
Ochratoxin A: Molecular Interactions, Mechanisms of Toxicity and Prevention at the Molecular Level



Abstract
:
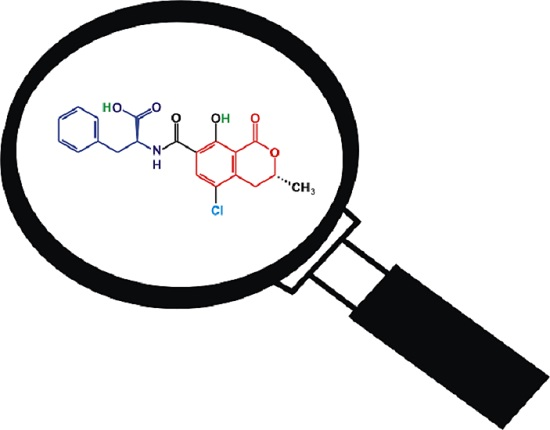{kind=link}
{kind=link}
{kind=link}
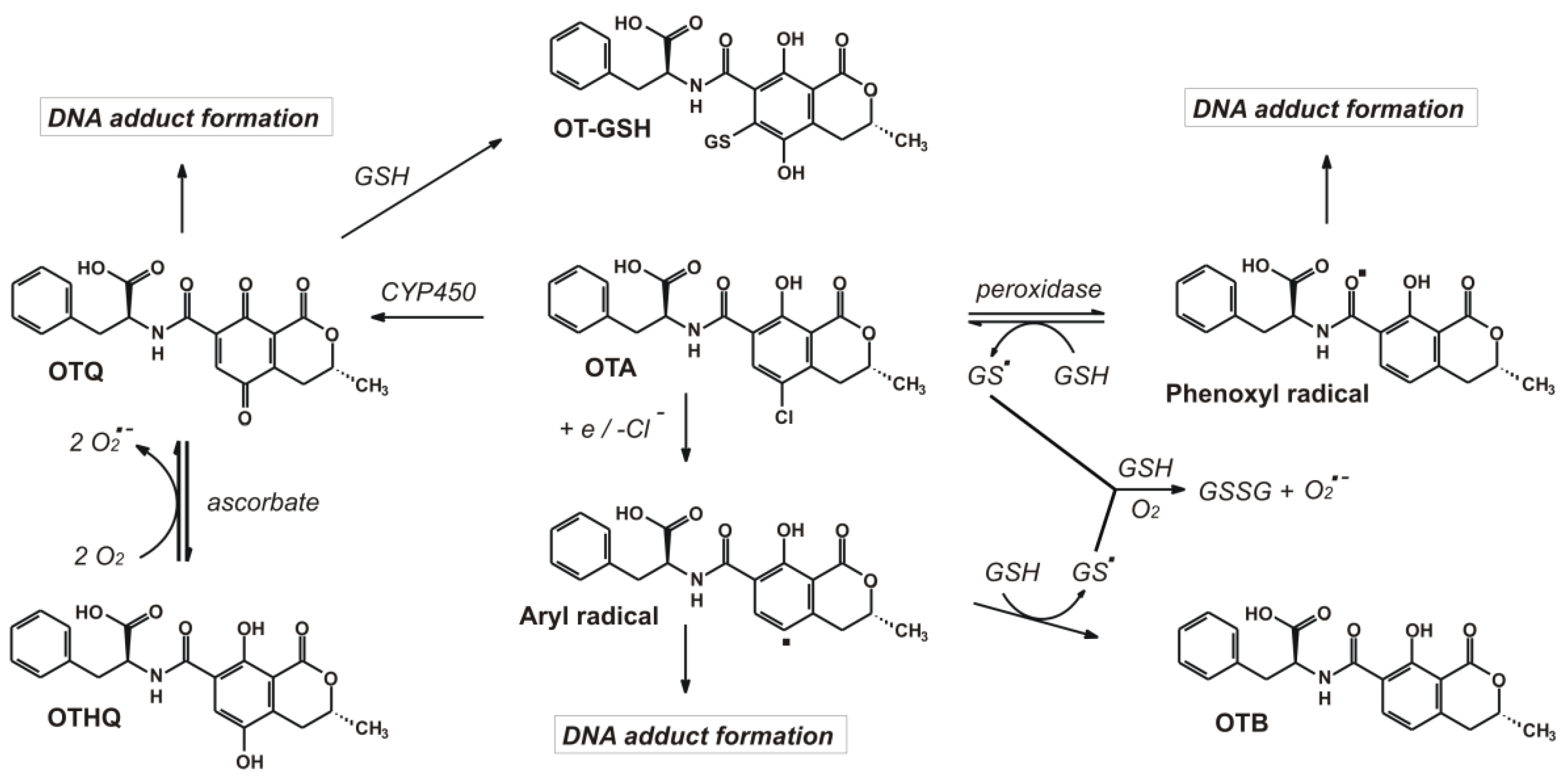{kind=link}
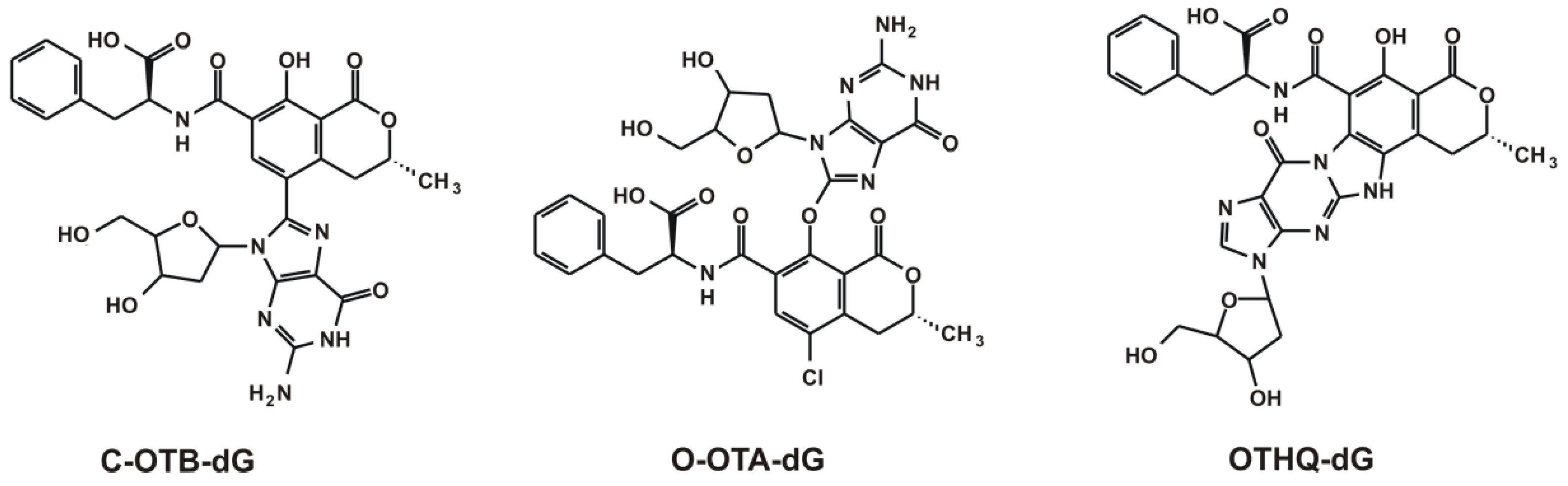{kind=link}
1. Introduction
2. Chemistry of Ochratoxins
3. Dietary Sources of OTA
4. Toxicokinetics of OTA
4.1. Absorption
4.2. Distribution
4.2.1. Binding to Plasma Proteins
4.2.2. Tissue Distribution
4.3. Biotransformation
4.4. Excretion
4.4.1. Renal Excretion
4.4.2. Fecal Excretion and Entero-Hepatic Circulation
4.4.3. Excretion through Breast Milk
5. Mode of Action of OTA
5.1. Inhibition of Protein Synthesis
5.2. Inhibition of Cellular Energy Production
5.3. Genotoxic Effect
5.4. Induction of Oxidative/Nitrosative Stress
5.5. Apoptosis
5.6. Influence on Mitosis
5.7. Induction of Cell Cycle Arrest
5.8. Other Mechanisms
6. Protective Agents—Overview
7. Displacement of OTA from Albumin
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ATP | Adenosine triphosphate |
| BCRP | Breast Cancer Resistance Protein |
| BEN | Balkan Endemic Nephropathy |
| C8-dG | C8-deoxyguanosine DNA-adduct |
| CYP450 | Cytochrom P450 enzyme family |
| DDAH | Dimethylarginine dimethylaminohydrolase |
| GSH | Glutathione (reduced) |
| GSSG | Glutathione (oxidized) |
| HSA | Human serum albumin |
| iNOS | Inducible Nitrogen Oxide Synthase |
| MRP2 | Multidrug Resistance-associated Protein 2 |
| NAC | N-acetyl cysteine |
| Nrf2 | Nuclear factor-erythroid 2 p45-related factor 2 |
| OAT | Organic Anion Transporter |
| OATP | Organic Anion-transporting Polypeptide |
| OTα | Ochratoxin α |
| OTA | Ochratoxin A |
| OTB | Ochratoxin B |
| OTC | Ochratoxin C |
| OTHQ | OTA-hydroquinone |
| OTQ | OTA-quinone |
| PEPCK | Phosphoenolpyruvate carboxykinase |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
References
- Ringot, D.; Chango, A.; Schneider, Y.J.; Larondelle, Y. Toxicokinetics and toxicodynamics of Ochratoxin A, an update. Chem. Biol. Interact. 2006, 159, 18–46. [Google Scholar] [CrossRef] [PubMed]
- Van Der Merwe, K.J.; Steyn, P.S.; Fourie, L. Part II. The constitution of Ochratoxins A, B, and C, metabolites of Aspergillus ochraceus wilh. J. Chem. Soc. 1965, 7083–7088. [Google Scholar] [CrossRef]
