
Partial Characterization of Venom from the Colombian Spider Phoneutria Boliviensis (Aranae:Ctenidae)

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Venom Physicochemical Properties

{kind=link}
{kind=link}
| Phoneutria boliviensis | ||
|---|---|---|
| Male | Female | |
| Wet weight ‡ | 9.7 ± 4.36 mg | 20.9 ± 7.68 mg |
| Dry weigh ‡ | 1.4 ± 0.92 mg | 3.7 ± 1.65 mg |
| Aqueous content * | 89% ± 0.03% | 85% ± 0.02% |
| CF Measures ‡ | L: 12.03 ± 0.69 mm W: 9.6 ± 0.29 mm | L: 14.36 ± 0.39 mm W: 10.65 ± 0.21 mm |
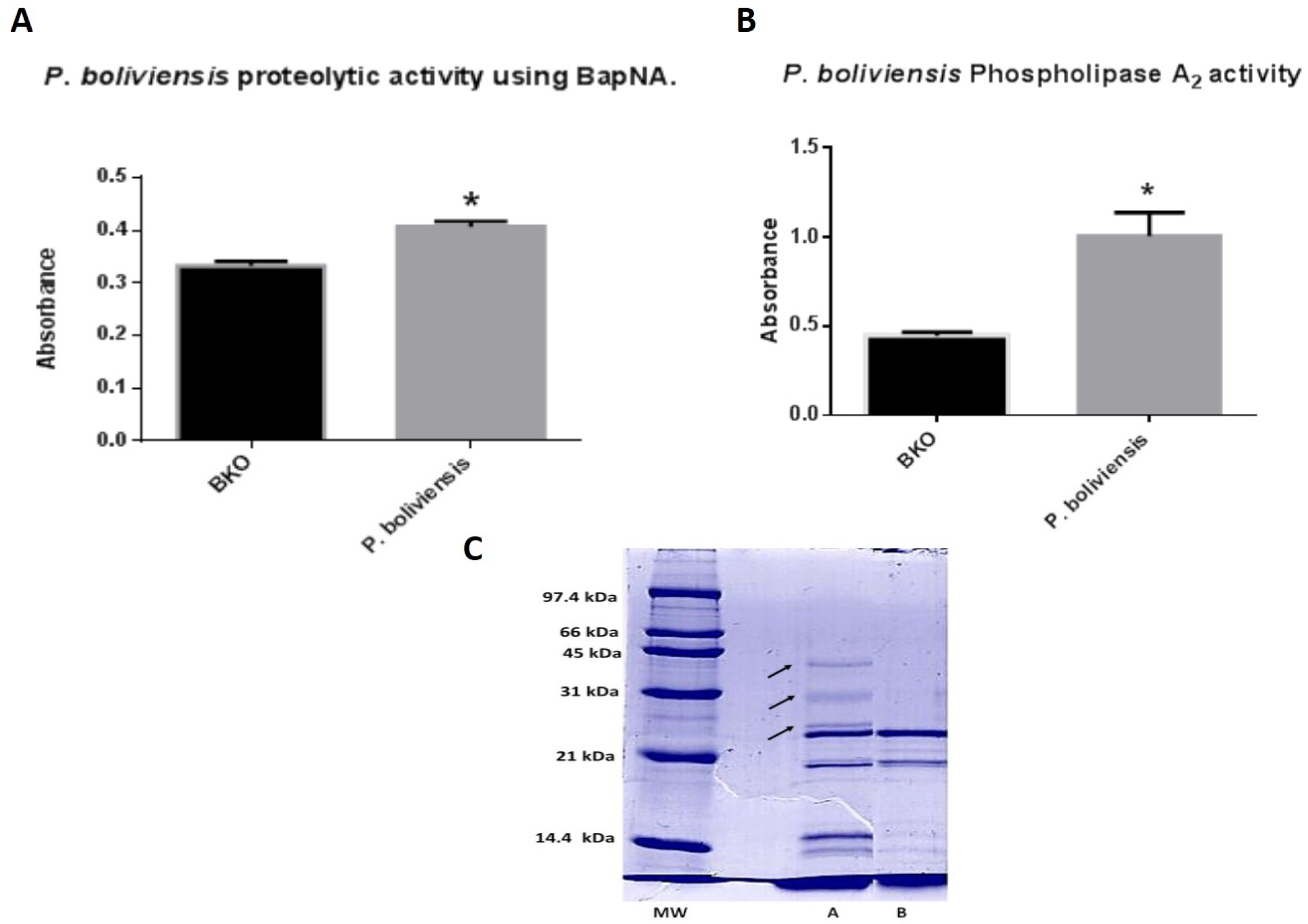
2.2. Enzymatic Properties

2.3. SDS-PAGE
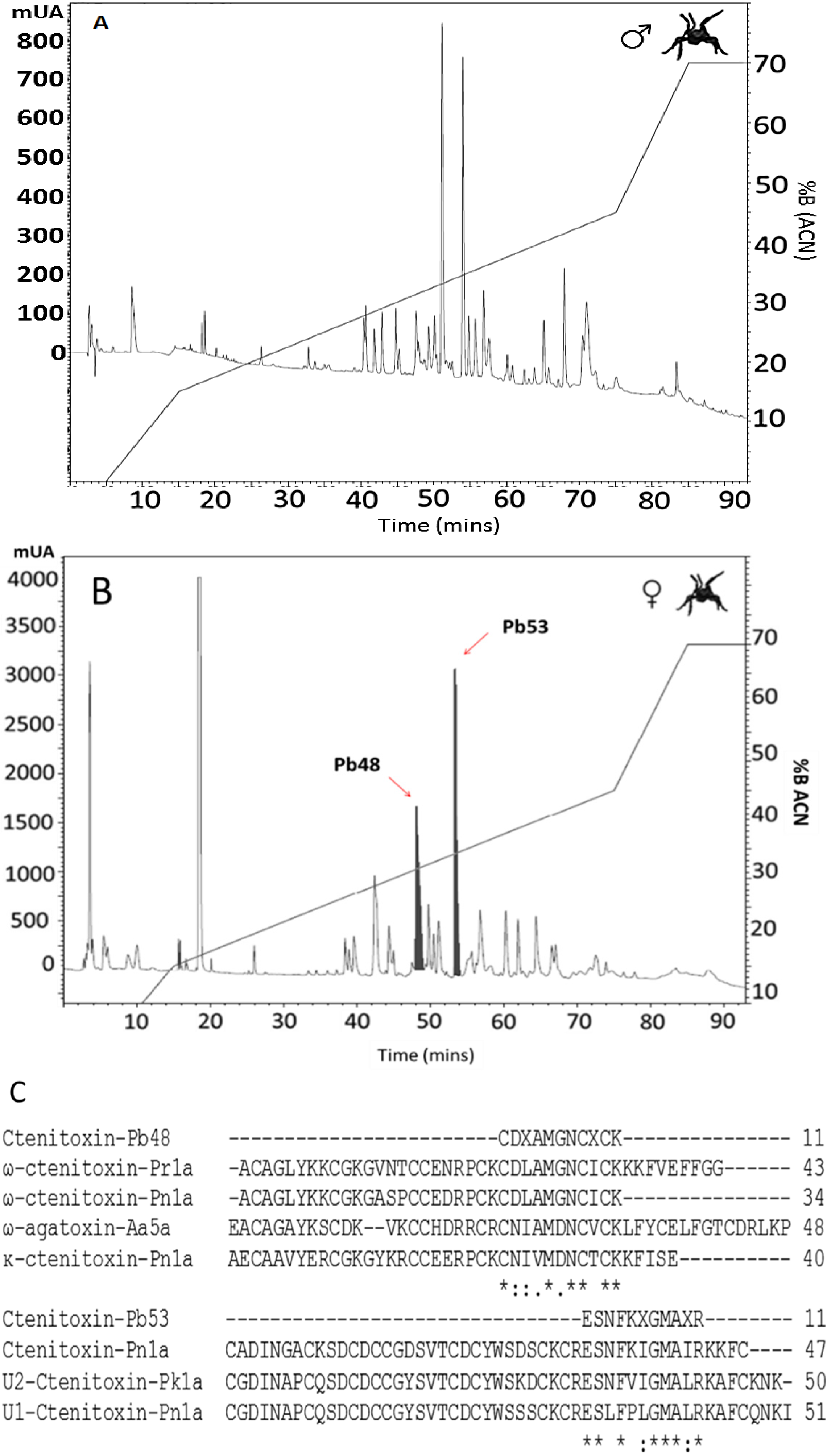
2.4. Reverse-Phase Chromatography

2.5. LC–MS Analysis
| rT (min) | Isotopic Mass (Da) | Average Mass (Da) | Z | rT (min) | Isotopic Mass (Da) | Average Mass (Da) | Z |
|---|---|---|---|---|---|---|---|
| 3.6 | 1271.90 | 1273.92 | +2 | 39.5 | 1376.97 | 1378.98 | +2 |
| 1529.09 | 1531.09 | +2 | 1573.12 | 1574.12 | +3 | ||
| 1599.14 | 1601.12 | +3 | 1617.15 | 1618.15 | +3 | ||
| 1617.15 | 1619.15 | +3 | 2937.37 | 2939.38 | +5 | ||
| 2939.38 | 2941.40 | +5 | 2994.44 | 2997.44 | +5 | ||
| 2993.44 | 2996.47 | +5 | 3180.48 | 3182.49 | +5 | ||
| 33.3 | 2511.59 | 2513.60 | +4 | 42.3 | 1114.82 | 1115.83 | +2 |
| 2939.38 | 2941.40 | +5 | 1118.75 | 1119.75 | +2 | ||
| 3180.48 | 3183.50 | +5 | 1467.04 | 1469.04 | +2 | ||
| 34.4 | 1047.71 | 1048.75 | +2 | 1493.04 | 1496.07 | +2 | |
| 1070.80 | 1071.80 | +2 | 60.2 | 1467.03 | 1469.04 | +2 | |
| 1398.81 | 1400.91 | +2 | 3180.50 | 3183.50 | +5 | ||
| 1511.06 | 1513.07 | +2 | 61.9 | 1510.04 | 1511.06 | +2 | |
| 35.9 | 1557.12 | 1558.13 | +3 | 1617.15 | 1619.15 | +3 | |
| 1644.17 | 1646.18 | +3 | 3180.50 | 3183.50 | +5 | ||
| 2467.56 | 2469.57 | +4 | |||||
| 2511.58 | 2513.62 | +4 | |||||
| 3278.07 | 3279.06 | +5 |
2.6. Peptide Identification
| Fraction | Monoisotopic mass MH+ | Z | MS/MS derived sequence | Data base ID. All UniProtKB | AA % ID | Species | Matched peptide |
|---|---|---|---|---|---|---|---|
| Ctenitoxin-Pb48 | 1341.542 | +2 | CDXAMGNCXCK | P83911 | 100 | Phoneutria reidy | ω-ctenitoxin-Pr1a (Tx3-7) |
| O76201 | 100 | Phonuetria nigriventer | ω-ctenitoxin-Pn1a (Tx3-2) | ||||
| O76200 | 54 | Phonuetria nigriventer | κ-ctenitoxin-Pn1a (Tx3-1) | ||||
| Ctenitoxin-Pb53 | 1265.667 | +3 | ESNFKXGMAXR | P83905 | 100 | Phoneutria keyserlingi | U2-Ctenitoxin-Pk1a (U2-CNTX-Pk1a) |
