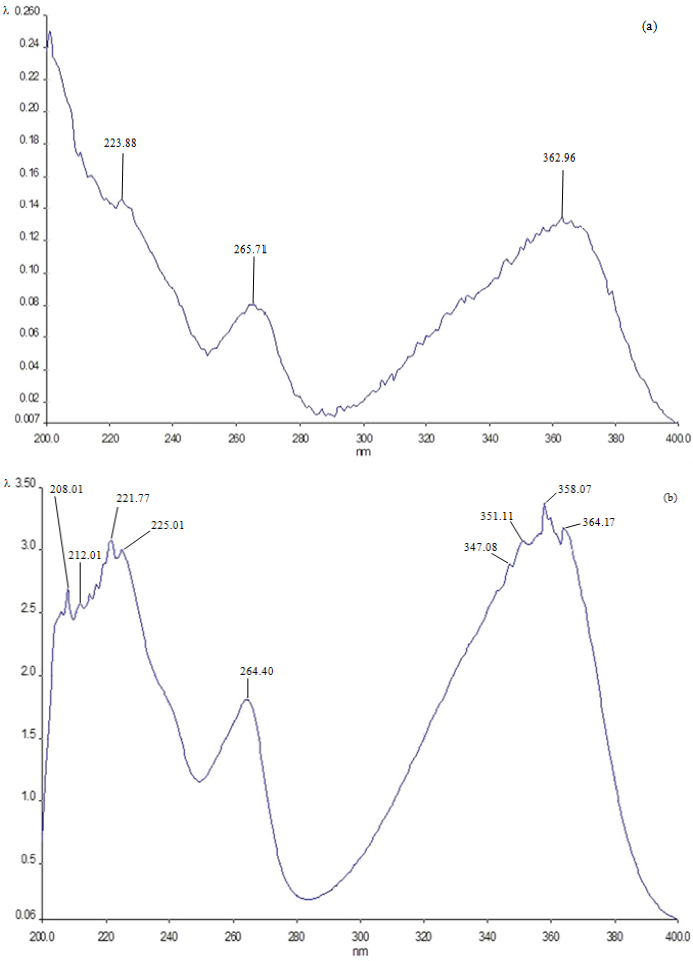
2.3. Melting Points of the Crystals Formed by the Compounds
| Chemical | Melting Point (°C) |
|---|---|
| AFB1/Glu | 180.5 |
| AFB1 | 132.5 |
| Glu | 240.0 |

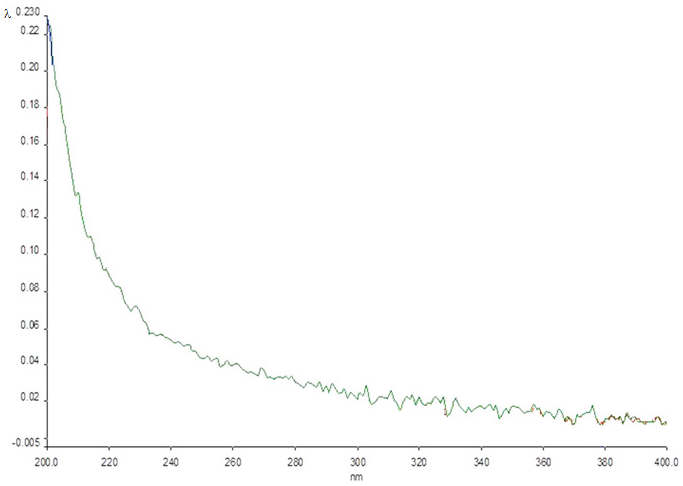
2.4. UV Spectrum Analysis of the Crystals Formed by the Compounds

3. Discussion
4. Experimental Section
4.1. Chemicals and Animals
4.2. Genotoxicity/Antigenotoxicity Protocol
Experimental Design
4.3. Unicellular Alkaline Electrophoresis (Comet) Assay
4.4. Crystallization, Melting Point Determination and Spectroscopic Analysis
5. Conclusions
Author Contributions
Conflicts of Interest
References
- IARC Monographs on the evaluation of carcinogenic risks to humans. Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. Int. Agency Res. Cancer Lyon 1993, 56, 245–397.
