
Multi-Year Assessment of Toxic Genotypes and Microcystin Concentration in Northern Lake Taihu, China


Abstract
:1. Introduction
2. Results
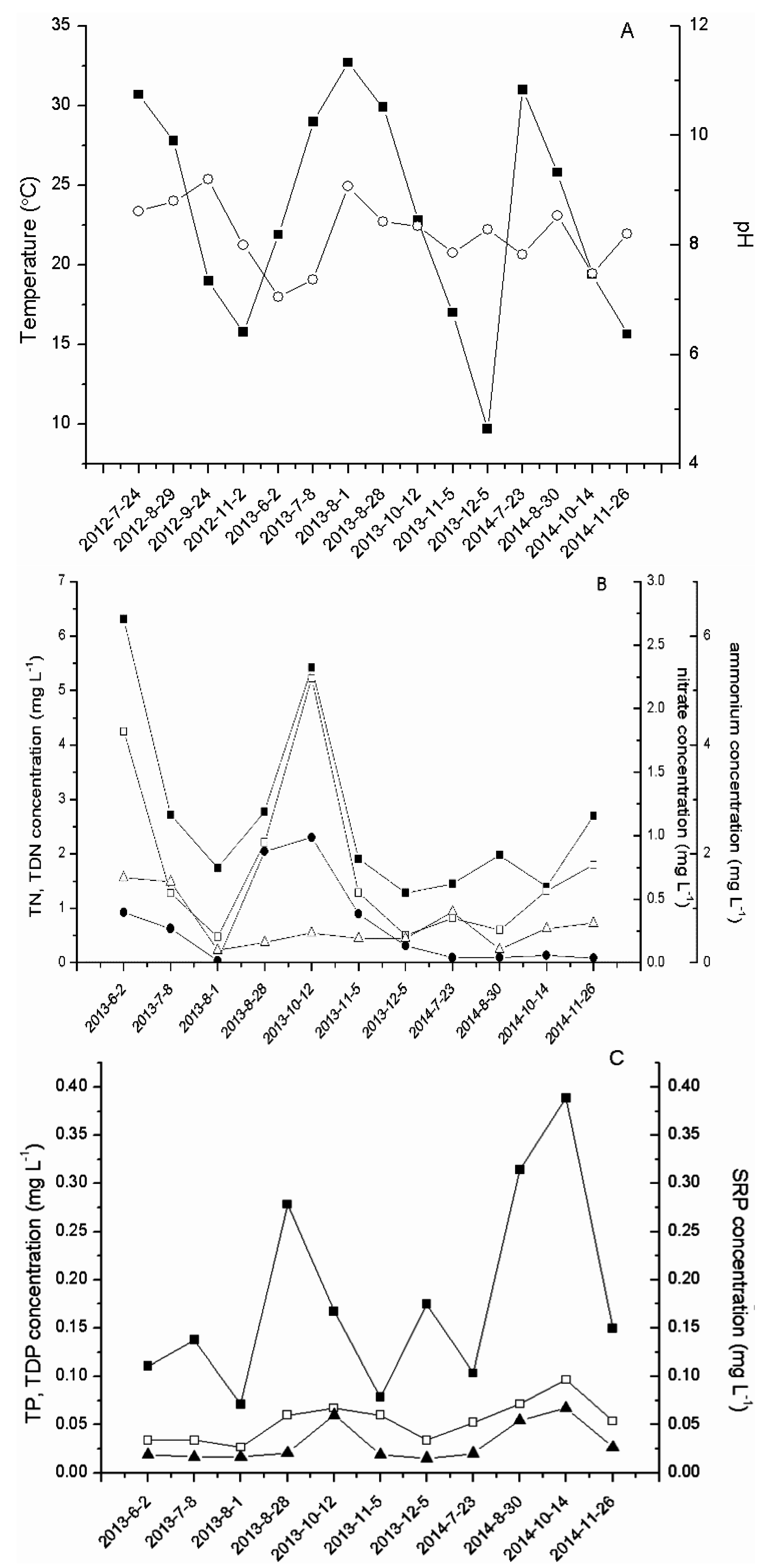
2.1. Physicochemical Parameters

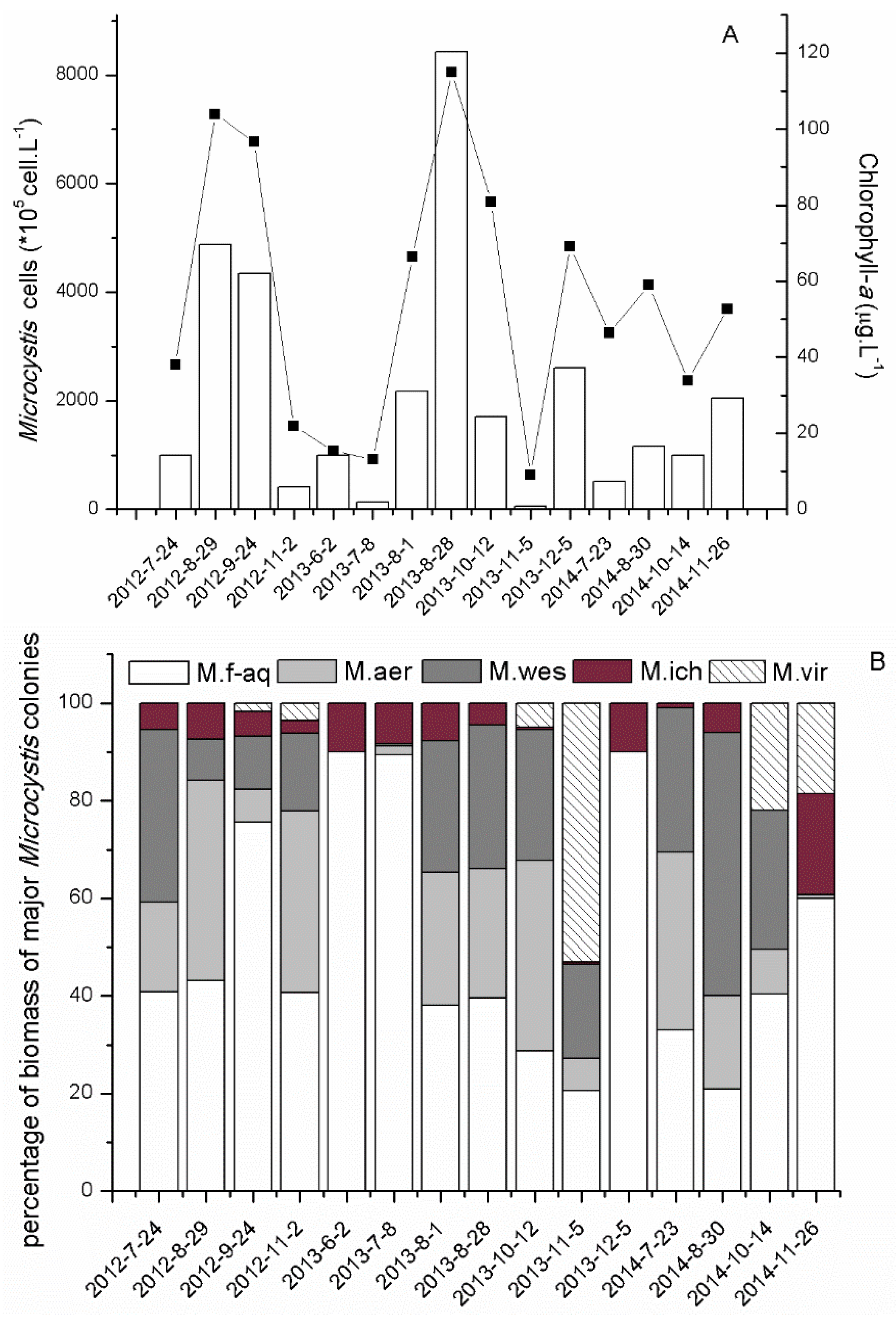
2.2. Dynamics of Abundance and Succession of Microcystis

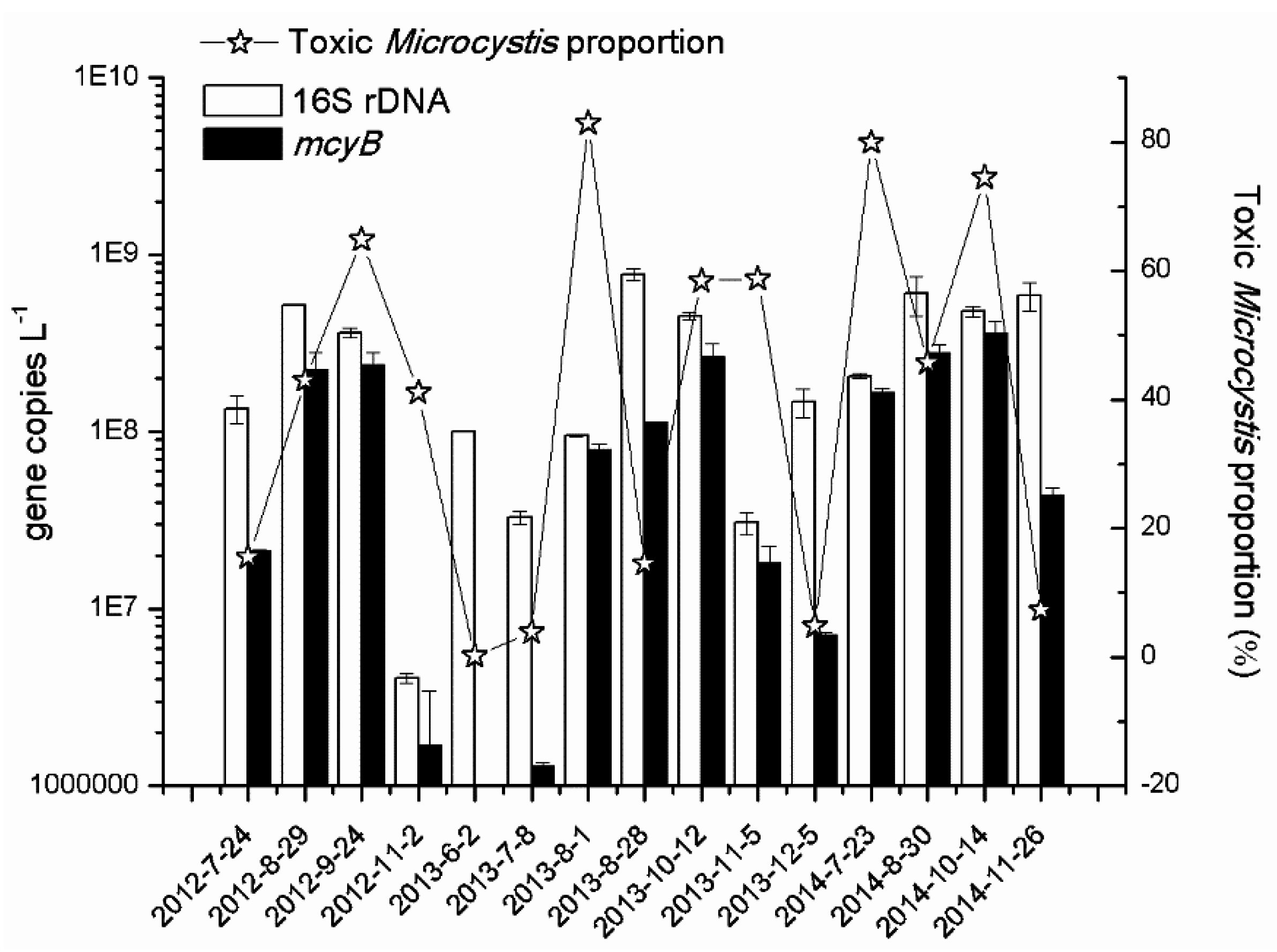
2.3. Dynamics of Toxic Microcystis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Sequence (5′–3′) | Length (bp) | Reference | Efficiency |
|---|---|---|---|---|
| 16S rDNA | ATGTGCCGCGAGGTGAAACCTAAT | 200 | Neilan et al., 1997 [42] | 99.9% |
| TTACAATCCAAAGACCTTCCTCCC | ||||
| mcyB | CCTACCGAGCGCTTGGG | 78 | Kurmayer et al., 2003 [11] | 93.6% |
| GAAAATCCCCTAAAGATTCCTGAGT |

2.4. Microcystin Content of Field Samples and Cultivated Morphospecies
| Site | Sampling Date (Year-Month-Day) | RR (µg/g) | LR (µg/g) | MCs (µg/g) | RR (%) |
|---|---|---|---|---|---|
| N2 | 2012-7-24 | 220.5 ± 19.3 | 143.7 ± 5.6 | 363.2 ± 74.3 | 61 |
| 2012-8-29 | 672.8 ± 178.2 | 349.3 ± 45.5 | 1022.1 ± 146.7 | 66 | |
| 2012-9-24 | 251.6 ± 38.5 | 155.0 ± 111.5 | 406.6 ± 22.8 | 62 | |
| 2012-11-2 | 615.4 ± 23.3 | 525.7 ± 127.6 | 1141.1 ± 104.3 | 54 | |
| 2013-6-2 | 78.9 ± 68.9 | 14.9 ± 15.0 | 93.8 ± 53.9 | 84 | |
| 2013-7-8 | 250.9 ± 2.9 | 145.5 ± 37.1 | 396.4 ± 62.7 | 63 | |
| 2013-8-1 | 359.9 ± 47.3 | 356.4 ± 63.2 | 716.3 ± 110.5 | 50 | |
| 2013-8-28 | 165.1 ± 2.9 | 173.5 ± 5.6 | 338.6 ± 8.5 | 48 | |
| 2013-10-12 | 769.9 ± 4.0 | 686.7 ± 81.5 | 1455.6 ± 77.5 | 52 | |
| 2013-11-5 | 838 | 826 | 1664 | 50 | |
| 2013-12-5 | ND | 45 | 45 | ND | |
| 2014-7-23 | 676.7 ± 173.3 | 642.0 ± 125.2 | 1318.7 ± 298.5 | 51 | |
| 2014-8-30 | 193.3 ± 11.8 | 152.4 ± 15.7 | 345.7 ± 27.5 | 55 | |
| 2014-10-14 | 769.6 ± 121.1 | 617.1 ± 50.3 | 1386.8 ± 171.4 | 55 | |
| 2014-11-26 | 229.2 ± 9.6 | 202.3 ± 3.5 | 431.5 ± 13.0 | 53 |
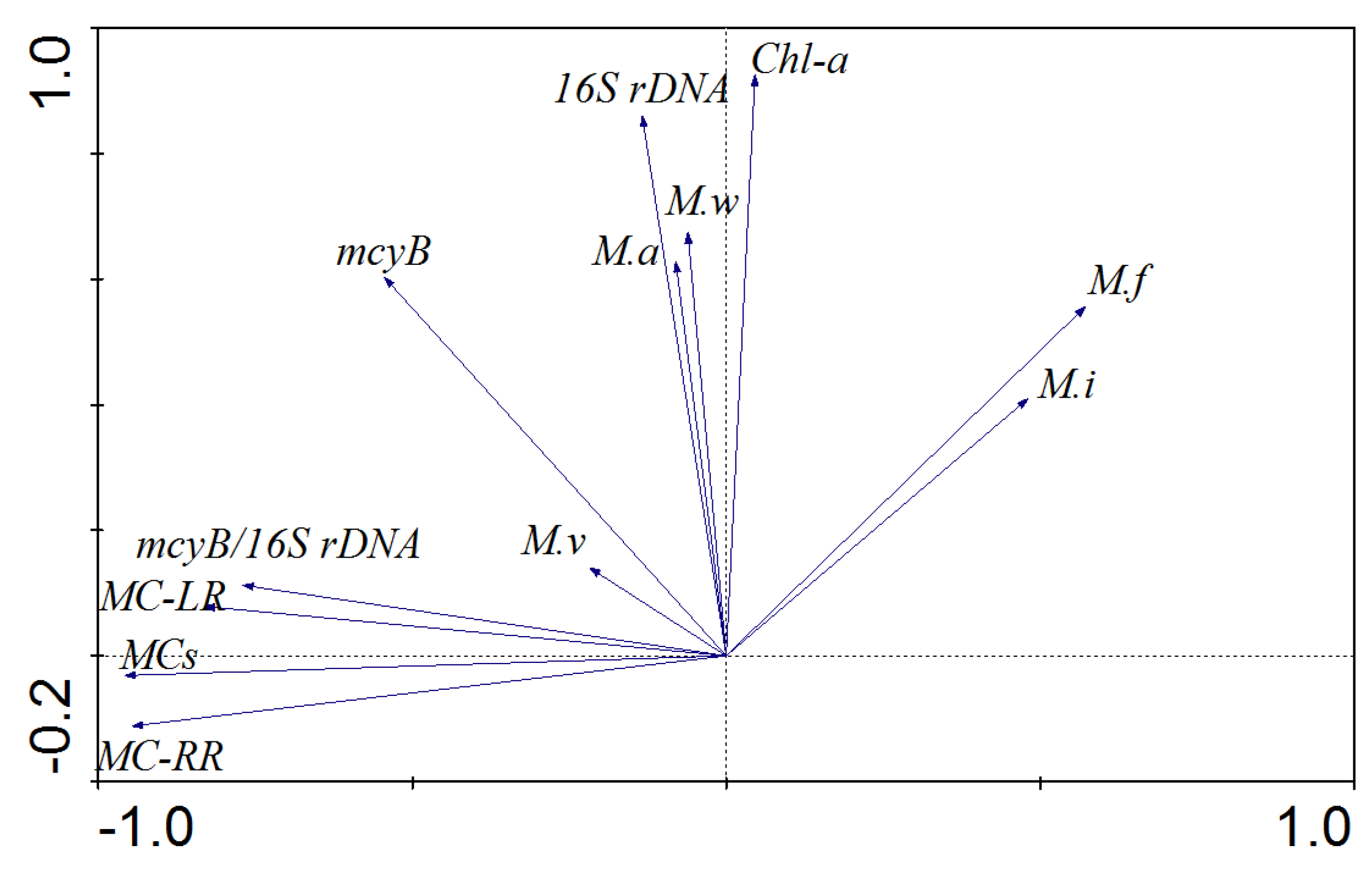
2.5. The Variations in the Microcystis Community and their Correlation with Environmental Factors

| Variable | Microcystis 16S rDNA (Gene Copies/L) | Microcystis mcyB (Gene Copies/L) | Microcystins (µg/g) |
|---|---|---|---|
| Microcystis cells (cells/L) | 0.664 ** | −0.248 | −0.371 |
| TN (mg/L) | 0.055 | 0.055 | −0.079 |
| TDN (mg/L) | 0.236 | −0.340 | 0.224 |