| P61229 | 81 | Phonuetria nigriventer | U1-Ctenitoxin-Pn1a (U2-CNTX-Pn1a) | ||||
| P59367 | 63 | Phonuetria nigriventer | Ctenitoxin-Pn1a (Tx4(5-5)) |
2.7. Larvicidal Activity
3. Discussion
4. Materials and Methods
4.1. Spiders
4.2. Venom Extraction
4.3. Physicochemical Properties
4.4. Indirect Hemolytic Activity
4.5. PLA2 Activity
4.6. Proteolytic Activity
4.7. Electrophoretic Profile
4.8. Chromatographic Profile
4.9. Peptide Mass Determination by High-Resolution LC-MS
4.10. Peptide Identification by LC-MS/MS
4.11. Larvicidal Activity
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Florez, E.; Ortiz, A.; Montoya, M. Accidentes por mordedura de la araña de las bananeras Phoneutria boliviensis (Araneae:Ctenidae) en la región de Urabá, Colombia. Entomólogo 2003, 96, 1–4. [Google Scholar]
- Foelix, R. Biology of the Spiders; Oxford University Press: New York, NY, USA, 2001; pp. 3–11. [Google Scholar]
- Martins, R.; Bertani, R. The non-Amazonian species of the Brazilian wandering spiders of the genus Phoneutria perty, 1833 (Araneae:Ctenidae), with the description of a new species. Zootaxa 2007, 1526, 1–36. [Google Scholar]
- Simó, M.; Brescovit, A.D. Revision and cladistic analysis of the neotropical spider genus Phoneutria Perty, 1833 (Araneae, Ctenidae), with notes on related Ctenidae. Bull. Br. Aracnol. Soc. 2001, 12, 67–82. [Google Scholar]
- Hazzi, N.A.; Valderrama-Ardila, C.; Brescovit, A.D.; Polotow, D.; Simó, M. New records and geographical distribution of ctenid spiders (Araneae:Ctenidae) in Colombia. ZOOTAXA 2013, 3709. [Google Scholar] [CrossRef]
- Vassilevski, A.A.; Kozlov, S.A.; Grishin, E.V. Molecular diversity of spider venom. Biochemistry 2009, 74, 1505–1534. [Google Scholar] [CrossRef] [PubMed]
- Escoubas, P.; Quinton, L.; Nicholson, G.M. Venomics: Unravelling the complexity of animal venoms with mass spectrometry. J. Mass Spectrom. 2008, 43, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Escoubas, P.; Rash, L. Tarantulas: Eight-legged pharmacists and combinatorial chemists. Toxicon 2004, 43, 555–574. [Google Scholar] [CrossRef] [PubMed]
- Estrada, G.S.; Vargas, M.L.; Ramirez, A.; Quintana, C.J. Milking and partial characterization pamphobeteus spp (aranae; theraphosidae) venom, from the colombian andean region. Toxicon 2012, 60, 231–232. [Google Scholar]
- Nicholson, G.M. Spider venom peptides. In Handbook of Biologically Active Peptides; Hastin, A., Ed.; Elsevier: Los Angeles, CA, USA, 2006; p. 1640. [Google Scholar]