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human aflatoxicosis in developing countries: A review of toxicology, exposure, potential health consequences, and interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [PubMed]
- Madrigal-Santillán, E.; Morales-González, J.A.; Vargas-Mendoza, N.; Reyes-Ramírez, P.; Cruz-Jaime, S.; Sumaya-Martínez, T.; Pérez-Pastén, R.; Madrigal-Bujaidar, E. Antigenotoxic studies of different substances to reduce the DNA damage induced by aflatoxin B(1) and ochratoxin A. Toxins 2010, 2, 738–757. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, T.E.; Dowd, P.E.; Desjardins, A.E.; Bhatnagar, D.; Coty, P.J. United States Department of Agriculture-Agricultural Research Service. Research on Preharvest Prevention of Mycotoxins and Mycotoxigenic Fungi in US Crops. Pest. Manag. Sci. 2003, 59, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Caldas, E.D.; Silva, S.C.; Oliveira, J.N. Aflatoxins and ochratoxin A in food and risks to human health. Rev. Saude. Publ. 2002, 36, 319–323. [Google Scholar] [CrossRef]
- Tam, J.; Mankotia, M.; Mably, M.; Pantazopoulos, P.; Neil, R.J.; Calway, P.; Scott, P.M. Survey of breakfast and infant cereals for aflatoxins B1, B2, G1 and G2. Food Addit. Contam. 2006, 23, 693–639. [Google Scholar] [CrossRef] [PubMed]
- Witt, K.L.; Knapton, A.; Wehr, C.M.; Hook, G.J.; Mirsalis, J.; Shelby, M.D.; MacGregor, J.T. Micronucleated Erythrocyte Frequency in Peripheral Blood of B6C3F(1) Mice from Short-Term, Prechronic, and Chronic Studies of the NTP Carcinogenesis Bioassay Program. Environ. Mol. Mutagen. 2000, 36, 163–194. [Google Scholar] [CrossRef]
- Woo, L.L.; Egner, P.A.; Belanger, C.L.; Wattanawaraporn, R.; Trudel, L.J.; Croy, R.G.; Groopman, J.D.; Essigmann, J.M.; Wogan, G.N.; Bouhenguel, J.T. Aflatoxin B1-DNA adduct formation and mutagenicity in livers of neonatal male and female B6C3F1 mice. Toxicol. Sci. 2011, 122, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Kabak, B.; Dobson, A.D.; Var, I. Strategies to prevent mycotoxin contamination of food and animal feed: A review. Crit. Rev. Food Sci. Nutr. 2006, 46, 593–619. [Google Scholar] [CrossRef] [PubMed]
- Robb, J. Mycotoxins: Contamination and Decontamination. Feed Mix. 1993, 1, 18–23. [Google Scholar] [CrossRef]
- Madrigal-Santillan, E.; Madrigal-Bujaidar, E.; Marquez-Marquez, R.; Reyes, A. Antigenotoxic Effect of Saccharomyces cerevisiae on the Damage Produced in Mice Fed with Aflatoxin B1 Contaminated Corn. Food Chem. Toxicol. 2006, 44, 2058–2063. [Google Scholar] [CrossRef] [PubMed]
- Kogan, G.; Pajtinka, M.; Babincova, M.; Miadoková, E.; Rauko, P.; Slamenova, D.; Korolenko, T.A. Yeast Cell Wall Polysaccharides as Antioxidants and Antimutagens: Can They Fight Cancer? Neoplasma 2008, 55, 387–393. [Google Scholar] [PubMed]
- Madrigal-Santillán, E.; Álvarez-González, I.; Márquez-Márquez, R.; Velázquez-Guadarrama, N.; Madrigal-Bujaidar, E. Inhibitory Effect of Mannan on the Toxicity Produced in Mice Fed Aflatoxin B1 Contaminated Corn. Arch. Environ. Contam. Toxicol. 2007, 53, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Angeli, J.P.; Ribeiro, L.R.; Gonzaga, M.L.; Soares Sde, A.; Ricardo, M.P.; Tsuboy, M.S.; Stidl, R.; Knasmueller, S.; Linhares, R.E.; Mantovani, M.S. Protective effects of beta-glucan extracted from Agaricus brasiliensis against chemically induced DNA damage in human lymphocytes. Cell Biol. Toxicol. 2006, 22, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Brendler-Schwaab, S.; Hartmann, A.; Pfuhler, S.; Speit, G. The in vivo comet assay: Use and status in genotoxicity testing. Mutagenesis 2005, 20, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Pfohl-Leszkowicz, A. Formation, persistence and significance of DNA adduct formation in relation to some pollutants from a board perspective. Adv. Toxicol. 2008, 2, 183–240. [Google Scholar]