- Pohland, A.E.; Nesheim, S.; Friedman, L. Ochratoxin A: A review. Pure Appl. Chem. 1992, 64, 1029–1046. [Google Scholar] [CrossRef]
- Speijers, G.J.A.; van Egmond, H.P. Worldwide Ochratoxin A levels in food and feeds. In Human Ochratoxicosis and its Pathologies; Creppy, E.E., Castegnaro, M., Dirheimer, G., Eds.; John Libbey Eurotext: London, UK, 1993; Vol. 231, pp. 85–100. [Google Scholar]
- Krogh, P.; Hald, B.; Plestina, R.; Ceović, S. Balkan (endemic) nephropathy and foodborn Ochratoxin A: Preliminary results of a survey of foodstuffs. Acta Pathol. Microbiol. Scand. B 1977, 85, 238–240. [Google Scholar] [CrossRef] [PubMed]
- Pavlovic, M.; Plestina, R.; Krogh, P. Ochratoxin A contamination of foodstuffs in an area with Balkan (endemic) nephropathy. Acta Pathol. Microbiol. Scand. B 1979, 87, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Petkova-Bocharova, T.; Castegnaro, M. Ochratoxin A contamination of cereals in an area of high incidence of Balkan endemic nephropathy in Bulgaria. Food Addit. Contam. 1985, 2, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Petkova-Bocharova, T.; Castegnaro, M.; Michelon, J.; Maru, V. Ochratoxin A and other mycotoxins in cereals from an area of Balkan endemic nephropathy and urinary tract tumours in Bulgaria. In Mycotoxins, Endemic Nephropathy and Urinary Tract Tumours; Castegnaro, M., Plestina, R., Dirheimer, G., Chernozemsky, I.N., Bartsch, H., Eds.; IARC Scientific Publications: Lyon, France, 1991; pp. 83–87. [Google Scholar]
- Pfohl-Leszkowicz, A.; Petkova-Bocharova, T.; Chernozemsky, I.N.; Castegnaro, M. Balkan endemic nephropathy and associated urinary tract tumours: A review on aetiological causes and the potential role of mycotoxins. Food Addit. Contam. 2002, 19, 282–302. [Google Scholar] [CrossRef] [PubMed]
- Vrabcheva, T.; Usleber, E.; Dietrich, R.; Martlbauer, E. Co-occurrence of Ochratoxin A and citrinin in cereals from Bulgarian villages with a history of Balkan endemic nephropathy. J. Agric. Food Chem. 2000, 48, 2483–2488. [Google Scholar] [CrossRef] [PubMed]
- Bui-Klimke, T.; Wu, F. Evaluating weight of evidence in the mystery of balkan endemic nephropathy. Risk Anal. 2014, 34, 1688–1705. [Google Scholar] [CrossRef] [PubMed]
- Pavlović, N.M. Balkan endemic nephropathy—Current status and future perspectives. Clin. Kidney J. 2013, 6, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Markovic, B.; Lebedev, S.; Djordjevic, M.; Arambasic, M. Endemic urinary tract cancer in Bulgaria, Yugoslavia and Romania: Etiology and pathogenesis. Med. Biol. Environ. 1976, 1–2. [Google Scholar]