- Nicholson, G.M. Insect-selective spider toxins targeting voltage-gated sodium channels. Toxicon 2007, 49, 490–512. [Google Scholar] [CrossRef] [PubMed]
- Munoz, L.J.V.; Estrada-Gomez, S. Purification and characterization of venom components as source for antibiotics. Mini-Rev. Org. Chem. 2014, 11, 15–27. [Google Scholar] [CrossRef]
- Escoubas, P.; Diochot, S.; Corzo, G. Structure and pharmacology of spider venom neurotoxins. Biochimie 2000, 82, 893–907. [Google Scholar] [CrossRef]
- Rocha-e-Silva, T.A.; Collares-Buzato, C.B.; da Cruz-Hofling, M.A.; Hyslop, S. Venom apparatus of the Brazilian tarantula Vitalius dubius Mello-Leitao 1923 (Theraphosidae). Cell Tissue Res. 2009, 335, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Estrada, G.; Villegas, E.; Corzo, G. Spider venoms: A rich source of acylpolyamines and peptides as new leads for CNS drugs. Nat. Prod. Rep. 2007, 24, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Gomez, S.; Vargas Munoz, L.J.; Quintana Castillo, J.C. Extraction and partial characterization of venom from the Colombian spider Pamphobeteus aff. nigricolor (Aranae:Theraphosidae). Toxicon 2013, 76, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.; Pimenta, A.M.; Bemquerer, M.P.; Santoro, M.M.; Beirao, P.S.; Lima, M.E.; Figueiredo, S.G.; Bloch, C., Jr.; Vasconcelos, E.A.; Campos, F.A.; et al. Comparison of the partial proteomes of the venoms of Brazilian spiders of the genus Phoneutria. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 142, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, M.D.; Defigueiredo, S.G.; Valentim, A.D.; Diniz, C.R.; Voneickstedt, V.R.D.; Gilroy, J.; Richardson, M. Purification and amino-acid-sequences of 6 Tx3-type neurotoxins from the venom of the brazilian armed spider phoneutria-nigriventer (Keys). Toxicon 1993, 31, 35–42. [Google Scholar] [CrossRef]
- Oliveira, L.C.; de Lima, M.E.; Pimenta, A.M.; Mansuelle, P.; Rochat, H.; Cordeiro, M.N.; Richardson, M.; Figueiredo, S.G. PnTx4–3, a new insect toxin from Phoneutria nigriventer venom elicits the glutamate uptake inhibition exhibited by PhTx4 toxic fraction. Toxicon 2003, 42, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Rezende, L.; Cordeiro, M.N.; Oliveira, E.B.; Diniz, C.R. Isolation of neurotoxic peptides from the venom of the armed spider phoneutria-nigriventer. Toxicon 1991, 29, 1225–1233. [Google Scholar] [CrossRef]
