
Pentahydroxyscirpene—Producing Strains, Formation In Planta, and Natural Occurrence

 ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Development and Validation of the Liquid Chromatographic-Tandem Mass Spectrometric (LC-MS/MS) Method
2.2. Molecular Identification of IFA189 as Fusarium kyushuense
2.3. PHS Production by Other NIV Producers
2.4. Tentative Identification of 4-Acetyl-Pentahydroxyscirpene
2.5. PHS-Impurity in Commercial NIV-Standards
2.6. Natural Occurrence of PHS
2.7. Toxicity of PHS to Wheat Ribosomes
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. LC-MS/MS Parameters
4.3. Sample Preparation and Method Validation
4.4. Molecular Classification of the Strain IFA189
4.5. Screening for PHS in Naturally Contaminated Cereals and in Known NIV Producing Strains
4.6. Plant Experiments
4.7. Toxicity Assays
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| (U)HPLC | (ultra-)high performance liquid chromatography |
| 15ADON | 15-acetyl-deoxynivalenol |
| 3ADON | 3-acetyl-deoxynivalenol |
| ACN | acetonitrile |
| ARS | Agricultural Research Service |
| b.w. | body weight |
| bp | base pairs |
| CONTAM | European Food Safety Authority Panel on Contaminants in the Food Chain |
| DON | deoxynivalenol |
| EFSA | European Food Safety Authority |
| FAO | Food and Agricultural Organization of the United Nations |
| FMM | Fusarium minimal medium |
| FUSX | fusarenon-X |
| IC50 | half maximal inhibitory concentration |
| JECFA | Joint FAO/WHO Expert Committee on Food Additives |
| LC-MS/MS | liquid chromatography-tandem mass spectrometry |
| LC-MS | liquid chromatography-mass spectrometry |
| MeOH | methanol |
| NCBI | National Center for Biotechnology Information |
| NIV | nivalenol |
| NMR | nuclear magnetic resonance |
| NRRL | Agricultural Research Service Culture Collection strain |
| PCR | polymerase chain reaction |
| PDA | potato dextrose agar |
| PHS | pentahydroxyscirpene |
| PMTDI | provisional maximum tolerable daily intake |
| QTOF | quadrupole time-of-flight mass spectrometer |
| TDI | tolerable daily intake |
| WHO | World Health Organization |
References
- Smith, J.E.; Solomons, G.L.; Lewis, C.W.; Anderson, J.G. Mycotoxins in Human Nutrition and Health; European Commission CG XII: Brussels, Belgium, 1994. [Google Scholar]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin occurrence in feed and feed raw materials worldwide: Long-term analysis with special focus on Europe and Asia. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K.; Gardiner, D.M.; Manners, J.M. On the trail of a cereal killer: Recent advances in Fusarium graminearum pathogenomics and host resistance. Mol. Plant Pathol. 2012, 13, 399–413. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Ward, T.J.; Aberra, D.; Kistler, H.C.; Aoki, T.; Orwig, N.; Kimura, M.; Bjornstad, A.; Klemsdal, S.S. Multilocus genotyping and molecular phylogenetics resolve a novel head blight pathogen within the Fusarium graminearum species complex from Ethiopia. Fungal Genet. Biol. 2008, 45, 1514–1522. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Ndoye, M.; Zhang, J.B.; Li, H.P.; Liao, Y.C. Population structure and genetic diversity of the Fusarium graminearum species complex. Toxins 2011, 3, 1020–1037. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Han, Y.K.; Kim, K.H.; Yun, S.H.; Lee, Y.W. Tri13 and Tri7 determine deoxynivalenol- and nivalenol-producing chemotypes of Gibberella zeae. Appl. Environ. Microbiol. 2002, 68, 2148–2154. [Google Scholar] [CrossRef] [PubMed]