| NO3− (mg/L) | −0.249 | −0.173 | 0.012 |
| NH4+ (mg/L) | −0.309 | −0.136 | 0.100 |
| TP (mg/L) | 0.755 ** | 0.518 | −0.273 |
| TDP (mg/L) | 0.589 | 0.767 ** | 0.420 |
| SRP (mg/L) | 0.715 * | 0.784 ** | 0.374 |
| WT (°C) | 0.068 | 0.186 | −0.018 |
| pH | 0.329 | 0.436 | −0.118 |
3. Discussion
4. Experimental Section
4.1. Study Site and Sampling

4.2. Physicochemical Parameters and Nutrients
4.3. Identification and Counting of Microcystis Morphospecies
4.4. DNA Extraction and qPCR
4.5. Extraction and Determination of Particulate Microcystins
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Falconer, I.R.; Humpage, A.R. Health risk assessment of cyanobacterial (blue-green algal) toxins in drinking water. Int. J. Environ. Res. Pubulic Health 2005, 2, 43–50. [Google Scholar] [CrossRef]
- Codd, G.A.; Lindsay, J.; Young, F.M.; Morrison, L.F.; Metcalf, J.S. From mass mortalities to management measures. In Harmful Cyanobacteria; Huisman, J., Matthijs, H.C.P., Visser, P.M., Eds.; Springer Netherlands: Dordrecht, The Netherland, 2005; pp. 1–23. [Google Scholar]
- Honkanen, R.E.; Zwiller, J.; Moore, R.E.; Daily, S.L.; Khatra, B.S.; Dukelow, M.; Boynton, A.L. Characterization of microcystin-LR, a potent inhibitor of type 1 and type 2A protein phosphatases. J. Biol. Chem. 1990, 265, 19401–19404. [Google Scholar] [PubMed]
- Meiβner, K.; Dittmann, E.; Börner, T. Toxic and non-toxic strains of the cyanobacterium Microcystis aeruginosa contain sequences homologous to peptide synthetase genes. FEMS Microbiol. Lett. 1996, 135, 295–303. [Google Scholar] [CrossRef]
- Dittmann, E.; Neilan, B.A.; Erhard, M.; Döhren, H.V.; Börner, T. Insertional mutagenesis of a peptide synthetase gene that is responsible for hepatotoxin production in the cyanobacterium Microcystis aeruginosa PCC 7806. Mol. Microbiol. 1997, 26, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Tillett, D.; Parker, D.L.; Neilan, B.A. Detection of toxigenicity by a probe for the microcystin synthetase a gene (mcyA) of the cyanobacterial genus Microcystis: Comparison of toxicities with 16S rRNA and phycocyanin operon (phycocyanin intergenic spacer) phylogenies. Appl. Environ. Microb. 2001, 67, 2810–2818. [Google Scholar] [CrossRef] [PubMed]