- Chernozemsky, I.N.; Stoyanov, I.S.; Petkova-Bocharova, T.K.; Nicolov, I.G.; Draganov, I.V.; Stoichev, I.I.; Tanchev, Y.; Naidenov, D.; Kalcheva, N.D. Geographic correlation between the occurrence of endemic nephropathy and urinary tract tumours in vratza district, Bulgaria. Int. J. Cancer 1977, 19, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pfohl-Leszkowicz, A. Ochratoxin A and aristolochic acid involvement in nephropathies and associated urothelial tract tumours. Arh. Hig. Rada Toksikol. 2009, 60, 465–483. [Google Scholar] [CrossRef] [PubMed]
- Duarte, S.C.; Pena, A.; Lino, C.M. Ochratoxin A non-conventional exposure sources—A review. Microchem. J. 2009, 93, 115–120. [Google Scholar] [CrossRef]
- Amézqueta, S.; González-Peñas, E.; Murillo-Arbizu, M.; López de Cerain, A. Ochratoxin A decontamination: A review. Food Control 2009, 20, 326–333. [Google Scholar] [CrossRef]
- Ciconová, P.; Laciková, A.; Máté, D. Prevention of Ochratoxin A contamination of food and Ochratoxin A detoxification by microorganisms—A review. Czech J. Food Sci. 2010, 28, 465–474. [Google Scholar]
- Pfohl-Leszkowicz, A.; Hadjeba-Medjdoub, K.; Ballet, N.; Schrickx, J.; Fink-Gremmels, J. Assessment and characterization of yeast-based products intended to mitigate ochratoxin exposure using in vitro and in vivo models. Food Addit. Contam. A 2015, 32, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Radic, B.; Fuchs, R.; Peraica, M.; Lucic, A. Ochratloxin A in human sera in the area with endemic nephropathy in Croatia. Toxicol. Lett. 1997, 91, 105–109. [Google Scholar] [CrossRef]
- Peraica, M.; Domijan, A.M.; Fuchs, R.; Lucic, A.; Radic, B. The occurrence of Ochratoxin A in blood in general population of Croatia. Toxicol. Lett. 1999, 110, 105–112. [Google Scholar] [CrossRef]
- Özcelik, N.; Kosar, A.; Soysal, D. Ochratoxin A in human serum samples collected in Isparta-Turkey from healthy individuals and individuals suffering from different urinary disorders. Toxicol. Lett. 2001, 121, 9–13. [Google Scholar] [CrossRef]