- Dusinska, M.; Collins, A.R. The comet assay in human biomonitoring: Gene-Environment interactions. Mutagenesis 2008, 23, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, A.; Bajpayee, M.; Parmar, D. Comet assay: A reliable tool for the assessment of DNA damage in different models. Cell Biol. Toxicol. 2009, 25, 5–32. [Google Scholar] [CrossRef] [PubMed]
- Bajpayee, M.; Kumar, A.; Dhawan, A. The comet assay: Assessment of in vitro and in vivo DNA damage. Methods Mol. Biol. 2013, 1044, 325–345. [Google Scholar] [PubMed]
- Corcuera, L.A.; Vettorazzi, A.; Arbillaga, L.; Pérez, N.; Gil, A.G.; Azqueta, A.; González-Peñas, E.; García-Jalón, J.A.; López de Cerain, A. Genotoxicity of Aflatoxin B1 and Ochratoxin A after simultaneous application of the in vivo micronucleus and comet assay. Food Chem. Toxicol. 2015, 76, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Monroe, D.H.; Eaton, D.L. Effects of modulation of hepatic glutathione on biotransformation and covalent binding of aflatoxin B1 to DNA in the mouse. Toxicol. Appl. Pharmacol. 1988, 94, 118–127. [Google Scholar] [CrossRef]
- Tice, R.R. The single cell gel comet assay. A microgel electrophoretic technique for the detection of DNA damage and repair in individual cells. In Environmental Mutagenesis; Phillips, D.H., Venitt, S., Eds.; Bios Scientific Publishers: Oxford, UK, 1995; pp. 315–339. [Google Scholar]
- Ezz El-Arab, A.M.; Girgis, S.M.; Hegazy, E.M.; Abd El-Khalek, A.B. Effect of dietary honey on intestinal microflora and toxicity of mycotoxins in mice. BMC Complement. Altern. Med. 2006, 14, 6. [Google Scholar] [CrossRef] [PubMed]
- Madrigal-Santillán, E.; Morales-González, J.A.; Sánchez-Gutiérrez, M.; Reyes-Arellano, A.; Madrigal-Bujaidar, E. Investigation on the protective effect of α-mannan against the DNA damage induced by aflatoxin B1 in mouse hepatocytes. Int. J. Mol. Sci. 2009, 10, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Adedara, I.A.; Owumi, S.E.; Uwaifo, A.O.; Farombi, E.O. Aflatoxin B1 and ethanol co-exposure induces hepatic oxidative damage in mice. Toxicol. Ind. Health 2010, 26, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.C.; Chung, W.T.; Kwon, J.K.; Yu, J.Y.; Jang, Y.S.; Park, S.M.; Lee, S.Y.; Lee, J.C. Inhibitory effects of quercetin on aflatoxin B1-induced hepatic damage in mice. Food Chem. Toxicol. 2010, 48, 2747–2753. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Heflich, R.H.; Moore, M.M.; Mei, N. Differential mutagenicity of aflatoxin B1 in the liver of neonatal and adult mice. Environ. Mol. Mutagen. 2010, 51, 156–163. [Google Scholar] [PubMed]
- Brahmi, D.; Bouaziz, C.; Ayed, Y.; Ben Mansour, H.; Zourgui, L.; Bacha, H. Chemopreventive effect of cactus Opuntia ficus indica on oxidative stress and genotoxicity of aflatoxin B1. Nutr. Metab. 2011, 18, 73. [Google Scholar] [CrossRef] [PubMed]