- Mikalsen, B.; Boison, G.; Skulberg, O.M.; Fastner, J.; Davies, W.; Gabrielsen, T.M.; Rudi, K.; Jakobsen, K.S. Natural variation in the microcystin synthetase operon mcyABC and impact on microcystin production in Microcystis strains. J. Bacteriol. 2003, 185, 2774–2785. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; von Döhren, H. Cyanobacterial peptides-nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed]
- Bortoli, S.; Oliveira-Silva, D.; Krüger, T.; Dörr, F.; Colepicolo, P.; Volmer, D.; Pinto, E. Growth and microcystin production of a Brazilian Microcystis aeruginosa strain (LTPNA 02) under different nutrient conditions. Rev. Bras. Farmacogn. 2014, 24, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Pant, S.C.; Vijayaraghavan, R.; Lakshmana Rao, P.V. Comparative toxicity evaluation of cyanobacterial cyclic peptide toxin microcystin variants (LR, RR, YR) in mice. Toxicology 2003, 188, 285–296. [Google Scholar] [CrossRef]
- Kurmayer, R.; Kutzenberger, T. Application of real-time PCR for quantification of microcystin genotypes in a population of the toxic cyanobacterium Microcystis sp. Appl. Environ. Microb. 2003, 69, 6723–6730. [Google Scholar] [CrossRef]
- Rinta-Kanto, J.M.; Ouellette, A.; Boyer, G.L.; Twiss, M.R.; Bridgeman, T.B.; Wilhelm, S.W. Quantification of toxic Microcystis spp. during the 2003 and 2004 blooms in western Lake Erie using quantitative real-time PCR. Environ. Sci. Technol. 2005, 39, 4198–4205. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Noda, N.; Tsuneda, S.; Saito, T.; Itayama, T.; Inamori, Y. Highly sensitive real-time PCR assay for quantification of toxic cyanobacteria based on microcystin synthetase A gene. J. Biosci. Bioeng. 2006, 102, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Hotto, A.M.; Satchwell, M.F.; Berry, D.L.; Gobler, C.J.; Boyer, G.L. Spatial and temporal diversity of microcystins and microcystin-producing genotypes in Oneida Lake, NY. Harmful Algae 2008, 7, 671–681. [Google Scholar] [CrossRef]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Okello, W.; Portmann, C.; Erhard, M.; Gademann, K.; Kurmayer, R. Occurrence of microcystin-producing cyanobacteria in Ugandan freshwater habitats. Environ. Toxicol. 