- Alexander, N.J.; McCormick, S.P.; Waalwijk, C.; van der Lee, T.; Proctor, R.H. The genetic basis for 3-ADON and 15-ADON trichothecene chemotypes in Fusarium. Fungal Genet. Biol. 2011, 48, 485–495. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Schmeitzl, C.; Varga, E.; Warth, B.; Kugler, K.G.; Malachová, A.; Michlmayr, H.; Wiesenberger, G.; Mayer, K.F.; Mewes, H.W.; Krska, R.; et al. Identification and Characterization of Carboxylesterases from Brachypodium distachyon Deacetylating Trichothecene Mycotoxins. Toxins 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Generotti, S.; Cirlini, M.; Malachova, A.; Sulyok, M.; Berthiller, F.; Dall’Asta, C.; Suman, M. Deoxynivalenol & Deoxynivalenol-3-Glucoside Mitigation through Bakery Production Strategies: Effective Experimental Design within Industrial Rusk-Making Technology. Toxins 2015, 7, 2773–2790. [Google Scholar] [PubMed]
- Wu, F.; Groopman, J.D.; Pestka, J.J. Public health impacts of foodborne mycotoxins. Annu. Rev. Food Sci. Technol. 2014, 5, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations; World Health Organization. Evaluation of Certain Contaminants in Food: Seventy-Second Report of the Joint FAO/WHO Expert Committee on Food Additives; WHO Technical Report Series 959; WHO: Geneva, Switzerland, 2011. [Google Scholar]
- European Food Safety Authority (EFSA) Panel on Contaminants in the Food Chain. Scientific opinion on risks for animal and public health related to the presence of nivalenol in food and feed. EFSA J. 2013, 11. [Google Scholar] [CrossRef]
- European Commission Regulation No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, L364, 5–24.
- Fruhmann, P.; Mikula, H.; Wiesenberger, G.; Varga, E.; Lumpi, D.; Stöger, B.; Häubl, G.; Lemmens, M.; Berthiller, F.; Krska, R.; et al. Isolation and Structure Elucidation of Pentahydroxyscirpene, a Trichothecene Fusarium Mycotoxin. J. Nat. Prod. 2014, 77, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Malachová, A.; Sulyok, M.; Beltrán, E.; Berthiller, F.; Krska, R. Optimization and validation of a quantitative liquid chromatography-tandem mass spectrometric method covering 295 bacterial and fungal metabolites including all regulated mycotoxins in four model food matrices. J. Chrom. A 2014, 1362, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Geiser, D.M.; del Mar Jiménez-Gasco, M.; Kang, S.; Makalowska, I.; Veeraraghavan, N.; Ward, T.J.; Zhang, N.; Kuldau, G.A.; O’Donnell, K. FUSARIUM-ID v. 1.0: A DNA Sequence Database for Identifying Fusarium. Eur. J. Plant Pathol. 2004, 110, 473–479. [Google Scholar] [CrossRef]
- Knutsen, A.K.; Torp, M.; Holst-Jensen, A. Phylogenetic analyses of the Fusarium poae, Fusarium sporotrichioides and Fusarium langsethiae species complex based on partial sequences of the translation elongation factor-1 alpha gene. Int. J. Food Microbiol. 2004, 95, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Fusarium ID database. Available online: http://isolate.fusariumdb.org/ (accessed on 6 July 2016).
- NCBI. Available online: https://blast.ncbi.nlm.nih.gov (accessed on 13 February 2015).