- Lino, C.M.; Baeta, M.L.; Henri, M.; Dinis, A.M.P.; Pena, A.S.; Silveira, M.I.N. Levels of Ochratoxin A in serum from urban and rural Portuguese populations and estimation of exposure degree. Food Chem. Toxicol. 2008, 46, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.L.; Christensen, T.; Goldsmith, M.R.; Toone, E.J.; Beratan, D.N.; Simon, J.D. Binding of Ochratoxin A to human serum albumin stabilized by a protein-ligand ion pair. J. Phys. Chem. B 2003, 107, 7884–7888. [Google Scholar] [CrossRef]
- Poór, M.; Kunsági-Máté, S.; Bencsik, T.; Petrik, J.; Vladimir-Knežević, S.; Kőszegi, T. Flavonoid aglycones can compete with Ochratoxin A for human serum albumin: A new possible mode of action. Int. J. Biol. Macromol. 2012, 51, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Poór, M.; Kunsági-Máté, S.; Matisz, G.; Li, Y.; Czibulya, Z.; Peles-Lemli, B.; Kőszegi, T. Interaction of alkali and alkaline earth ions with Ochratoxin A. J. Lumin. 2013, 135, 276–280. [Google Scholar]
- Hagelberg, S.; Hult, K.; Fuchs, R. Toxicokinetics of Ochratoxin A in several species and its plasma-binding properties. J. Appl. Toxicol. 1989, 9, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Studer-Rohr, I.; Schlatter, J.; Dietrich, D.R. Kinetic parameters and intraindividual fluctuations of Ochratoxin A plasma levels in human. Arch. Toxicol. 2000, 74, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, S. Ochratoxin A: Plasma concentration and excretion into bile and urine in albumin-deficient rats. Food Chem. Toxicol. 1985, 23, 941–943. [Google Scholar] [CrossRef]
- Poór, M.; Kunsági-Máté, S.; Czibulya, Z.; Li, Y.; Peles-Lemli, B.; Petrik, J.; Vladimir-Knežević, S.; Kőszegi, T. Fluorescence spectroscopic investigation of competitive interactions between Ochratoxin A and 13 drug molecules for binding to human serum albumin. Luminescence 2013, 28, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Poór, M.; Li, Y.; Matisz, G.; Kiss, L.; Kunsági-Máté, S.; Kőszegi, T. Quantitation of species differences in albumin-ligand interactions for bovine, human and rat serum albumins using fluorescence spectroscopy: A test case with some Sudlow's site I ligands. J. Lumin. 2014, 145, 767–773. [Google Scholar]
- Li, Y.; Czibulya, Z.; Poór, M.; Lecomte, S.; Kiss, L.; Harte, E.; Kőszegi, T.; Kunsági-Máté, S. Thermodynamic study of the effects of ethanol on the interaction of Ochratoxin A with human serum albumin. J. Lumin. 2014, 148, 18–25. [Google Scholar] [CrossRef]