- Miadoková, E.; Svidová, S.; Ulčová, V.; Dǜhová, V.; Nad’ová, S.; Rauko, P.; Kogan, G. Diverse biomodulatory effects of glucomannan from Candida utilis. Toxicol. Vitro 2006, 20, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V. Effects of β-glucan on some environmental toxins: An overview. Biomed. Pap. Med. Fac. Univ. Palacky. Olomouc Czech. Repub. 2014, 158, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.N.; Nan, F.H.; Chen, S.; Wu, J.F.; Lu, C.L.; Soni, M.G. Safety assessment of mushroom β-glucan: Subchronic toxicity in rodents and mutagenicity studies. Food Chem. Toxicol. 2011, 49, 2890–2898. [Google Scholar] [CrossRef] [PubMed]
- Angeli, J.P.; Ribeiro, L.R.; Bellini, M.F.; Mantovani, M.S. Beta-glucan extracted from the medicinal mushroom Agaricus blazei prevents the genotoxic effects of benzo[a]pyrene in the human hepatoma cell line HepG2. Arch. Toxicol. 2009, 83, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Križková, L.; Ďuracková, Z.; Šandula, J.; Sasinková, V.; Krajčovič, J. Antioxidative and Antimutagenic Activity of Yeast Cell Wall Mannans in Vitro. Mutat. Res. 2001, 497, 213–222. [Google Scholar] [CrossRef]
- Tohamy, A.A.; El Ghor, A.A.; El Nahas, S.M.; Noshy, M.M. β-Glucan Inhibits the Genotoxicity of Cyclophosphamide, Adriamycin and Cisplatin. Mutat. Res. 2003, 541, 45–53. [Google Scholar] [CrossRef]
- Toklu, H.S.; Öser Seirli, A.; Velioğlu-Öğunc, A.; Centinel, S.; Sener, G. Acetaminophen-Induced Toxicity is Prevented by β-d-Glucan Treatment in Mice. Eur. J. Pharmacol. 2006, 543, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Babincova, M.; Bacova, Z.; Machova, E.; Kogan, G. Antioxidant Properties of Carboxymethyl Glucan: Comparative Analysis. J. Med. Food. 2002, 5, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Križkova, L.; Žitňanová, I.; Mislovičová, D.; Masárová, J.; Sasinková, V.; Ďuračková, Z.; Krajčovič, J. Antioxidant and Antimutagenic Activity of Mannan Neoglycoconjugates: Mannan-Human Serum Albumine and Mannan-Penicillin G Acylase. Mutat. Res. 2006, 606, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Toklu, H.Z.; Şener, G.; Jahovic, N.; Uslu, B.; Arbak, S.; Yeğen, B. B-Glucan Protects Against Burn-Induced Oxidative Organ Damage in Rats. Intern. Immunopharmacol. 2006, 6, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Weisburger, J.H. Antimutagens, Anticarcinogens, and Effective Worldwide Cancer Prevention. J. Environ. Pathol. Toxicol. Oncol. 1999, 18, 85–93. [Google Scholar] [PubMed]
- Weisburger, J.H. Lifestyle, Health and disease prevention: The Underlying Mechanisms. Eur. J. Cancer Prev. 2002, 11 (Suppl. 2), 51–57. [Google Scholar]
- Rao, C.V.; Chou, D.; Simi, B.; Ku, H.; Reddy, B.S. Prevention of Colonic Aberrant Crypt Foci and Modulation of Large Bowel Microbial Activity by Dietary Coffee Fiber, Inulin and Pectin. Carcinogenesis 1998, 19, 1815–1819. [Google Scholar] [CrossRef] [PubMed]
- Glei, M.; Hofman, T.; Küster, K.; Hollmann, J.; Linhahuer, M.G.; Pool-Zobel, B.L. Both Weath (Triticum. aestivum) Bran Arabinoxylans and Gut Flora-Mediated Fermentation Products Protect Human Colon Cells from Genotoxic Activities of 4-Hydroxynonenal and Hydrogen Peroxide. J. Agric. Food Chem. 2006, 54, 2088–2095. [Google Scholar] [CrossRef] [PubMed]
- Pereyra, C.M.; Cavaglieri, L.R.; Chiacciera, S.M.; Dalcero, A. The corn influence on the adsorption levels of aflatoxin B1, and zearalenone by yeast cell wall. J. Appl. Microbiol. 2012, 114, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Freimund, S.; Sauter, M.; Rys, P. Efficient adsorption of the mycotoxins zearalenone and T-2 toxin on a modified yeast glucan. J. Environ. Sci. Health B 2003, B38, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Steed, J.W.; Turner, D.R.; Wallace, K.J. Core Concepts in Supramolecular Chemistry and Nanochemistry; John Wiley & Sons, Ltd.: Chichester, UK, 2007; pp. 5–25. [Google Scholar]