- Kim, J.-E.; Han, K.-H.; Jin, J.; Kim, H.; Kim, J.-C.; Yun, S.-H.; Lee, Y.-W. Putative Polyketide Synthase and Laccase Genes for Biosynthesis of Aurofusarin in Gibberella zeae. Appl. Environ. Microbiol. 2005, 71, 1701–1708. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Heydt, M.; Häckel, S.; Rüfer, C.E.; Geisen, R. A strain of Fusarium kyushuense is able to produce aflatoxin B1 and G1. Mycotoxin Res. 2009, 25, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Krska, R.; Schothorst, R.C.; van Egmond, H.P.; Josephs, R.D.; Lepschy, J.; Pettersson, H.; Chan, D.; Berthiller, F.; Schuhmacher, R.; Kandler, W.; et al. Processing and purity assessment of standards for the analysis of type-B trichothecene mycotoxins. Anal. Bioanal. Chem. 2005, 382, 1848–1858. [Google Scholar] [CrossRef] [PubMed]
- Steiner, B.; Kurz, H.; Lemmens, M.; Buerstmayr, H. Differential gene expression of related wheat lines with contrasting levels of head blight resistance after Fusarium graminearum inoculation. Theory Appl. Genet. 2009, 118, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Garreau de Loubresse, N.; Prokhorova, I.; Holtkamp, W.; Rodnina, M.V.; Yusupova, G.; Yusupov, M. Structural basis for the inhibition of the eukaryotic ribosome. Nature 2014, 513, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Hedman, R.; Pettersson, H. Purification and quantification of nivalenol. Mycotoxin Res. 1996, 12, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Tatsuno, T. Toxicologic Research on Substances from Fusarium nivale. Cancer Res. 1968, 28, 2393–2396. [Google Scholar] [PubMed]
- Nielsen, L.K.; Jensen, J.D.; Nielsen, G.C.; Jensen, J.E.; Spliid, N.H.; Thomsen, I.K.; Justesen, A.F.; Collinge, D.B.; Jørgensen, L.N. Fusarium head blight of cereals in Denmark: Species complex and related mycotoxins. Phytopathology 2011, 101, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.W.; Mirocha, C.J. Production of Nivalenol and Fusarenone-X by Fusarium tricinctum Fn-2B on a Rice Substrate. Appl. Environ. Micribiol. 1984, 48, 857–858. [Google Scholar]
- Marasas, W.F.O.; Nelson, P.E.; Toussoun, T.A. Toxigenic Fusarium Species—Identification and Mycotoxicology; Pennsylvania State University Press: University Park, PA, USA, 1984. [Google Scholar]
- Mule, G.; Logrieco, A.; Bottalico, A. Fusarium sp. FN-2B: A controversial strain genetically close to Fusarium poae. Mycotoxin Res. 1995, 11, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; O’Donnell, K. Fusarium kyushuense sp. nov. form Japan. Mycoscience 1998, 39. [Google Scholar] [CrossRef]