- Castegnaro, M.; Canadas, D.; Vrabcheva, T.; Petkova-Bocharova, T.; Chernozemsky, I.N.; Pfohl-Leszkowicz, A. Balkan endemic nephropathy: Role of Ochratoxins A through biomarkers. Mol. Nutr. Food Res. 2006, 50, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Pfohl-Leszkowicz, A.; Manderville, R.A. Ochratoxin A: An overview on toxicity and carcinogenicity in animals and humans. Mol. Nutr. Food Res. 2007, 51, 61–99. [Google Scholar] [CrossRef] [PubMed]
- Manderville, R.A. A case for the genotoxicity of Ochratoxin A by bioactivation and covalent DNA adduction. Chem. Res. Toxicol. 2005, 18, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Turesky, R.J. Perspective: Ochratoxin A is not a genotoxic carcinogen. Chem. Res. Toxicol. 2005, 18, 1082–1090. [Google Scholar] [CrossRef] [PubMed]
- Manderville, R.; Pfohl-Leszkowicz, A. Bioactivation and DNA adduction as a rationale for Ochratoxin A carcinogenesis. World Mycotoxin J. 2008, 1, 357–367. [Google Scholar] [CrossRef]
- Pfohl-Leszkowicz, A.; Manderville, R.A. An update on direct genotoxicity as a molecular mechanism of Ochratoxin A carcinogenicity. Chem. Res. Toxicol. 2012, 25, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Hadjeba-Medjdoub, K.; Tozlovanu, M.; Pfohl-Leszkowicz, A.; Frenette, C.; Paugh, R.J.; Manderville, R.A. Structure-activity relationships imply different mechanisms of action for Ochratoxin A-mediated cytotoxicity and genotoxicity. Chem. Res. Toxicol. 2012, 25, 181–190. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, A.; Atoui, A. Ochratoxin A: General overview and actual molecular status. Toxins 2010, 2, 461–493. [Google Scholar] [CrossRef] [PubMed]
- IARC. Ochratoxin A. Monographs on the Evaluation of Carcinogenic Risks to Humans. In Some Naturally Occurring Substances: Food Items and Constituents, Heterocyclic Aromatic Amines and Mycotoxins; International Agency for Research on Cancer: Lyon, France, 1993; pp. 489–521. [Google Scholar]
- Boorman, G. NTP Technical Report on the Toxicology and Carcinogenesis Studies of Ochratoxin A (CAS No. 303-47-9) in F344/N Rats (Gavage Studies); NIH Publication No. 89-2813; U.S. Department of Health and Human Services, National Institutes of Health: Research Triangle Park, NC, USA, 1989.
- Chernozemsky, I.N. Balkan endemic nephropathy and the associated tumours of the urinary system: A summary of epidemiological features in Bulgaria. IARC Sci. Publ. 1991, 115, 3–4. [Google Scholar] [PubMed]