2010, 25, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Choi, G.G.; Ahn, C.Y.; Oh, Hee-Mock.; Ravi, A.K.; Asthana, R.K. Dynamics of microcystin production and quantification of potentially toxigenic Microcystis sp. using real-time PCR. Water Res. 2012, 46, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Liu, X.L.; Tan, J.; Li, D.; Yang, H. Diversity and dynamics of microcystin—Producing cyanobacteria in China's third largest lake, Lake Taihu. Harmful Algae 2009, 8, 637–644. [Google Scholar] [CrossRef]
- Sabart, M.; Pobel, D.; Briand, E.; Combourieu, B.; Salencon, M.J.; Humbert, J.F.; Latour, D. Spatiotemporal variations in microcystin concentrations and in the proportions of microcystin-producing cells in several Microcystis aeruginosa populations. Appl. Environ. Microb. 2010, 76, 4750–4759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guedes, I.A.; da Costa Leite, D.M.; Manhães, L.A.; Bisch, P.M.; Azevedo, S.; Pacheco, A. Fluctuations in microcystin concentrations, potentially toxic Microcystis and genotype diversity in a cyanobacterial community from a tropical reservoir. Harmful Algae 2014, 39, 303–309. [Google Scholar] [CrossRef]
- Jacoby, J.; Collier, D.C.; Welch, E.B.; Hardy, F.J.; Crayto, M. Environmental factors associated with a toxic bloom of Microcystis aeruginosa. Can. J. Fish. Aquat. Sci. 2000, 57, 231–240. [Google Scholar] [CrossRef]
- Kotak, B.G.; Lam, A.; Prepas, E.E.; Hrudey, S. Role of chemical and physical variables in regulating microcystin-LR concentration in phytoplankton of eutrophic lakes. Can. J. Fish. Aquat. Sci. 2000, 57, 1584–1593. [Google Scholar] [CrossRef]
- Izydorczyk, K.; Jurczak, T.; Wojtal-Frankiewicz, A.; Skowron, A.; Mankiewicz-Boczek, J.; Tarczyńska, M. Influence of abiotic and biotic factors on microcystin content in Microcystis aeruginosa cells in a eutrophic temperate reservoir. J. Plankton Res. 2008, 30, 393–400. [Google Scholar] [CrossRef]
- Horst, G.P.; Sarnelle, O.; White, J.D.; Hamilton, S.K.; Kaul, R.; Bressie, J.D. Nitrogen availability increases the toxin quota of a harmful cyanobacterium, Microcystis aeruginosa. Water Res. 2014, 54, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Park, H.D.; Kim, B.; Kim, E.; Okino, T. Hepatotoxic microcystins and neurotoxic anatoxin-a in cyanobacterial blooms from Korean lakes. Environ. Toxicol. Water Qual. 1998, 13, 225–234. [Google Scholar] [CrossRef]