- Ohtsubo, K.; Ryu, J.C.; Nakamura, K.; Izumiyama, N.; Tanaka, T.; Yamamura, H.; Kobayashi, T.; Ueno, Y. Chronic toxicity of nivalenol in female mice: A 2-year feeding study with Fusarium nivale Fn-2B-moulded rice. Food Chem. Toxicol. 1989, 27, 591–598. [Google Scholar] [CrossRef]
- Ryu, J.C.; Ohtsubo, K.; Izumiyama, N.; Nakamura, K.; Tanaka, T.; Yamamura, H.; Ueno, Y. The acute and chronic toxicities of nivalenol in mice. Fundam. Appl. Toxicol. 1988, 11, 38–47. [Google Scholar] [CrossRef]
- Yamamura, H.; Kobayashi, T.; Ryu, J.C.; Ueno, Y.; Nakamura, K.; Izumiyama, N.; Ohtsubo, K. Subchronic feeding studies with nivalenol in C57BL/6 mice. Food Chem. Toxicol. 1989, 27, 585–590. [Google Scholar] [CrossRef]
- Meek, I.B.; Peplow, A.W.; Ake, C., Jr.; Phillips, T.D.; Beremand, M.N. Tri1 encodes the cytochrome P450 monooxygenase for C-8 hydroxylation during trichothecene biosynthesis in Fusarium sporotrichioides and resides upstream of another new Tri gene. Appl. Environ. Microbiol. 2003, 69, 1607–1613. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P.; Harris, L.J.; Alexander, N.J.; Ouellet, T.; Saparno, A.; Allard, S.; Desjardins, A.E. Tri1 in Fusarium graminearum Encodes a P450 Oxygenase. Appl. Environ. Microbiol. 2004, 70, 2044–2051. [Google Scholar] [CrossRef] [PubMed]
- Thrane, U.; Adler, A.; Clasen, P.E.; Galvano, F.; Langseth, W.; Lew, H.; Logrieco, A.; Nielsen, K.F.; Ritieni, A. Diversity in metabolite production by Fusarium langsethiae, Fusarium poae, and Fusarium sporotrichioides. Int. J. Food Microbiol. 2004, 95, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Vogelgsang, S.; Sulyok, M.; Bänzinger, I.; Krska, R.; Schuhmacher, R.; Forrer, H.R. Effect of fungal strain and cereal substrate on in vitro mycotoxin production by Fusarium poae and Fusarium avenaceum. Food Addit. Contam. Part A 2008, 25, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Fruhmann, P.; Weigl-Pollack, T.; Mikula, H.; Wiesenberger, G.; Adam, G.; Varga, E.; Berthiller, F.; Krska, R.; Hametner, C.; Fröhlich, J. Methylthiodeoxynivalenol (MTD): Insight into the chemistry, structure and toxicity of thia-Michael adducts of trichothecenes. Org. Biomol. Chem. 2014, 12, 5144–5150. [Google Scholar] [CrossRef] [PubMed]
- Stanic, A.; Uhlig, S.; Solhaug, A.; Rise, F.; Wilkins, A.L.; Miles, C.O. Nucleophilic Addition of Thiols to Deoxynivalenol. J. Agric. Food Chem. 2015, 63, 7556–7566. [Google Scholar] [CrossRef] [PubMed]