- Araujo, D.A.; Cordeiro, M.N.; Diniz, C.R.; Beirao, P.S. Effects of a toxic fraction, PhTx2, from the spider Phoneutria nigriventer on the sodium current. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1993, 347, 205–208. [Google Scholar]
- Moura, J.R.; Prado, M.A.M.; Gomez, M.V.; Kalapothakis, E.; Diniz, C.R.; Cordeiro, M.N.; Romano-Silva, M.A. Investigation of the effect of PhTx2, from the venom of the spider Phoneutria nigriventer, on the release of [3H]-acetylcholine from rat cerebrocortical synaptosomes. Toxicon 1998, 36, 1189–1192. [Google Scholar] [CrossRef]
- Brazil, O.V.; Fontana, M.D. Toxins as tools in the study of sodium channel distribution in the muscle fibre membrane. Toxicon 1993, 31, 1085–1098. [Google Scholar] [CrossRef]
- Cordeiro, M.N.; Diniz, C.R.; do Carmo Valentim, A.; von Eickstedt, V.R.D.; Gilroy, J.; Richardson, M. The purification and amino acid sequences of four Tx2 neurotoxins from the venom of the Brazilian “armed” spider Phoneutria nigriventer (Keys). FEBS Lett. 1992, 310, 153–156. [Google Scholar] [CrossRef]
- Prado, M.A.; Guatimosim, C.; Gomez, M.V.; Diniz, C.R.; Cordeiro, M.N.; Romano-Silva, M.A. A novel tool for the investigation of glutamate release from rat cerebrocortical synaptosomes: The toxin Tx3–3 from the venom of the spider Phoneutria nigriventer. Biochem. J. 1996, 314 Pt 1, 145–150. [Google Scholar] [PubMed]
- Kushmerick, C.; Kalapothakis, E.; Beirao, P.S.; Penaforte, C.L.; Prado, V.F.; Cruz, J.S.; Diniz, C.R.; Cordeiro, M.N.; Gomez, M.V.; Romano-Silva, M.A.; et al. Phoneutria nigriventer toxin Tx3-1 blocks A-type K+ currents controlling Ca2+ oscillation frequency in GH3 cells. J. Neurochem. 1999, 72, 1472–1481. [Google Scholar] [CrossRef] [PubMed]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Gasnier, T.R.; de Azevedo, C.S.; Torres-Sanchez, M.P.; Hofer, H. Adult size of eight hunting spider species in central Amazonia: Temporal variations and sexual dimorphisms. J. Arachnol. 2002, 30, 146–154. [Google Scholar] [CrossRef]
- Estrada-Gómez, S.; Cupitra, N.; Arango, W.; Muñoz, L. Intraspecific variation of centruroides edwardsii venom from two regions of colombia. Toxins 2014, 6, 2082–2096. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, E.F.; Camargos, T.S.; Zamudio, F.Z.; Silva, L.P.; Bloch, C.; Caixeta, F.; Schwartz, C.A.; Possani, L.D. Mass spectrometry analysis, amino acid sequence and biological activity of venom components from the Brazilian scorpion Opisthacanthus cayaporum. Toxicon 2008, 51, 1499–1508. [Google Scholar] [CrossRef] [PubMed]