- Honma, T.; Park, H.D. Influences of nitrate and phosphate concentrations on Microcystis species composition and microcystin concentration in Lake Suwa. J. Jpn. Soc. Water Environ. 2005, 28, 373–378. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Nakahara, H. Seasonal variations in the morphology of bloom-forming cyanobacteria in a eutrophic pond. Limnology 2009, 10, 185–193. [Google Scholar] [CrossRef]
- Kurmayer, R.; Dittmann, E.; Fastner, J.; Chorus, I. Diversity of microcystin genes within a population of the toxic cyanobacterium Microcystis spp. in lake Wannsee (Berlin, Germany). Microb. Ecol. 2002, 43, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Via-Ordorika, L.; Fastner, J.; Kurmayer, R.; Hisbergues, M.; Dittmann, E.; Komarek, J.; Erhard, M.; Chorus, I. Distribution of microcystin-producing and non-microcystin-producing Microcystis sp. in European freshwater bodies: Detection of microcystins and microcystin genes in individual colonies. Syst. Appl. Microbiol. 2004, 27, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Peng, L.; Wan, N.; Song, L. Mechanism study on the frequent variations of cell-bound microcystins in cyanobacterial blooms in Lake Taihu: Implications for Water Quality Monitoring and Assessments. Chemosphere 2009, 77, 1585–1593. [Google Scholar] [CrossRef] [PubMed]
- Kurmayer, R.; Christiansen, G.; Chorus, I. The abundance of microcystin-producing genotypes correlates positively with colony size in Microcystis sp. and determines its microcystin net production in Lake Wannsee. Appl. Environ. Microb. 2003, 69, 787–795. [Google Scholar] [CrossRef]
- Wang, X.; Sun, M.; Xie, M.; Liu, M.; Luo, L.; Li, P.F.; Kong, F.X. Differences in microcystin production and genotype composition among Microcystis colonies of different sizes in Lake Taihu. Water Res. 2013, 47, 5659–5669. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Xu, P.; Wu, Q.; Luo, L.C.; Zhang, Y.L. Environmental issues of lake Taihu, China. Hydrobiologia 2007, 581, 3–14. [Google Scholar] [CrossRef]