- Sulyok, M.; Berthiller, F.; Krska, R.; Schuhmacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Cigelnik, E.; Weber, N.S.; Trappe, J.M. Phylogenetic Relationships among Ascomycetous Truffles and the True and False Morels Inferred from 18S and 28S Ribosomal DNA Sequence Analysis. Mycologia 1997, 89, 48–65. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006; pp. 10–11. [Google Scholar]
- Warth, B.; Parich, A.; Bueschl, C.; Schoefbeck, D.; Neumann, N.K.N.; Kluger, B.; Schuster, K.; Krska, R.; Adam, G.; Lemmens, M.; et al. GC–MS based targeted metabolic profiling identifies changes in the wheat metabolome following deoxynivalenol treatment. Metabolomics 2015, 11, 722–738. [Google Scholar] [CrossRef] [PubMed]
- Varga, E.; Wiesenberger, G.; Hametner, C.; Ward, T.J.; Dong, Y.; Schöfbeck, D.; McCormick, S.; Broz, K.; Stückler, R.; Schuhmacher, R.; et al. New tricks of an old enemy: Isolates of Fusarium graminearum produce a type A trichothecence mycotoxin. Environ. Microbiol. 2015, 17, 2588–2600. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
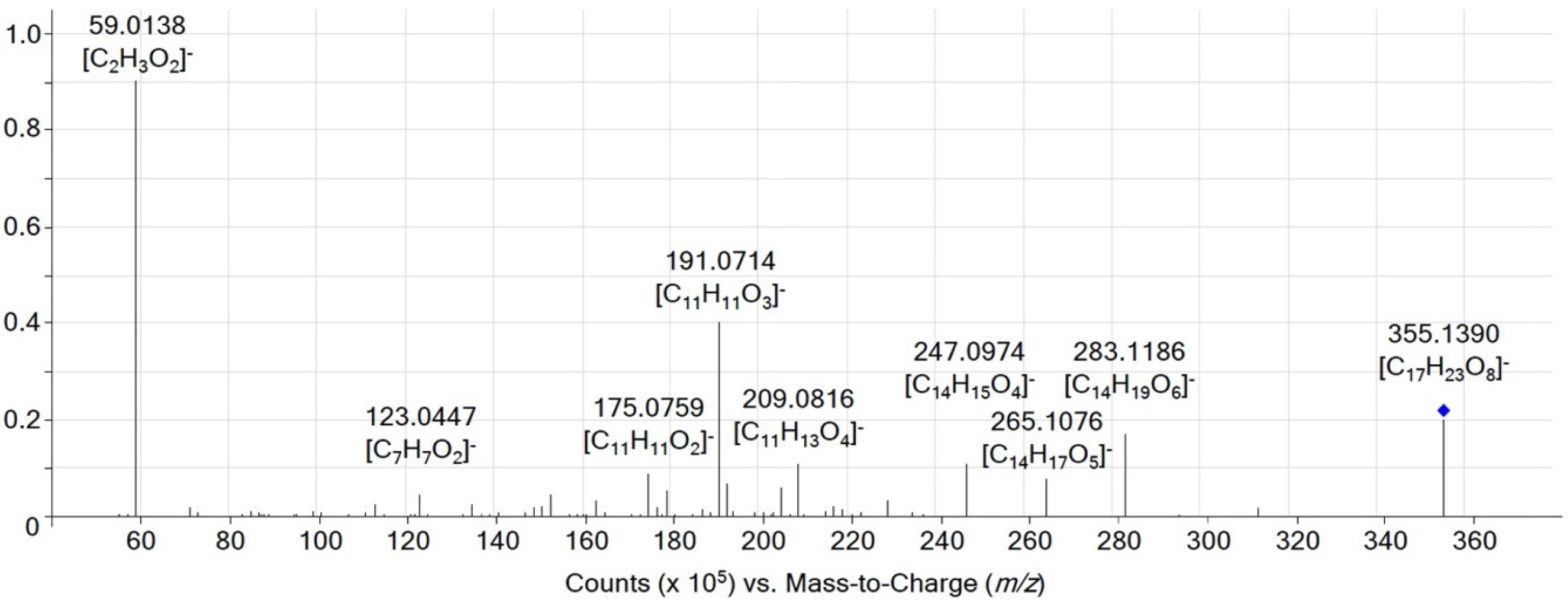{kind=link}
{kind=link}
| Analyte | Retention Time (min) | m/z Precursor Ion (Declustering Potential in V) | Product Ions (Collision Energy in eV) |
|---|---|---|---|
| Pentahydroxyscirpene | 4.0 | 313.1 (−110) | 163.1 (−39) |
| 175.0 (−30) | |||
| 191.1 (−21) | |||
| Nivalenol | 4.8 | 371.1 (−75) | 59.1 (−42) |
| 281.1 (−22) | |||
| Fusarenon X | 6.1 | 413.2 (−70) | 59.1 (−44) |
| 263.0 (−22) |
| Fusarium Strains | PHS (mg/kg) | NIV (mg/kg) | FUSX (mg/kg) |
|---|---|---|---|
| F. asiaticum SCK04 (#1) 1,2 | <0.15 | 4.84 | 67.0 |
| F. asiaticum SCK04 (#1, PDA) 3 | <0.15 | 5.24 | 42.4 |
| F. asiaticum SCK04 (#2) 2 | <0.15 | 2.65 | 12.1 |
| F. asiaticum SCK04 (#2, PDA) 3 | <0.15 | 3.26 | 20.8 |
| F. culmorum IFA450 | <0.15 | 0.15 | 0.74 |
| F. equiseti IFA33 | <0.15 | 0.23 | 0.86 |
| F. equiseti IFA34 | <0.15 | <0.012 | <0.04 |
| F. equiseti IFA63 | <0.15 | 0.07 | 0.16 |
| F. equiseti IFA64 | <0.15 | 0.28 | 1.23 |
| F. equiseti IFA157 | <0.15 | <0.012 | 0.05 |
| F. equiseti IFA336 | <0.15 | <0.012 | <0.04 |
| F. equiseti IFA408 | <0.15 | 0.036 | 0.15 |
| F. equiseti IFA409 | <0.15 | 0.10 | 0.33 |
| F. equiseti IFA410 | 0.23 | 60.0 | 110 |
| F. graminearum DAGZ5 | <0.15 | 0.67 | 20.0 |
| F. graminearum DAGZ8, gray mycelium 4 | 0.94 | 294 | 580 |
| F. graminearum DAGZ8, pink mycelium 4 | <0.15 | 5.00 | 212 |
| F. graminearum DAGZ13 | 0.17 | 212 | >6000 |
| F. graminearum DAGZ13 (PDA) 3 | 0.29 | 238 | >5000 |
| F. graminearum DAGZ22 | <0.15 | 0.37 | 43.6 |
| F. graminearum DAGZ23 | <0.15 | 2.71 | 49.6 |