- Hassen, W.; Abid-Essafi, S.; Achour, A.; Guezzah, N.; Zakhama, A.; Ellouz, F.; Creppy, E.E.; Bacha, H. Karyomegaly of tubular kidney cells in human chronic interstitial nephropathy in Tunisia: Respective role of Ochratoxin A and possible genetic predisposition. Hum. Exp. Toxicol. 2004, 23, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Xing, L.; Li, Z.; Wu, S.; Wang, J.; Liu, J.; Wang, J.; Yan, X.; Zhang, X. Ochratoxin A induces G(2) phase arrest in human gastric epithelium GES-1 cells in vitro. Toxicol. Lett. 2010, 193, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, S.; Shen, H.; Cui, J.; Wang, Y.; Xing, L.; Wang, J.; Yan, X.; Zhang, X. Ochratoxin A induces DNA damage and G2 phase arrest in human esophageal epithelium Het-1A cells in vitro. J. Toxicol. Sci. 2015, 40, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, G.G. Hypothesis: Does Ochratoxin A cause testicular cancer? Cancer Causes Control 2002, 13, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Pohland, A.E.; Schuller, P.L.; Steyn, P.S. Physicochemical data for some selected mycotoxins. Pure Appl. Chem. 1982, 54, 2219–2284. [Google Scholar] [CrossRef]
- Vidal, A.; Sanchis, V.; Ramos, A.J.; Marín, S. Thermal stability and kinetics of degradation of deoxynivalenol, deoxynivalenol conjugates and Ochratoxin A during baking of wheat bakery products. Food Chem. 2015, 178, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, A.B.; Doi, E.; Kitabatake, N. Detoxification of Ochratoxin A on heating under acidic and alkaline conditions. Biosci. Biotech. Biochem. 1992, 56, 741–745. [Google Scholar] [CrossRef]
- Boudra, H.; Le Bars, P.; Le Bars, J. Thermostability of Ochratoxin A in wheat under two moisture conditions. Appl. Environ. Microbiol. 1995, 61, 1156–1158. [Google Scholar] [PubMed]
- Blanc, M.; Pittet, A.; Munoz-Box, R.; Viani, R. Behavior of Ochratoxin A during green coffee roasting and soluble coffee manufacture. J. Agric. Food Chem. 1998, 46, 673–675. [Google Scholar] [CrossRef] [PubMed]
- Van der Stegen, G.H.; Essens, P.J.; van der Lijn, J. Effect of roasting conditions on reduction of Ochratoxin A in coffee. J. Agric. Food Chem. 2001, 49, 4713–4715. [Google Scholar] [CrossRef] [PubMed]