- Valdez-Velazquez, L.L.; Quintero-Hernandez, V.; Romero-Gutierrez, M.T.; Coronas, F.I.; Possani, L.D. Mass fingerprinting of the venom and transcriptome of venom gland of scorpion Centruroides tecomanus. PLoS ONE 2013, 8, e66486. [Google Scholar] [CrossRef] [PubMed]
- Veiga, S.S.; da Silveira, R.B.; Dreyfuss, J.L.; Haoach, J.; Pereira, A.M.; Mangili, O.C.; Gremski, W. Identification of high molecular weight serine-proteases in Loxosceles intermedia (brown spider) venom. Toxicon 2000, 38, 825–839. [Google Scholar] [CrossRef]
- Pimenta, A.M.; Rates, B.; Bloch, C., Jr.; Gomes, P.C.; Santoro, M.M.; de Lima, M.E.; Richardson, M.; Cordeiro, M.N. Electrospray ionization quadrupole time-of-flight and matrix-assisted laser desorption/ionization tandem time-of-flight mass spectrometric analyses to solve micro-heterogeneity in post-translationally modified peptides from Phoneutria nigriventer (Aranea, Ctenidae) venom. Rapid Commun. Mass Spectrom. 2005, 19, 31–37. [Google Scholar] [PubMed]
- Gentz, M.C.; Jones, A.; Clement, H.; King, G.F. Comparison of the peptidome and insecticidal activity of venom from a taxonomically diverse group of theraphosid spiders. Toxicon 2009, 53, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Giangaspero, A.; Sandri, L.; Tossi, A. Amphipathic alpha helical antimicrobial peptides-A systematic study of the effects of structural and physical properties on biological activity. Eur. J. Biochem. 2001, 268, 5589–5600. [Google Scholar] [CrossRef] [PubMed]
- Sousa, S.R.; Vetter, I.; Lewis, R.J. Venom peptides as a rich source of cav2.2 channel blockers. Toxins 2013, 5, 286–314. [Google Scholar] [CrossRef] [PubMed]
- Vieira, L.B.; Pimenta, A.M.; Richardson, M.; Bemquerer, M.P.; Reis, H.J.; Cruz, J.S.; Gomez, M.V.; Santoro, M.M.; Ferreira-de-Oliveira, R.; Figueiredo, S.G.; et al. Leftward shift in the voltage-dependence for Ca2+ currents activation induced by a new toxin from Phoneutria reidyi (Aranae, Ctenidae) venom. Cell. Mol. Neurobiol. 2007, 27, 129–146. [Google Scholar] [CrossRef] [PubMed]