| F. graminearum DAGZ24 | 0.49 | 88.8 | 788 |
| F. graminearum DAGZ25 | <0.15 | 0.10 | 4.84 |
| F. graminearum DAGZ29 | <0.15 | 10.3 | 242 |
| F. graminearum DAGZ31 | <0.15 | 44.0 | 107 |
| F. graminearum DAGZ36 | <0.15 | 0.25 | 21.9 |
| F. graminearum DAGZ37 | <0.15 | 1.91 | 33.4 |
| F. graminearum DAGZ39 | <0.15 | 0.14 | 1.68 |
| F. graminearum DAGZ46 | 1.44 | 399 | 1370 |
| F. graminearum DAGZ47 | <0.15 | 7.72 | 820 |
| F. graminearum DAGZ50 (#1) 2 | 1.65 | 199 | 283 |
| F. graminearum DAGZ50 (#2) 2 | 1.04 | 359 | >1000 |
| F. graminearum DAGZ50 (PDA) 3 | 2.93 | >900 | >3000 |
| F. graminearum DAGZ55 | <0.15 | 32.2 | 378 |
| F. graminearum DAGZ62 | <0.15 | 13.4 | 133 |
| F. graminearum DAGZ63 (#1) 2 | <0.15 | <0.012 | 16.0 |
| F. graminearum DAGZ63 (#1, PDA) 2,3 | <0.15 | 0.32 | 0.568 |
| F. graminearum DAGZ63 (#2) 2 | <0.15 | 0.06 | 0.94 |
| F. graminearum NRRL 26752 | <0.15 | 1.46 | 2.42 |
| F. graminearum NRRL 26752 (PDA) 3 | <0.15 | 1.26 | 1.48 |
| F. kyushuense IFA189 (#1) 2 | 110 | 1370 | 1980 |
| F. kyushuense IFA189 (#2) 2 | 22.7 | 536 | 4560 |
| F. kyushuense NRRL 3509 | <0.15 | <0.012 | <0.04 |
| F. kyushuense NRRL 3510 | <0.15 | 0.03 | 0.11 |
| F. kyushuense NRRL 6490 (=Fn-2B) | 10.0 | 404 | 2890 |
| F. kyushuense NRRL 6491 | 3.57 | 468 | 2930 |
| F. kyushuense NRRL 25348 (backup of 6490) | 15.0 | 328 | 1440 |
| F. kyushuense NRRL 26204 | <0.15 | 0.07 | 0.15 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varga, E.; Wiesenberger, G.; Fruhmann, P.; Malachová, A.; Svoboda, T.; Lemmens, M.; Adam, G.; Berthiller, F. Pentahydroxyscirpene—Producing Strains, Formation In Planta, and Natural Occurrence. Toxins 2016, 8, 295. https://doi.org/10.3390/toxins8100295
Varga E, Wiesenberger G, Fruhmann P, Malachová A, Svoboda T, Lemmens M, Adam G, Berthiller F. Pentahydroxyscirpene—Producing Strains, Formation In Planta, and Natural Occurrence. Toxins. 2016; 8(10):295. https://doi.org/10.3390/toxins8100295
Chicago/Turabian StyleVarga, Elisabeth, Gerlinde Wiesenberger, Philipp Fruhmann, Alexandra Malachová, Thomas Svoboda, Marc Lemmens, Gerhard Adam, and Franz Berthiller. 2016. "Pentahydroxyscirpene—Producing Strains, Formation In Planta, and Natural Occurrence" Toxins 8, no. 10: 295. https://doi.org/10.3390/toxins8100295
APA StyleVarga, E., Wiesenberger, G., Fruhmann, P., Malachová, A., Svoboda, T., Lemmens, M., Adam, G., & Berthiller, F. (2016). Pentahydroxyscirpene—Producing Strains, Formation In Planta, and Natural Occurrence. Toxins, 8(10), 295. https://doi.org/10.3390/toxins8100295