- Song, L.; Chen, W.; Peng, L.; Wan, N.; Gan, N.Q.; Zhang, X.M. Distribution and bioaccumulation of microcystins in water columns: A Systematic Investigation into the Environmental Fate and the Risks Associated with Microcystins in Meiliang Bay, Lake Taihu. Water Res. 2007, 41, 2853–2864. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Song, L.; Peng, L.; Wan, N.; Zhang, X.; Gan, N. Reduction in microcystin concentrations in large and shallow lakes: Water and Sediment-Interface Contributions. Water Res. 2008, 42, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Niu, Y.; Xie, P.; Chen, J.; Ma, Z.; Tao, M.; Qi, M.; Wu, L.; Guo, L. Factors affecting temporal and spatial variations of microcystins in Gonghu Bay of Lake Taihu, with potential risk of microcystin contamination to human health. Sci. World J. 2010, 10, 1795–1809. [Google Scholar] [CrossRef] [PubMed]
- Otten, T.G.; Paerl, H.W. Phylogenetic inference of colony isolates comprising seasonal Microcystis blooms in Lake Taihu, China. Microb. Ecol. 2011, 62, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, M.; Wang, J.; Yang, L.; Luo, L.; Li, P.; Kong, F. Microcystis genotype succession and related environmental factors in Lake Taihu during cyanobacterial blooms. Microb. Ecol. 2012, 64, 986–999. [Google Scholar] [CrossRef] [PubMed]
- Tao, M.; Xie, P.; Chen, J.; Qin, B.; Zhang, D.; Niu, Y.; Zhang, M.; Wang, Q.; Wu, L. Use of a generalized additive model to investigate key abiotic factors affecting microcystin cellular quotas in heavy bloom areas of Lake Taihu. PLoS ONE 2012, 7, e32020. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xie, P.; Zhang, D.; Wen, Z. Seasonal dynamics of microcystins with associated biotic and abiotic parameters in two bays of Lake Taihu, the third largest freshwater lake in China. Bull. Environ. Contam. Toxicol. 2008, 80, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Otten, T.G.; Xu, H.; Qin, B.; Zhu, G.; Paerl, H.W. Spatiotemporal patterns and ecophysiology of toxigenic Microcystis blooms in Lake Taihu, China: implications for water quality management. Environ. Sci. Technol. 2012, 46, 3480–3488. [Google Scholar] [CrossRef] [PubMed]