- De Figueiredo, S.G.; de Lima, M.E.; Nascimento Cordeiro, M.; Diniz, C.R.; Patten, D.; Halliwell, R.F.; Gilroy, J.; Richardson, M. Purification and amino acid sequence of a highly insecticidal toxin from the venom of the brazilian spider Phoneutria nigriventer which inhibits NMDA-evoked currents in rat hippocampal neurones. Toxicon 2001, 39, 309–317. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Avila, C.; Rojas, E.; Cerdas, L. An alternative in vitro method for testing the potency of the polyvalent antivenom produced in Costa Rica. Toxicon 1988, 26, 411–413. [Google Scholar] [CrossRef]
- Cho, W.; Kezdy, F.J. Chromogenic substrates and assay of phospholipases A2. Methods Enzymol. 1991, 197, 75–79. [Google Scholar] [PubMed]
- Holzer, M.; Mackessy, S.P. An aqueous endpoint assay of snake venom phospholipase A2. Toxicon 1996, 34, 1149–1155. [Google Scholar] [CrossRef]
- Wang, W.J.; Shih, C.H.; Huang, T.F. A novel P-I class metalloproteinase with broad substrate-cleaving activity, agkislysin, from Agkistrodon acutus venom. Biochem. Biophys. Res. Commun. 2004, 324, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Patiño, A.C.; Pereañez, J.A.; Gutiérrez, J.M.; Rucavado, A. Biochemical and biological characterization of two serine proteinases from Colombian Crotalus durissus cumanensis snake venom. Toxicon 2013, 63, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.; Gutierrez, J.M.; Angulo, Y.; Sanz, L.; Juarez, P.; Calvete, J.J.; Lomonte, B. Isolation of an acidic phospholipase A2 from the venom of the snake Bothrops asper of Costa Rica: Biochemical and toxicological characterization. Biochimie 2010, 92, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estrada-Gomez, S.; Muñoz, L.J.V.; Lanchero, P.; Latorre, C.S. Partial Characterization of Venom from the Colombian Spider Phoneutria Boliviensis (Aranae:Ctenidae). Toxins 2015, 7, 2872-2887. https://doi.org/10.3390/toxins7082872
Estrada-Gomez S, Muñoz LJV, Lanchero P, Latorre CS. Partial Characterization of Venom from the Colombian Spider Phoneutria Boliviensis (Aranae:Ctenidae). Toxins. 2015; 7(8):2872-2887. https://doi.org/10.3390/toxins7082872
Chicago/Turabian StyleEstrada-Gomez, Sebastian, Leidy Johana Vargas Muñoz, Paula Lanchero, and Cesar Segura Latorre. 2015. "Partial Characterization of Venom from the Colombian Spider Phoneutria Boliviensis (Aranae:Ctenidae)" Toxins 7, no. 8: 2872-2887. https://doi.org/10.3390/toxins7082872
APA StyleEstrada-Gomez, S., Muñoz, L. J. V., Lanchero, P., & Latorre, C. S. (2015). Partial Characterization of Venom from the Colombian Spider Phoneutria Boliviensis (Aranae:Ctenidae). Toxins, 7(8), 2872-2887. https://doi.org/10.3390/toxins7082872