- Reyes-Arellano, A.; Rose, R.; Steller, I.; Sustmann, R. Crystal engineering: Supramolecular Structures by Cocrystallization of meso-1,2-diphenyl-1,2-ethanediol and bisimines. Simple Cases of Moleular Recognition. J. Struct. Chem. 1995, 6, 391–396. [Google Scholar] [CrossRef]
- Reyes, A.; Nájera, J.; Mendoza, R.; Molins, I.; Grau, E.; Espinosa, E.; Salgado, H.; Tamariz, J.; Soriano, M.; Paneerselvam, K. Bisimines in Supramolecular Synthesis. Supramol. Chem. 1999, 11, 17–24. [Google Scholar] [CrossRef]
- Murthy, T.N.; Reddy, B.N.; Devegowda, G. Evaluation of glucomannan for its adsorbing ability of Aflatoxin B1 and T-2 Toxin in the Gastrointestinal Tract of Broiler Chickens. Mycotoxin Res. 2002, 18, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Gallaher, C.M.; Munion, J.; Hesslink, R., Jr.; Wise, J.; Gallaher, D.D. Cholesterol reduction by glucomannan and chitosan is mediated by changes in cholesterol absorption and bile acid and fat excretion in rats. J. Nutr. 2000, 130, 2753–2759. [Google Scholar] [PubMed]
- De Flora, S. Mechanisms of inhibitors of mutagenesis and carcinogenesis. Mutat. Res. 1998, 402, 151–158. [Google Scholar] [CrossRef]
- Machová, E.; Kogan, G.; Alföldi, J.; Sotés, L.; Sándula, J. Enzymic and ultrasonic depolymerization of carboxymethylated β-1,3-D-glucans derived from Saccharomyces cerevisiae. J. Appl. Polym. Sci. 1995, 55, 699–704. [Google Scholar] [CrossRef]
- Hartmann, A.; Agurell, E.; Beevers, C.; Brendler-Schaab, S.; Burlinson, B.; Clay, P.; Collins, A.; Smith, A.; Speit, G.; Thybaud, V.; et al. Recommendations for Conducting the in Vivo Alkaline Comet Assay. Mutagenesis 2003, 18, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Cadena, J.; Tenorio-Vieyra, L.E.; Quintana-Carabia, A.I.; Garcia-Fabila, M.M.; Ramírez-San Juan, E.; Madrigal-Bujaidar, E. Determination of DNA Damage in Floriculturists Exposed to Mixtures of Pesticides. J. Biomed. Biotechnol. 2006, 2006, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bergfors, T.M. Protein Crystallization Technique. Strategies and Tips: A Laboratory Manual; International University Line: La Jolla, CA, USA, 2001. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madrigal-Bujaidar, E.; Morales-González, J.A.; Sánchez-Gutiérrez, M.; Izquierdo-Vega, J.A.; Reyes-Arellano, A.; Álvarez-González, I.; Pérez-Pasten, R.; Madrigal-Santillán, E. Prevention of Aflatoxin B1-Induced DNA Breaks by β-D-Glucan. Toxins 2015, 7, 2145-2158. https://doi.org/10.3390/toxins7062145
Madrigal-Bujaidar E, Morales-González JA, Sánchez-Gutiérrez M, Izquierdo-Vega JA, Reyes-Arellano A, Álvarez-González I, Pérez-Pasten R, Madrigal-Santillán E. Prevention of Aflatoxin B1-Induced DNA Breaks by β-D-Glucan. Toxins. 2015; 7(6):2145-2158. https://doi.org/10.3390/toxins7062145
Chicago/Turabian StyleMadrigal-Bujaidar, Eduardo, José Antonio Morales-González, Manuel Sánchez-Gutiérrez, Jeannett A. Izquierdo-Vega, Alicia Reyes-Arellano, Isela Álvarez-González, Ricardo Pérez-Pasten, and Eduardo Madrigal-Santillán. 2015. "Prevention of Aflatoxin B1-Induced DNA Breaks by β-D-Glucan" Toxins 7, no. 6: 2145-2158. https://doi.org/10.3390/toxins7062145
APA StyleMadrigal-Bujaidar, E., Morales-González, J. A., Sánchez-Gutiérrez, M., Izquierdo-Vega, J. A., Reyes-Arellano, A., Álvarez-González, I., Pérez-Pasten, R., & Madrigal-Santillán, E. (2015). Prevention of Aflatoxin B1-Induced DNA Breaks by β-D-Glucan. Toxins, 7(6), 2145-2158. https://doi.org/10.3390/toxins7062145