- Neilan, B.A.; Jacobs, D.; Blackall, L.L.; Hawkins, P.R.; Cox, P.T.; Goodman, A.E. rRNA sequences and evolutionary relationships among toxic and nontoxic cyanobacteria of the genus Microcystis. Int. J. Syst. Bacteriol. 1997, 47, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Xu, H.; McCarthy, M.J.; Zhu, G.; Qin, B.; Li, Y.; Gardner, W. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The Need for a Dual Nutrient (N & P) Management Strategy. Water Res. 2011, 45, 1973–1983. [Google Scholar] [PubMed]
- Li, D.; Kong, F.; Shi, X.; Ye, L.; Yu, Y.; Yang, Z. Quantification of microcystin-producing and non-microcystin producing Microcystis populations during the 2009 and 2010 blooms in Lake Taihu using quantitative real-time PCR. J. Environ. Sci. 2012, 24, 284–290. [Google Scholar] [CrossRef]
- Yu, L.; Kong, F.; Zhang, M.; Yang, Z.; Shi, X.; Du, M. The Dynamics of Microcystis Genotypes and Microcystin Production and Associations with Environmental Factors during Blooms in Lake Chaohu, China. Toxins 2014, 6, 3238–3257. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Wu, Y.; Song, L.; Gan, N. Ecological dynamics of toxic Microcystis spp. and microcystin-degrading bacteria in Dianchi Lake, China. Appl. Environ. Microb. 2014, 80, 1874–1881. [Google Scholar] [CrossRef] [PubMed]
- Rantala, A.; Rajaniemi-Wacklin, P.; Lyra, C.; Lepisto, L.; Rintala, J.; MankiewiczBoczek, J.; Sivonen, K. Detection of microcystin-producing cyanobacteria in Finnish lakes with genus-specific microcystin synthetase gene E (mcyE) PCR and associations with environmental factors. Appl. Environ. Microb. 2006, 72, 6101–6110. [Google Scholar] [CrossRef] [PubMed]
- Rinta-Kanto, J.M.; Konopko, E.A.; DeBruyn, J.M.; Bourbonniere, R.A.; Boyer, G.L.; Wilhelm, S.W. Lake Erie Microcystis: Relationship between Microcystin Production, Dynamics of Genotypes and Environmental Parameters in a Large Lake. Harmful Algae 2009, 8, 665–673. [Google Scholar] [CrossRef]
- Zhu, W.; Li, M.; Dai, X.; Xiao, M. Differences in vertical distribution of Microcystis morphospecies composition in a shallow hypertrophic lake (Lake Taihu, China). Environ. Earth Sci. 2015, 73, 5721–5730. [Google Scholar] [CrossRef]
- Chan, W.S.; Recknagel, F.; Cao, H.; Park, H.D. Elucidation and short-term forecasting of microcystin concentrations in Lake Suwa (Japan) by means of artificial neural networks and evolutionary algorithms. Water Res. 2007, 41, 2247–2255. [Google Scholar] [CrossRef] [PubMed]
- Namikoshi, M.; Rinehart, K.L.; Sakai, R.; Stotts, R.R.; Dahlem, A.M.; Beasley, V.R.; Carmichael, W.W.; Evans, W.R. Identification of 12 hepatotoxins from a Homer Lake bloom of the cyanobacteria Microcystis aeruginosa, Microcystis viridis and Microcystis wesenbergii: Nine New Microcystins. J. Org. Chem. 1992, 57, 866–872. [Google Scholar] [CrossRef]
- Otsuka, S.; Suda, S.; Li, R.; Watanabe, M.; Oyaizu, H.; Matsumoto, S.; Watanabe, M. Phylogenetic relationships between toxic and non-toxic strains of the genus Microcystis based on 16S to 23S internal transcribed spacer sequence. FEMS Microbiol. Lett. 1999, 172, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.H.; Hidaka, T.; Tsuno, H. Quantification of toxic Microcystis and evaluation of its dominance ratio in blooms using real-time PCR. Environ. Sci. Technol. 2009, 43, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Nusch, E.A. Comparison of different methods for chlorophyll and phaeopigment determination. Archiv. Hydrobiol. Beih. Ergebn. Limnol. 1980, 14, 14–36. [Google Scholar]
- Chen, W.; Gan, N.Q.; Song, L.R. Photodegradation dynamics of pure microcystin variants with illumination of fixed wavelength UV-lights. Acta Chim. Sin. 2004, 62, 142–147. [Google Scholar]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, L.; Shan, K.; Lin, L.; Shen, W.; Huang, L.; Gan, N.; Song, L. Multi-Year Assessment of Toxic Genotypes and Microcystin Concentration in Northern Lake Taihu, China. Toxins 2016, 8, 23. https://doi.org/10.3390/toxins8010023
Hu L, Shan K, Lin L, Shen W, Huang L, Gan N, Song L. Multi-Year Assessment of Toxic Genotypes and Microcystin Concentration in Northern Lake Taihu, China. Toxins. 2016; 8(1):23. https://doi.org/10.3390/toxins8010023
Chicago/Turabian StyleHu, Lili, Kun Shan, Lizhou Lin, Wei Shen, Licheng Huang, Nanqin Gan, and Lirong Song. 2016. "Multi-Year Assessment of Toxic Genotypes and Microcystin Concentration in Northern Lake Taihu, China" Toxins 8, no. 1: 23. https://doi.org/10.3390/toxins8010023
APA StyleHu, L., Shan, K., Lin, L., Shen, W., Huang, L., Gan, N., & Song, L. (2016). Multi-Year Assessment of Toxic Genotypes and Microcystin Concentration in Northern Lake Taihu, China. Toxins, 8(1), 23. https://doi.org/10.3390/toxins8010023