- Cramer, B.; Königs, M.; Humpf, H.U. Identification and in vitro cytotoxicity of Ochratoxin A degradation products formed during coffee roasting. J. Agric. Food Chem. 2008, 56, 5673–5681. [Google Scholar] [CrossRef] [PubMed]
- Bellver Soto, J.; Fernández-Franzón, M.; Ruiz, M.J.; Juan-García, A. Presence of Ochratoxin A (OTA) mycotoxin in alcoholic drinks from southern European countries: Wine and beer. J. Agric. Food Chem. 2014, 62, 7643–7651. [Google Scholar] [CrossRef] [PubMed]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Toman, J.; Bazin, I.; Roubal, T. Transfer of Ochratoxin A into tea and coffee beverages. Toxins 2014, 6, 3438–3453. [Google Scholar] [CrossRef] [PubMed]
- Flores-Flores, M.E.; Lizarraga, E.; López de Cerain, A.; González-Penas, E. Presence of mycotoxins in animal milk: A review. Food Control 2015, 53, 163–176. [Google Scholar] [CrossRef]
- Shim, W.B.; Ha, K.S.; Kim, M.G.; Kim, J.S.; Chung, D.H. Evaluation of the transfer rate of Ochratoxin A to decoctions of herbal medicines. Food Sci. Biotechnol. 2014, 23, 2103–2108. [Google Scholar] [CrossRef]
- Chen, A.J.; Jiao, X.; Hu, Y.; Lu, X.; Gao, W. Mycobiota and Mycotoxins in traditional medicinal seeds from China. Toxins 2015, 7, 3858–3875. [Google Scholar] [CrossRef] [PubMed]
- Veprikova, Z.; Zachariasova, M.; Dzuman, Z.; Zachariasova, A.; Fenclova, M.; Slavikova, P.; Vaclavikova, M.; Mastovska, K.; Hengst, D.; Hajslova, J. Mycotoxins in plant-based dietary supplements: Hidden health risk for consumers. J. Agric. Food Chem. 2015, 63, 6633–6643. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzo, M.; Piemontese, L.; Gambacorta, L.; Zivoli, R.; Longobardi, F. Food coloring agents and plant food supplements derived from Vitis vinifera: A new source of human exposure to Ochratoxin A. J. Agric. Food Chem. 2015, 63, 3609–3614. [Google Scholar] [CrossRef] [PubMed]
- Ostry, V.; Malir, F.; Ruprich, J. Producers and important dietary sources of Ochratoxin A and citrinin. Toxins 2013, 5, 1574–1586. [Google Scholar] [CrossRef] [PubMed]
- Jeswal, P.; Kumar, D. Mycobiota and natural incidence of Aflatoxins, Ochratoxin A, and Citrinin in Indian spices confirmed by LC-MS/MS. Int. J. Microbiol. 2015, 2015, 242486. [Google Scholar] [CrossRef] [PubMed]
- Do, K.H.; An, T.J.; Oh, S.K.; Moon, Y. Nation-based occurrence and endogenous biological reduction of mycotoxins in medicinal herbs and spices. Toxins 2015, 7, 4111–4130. [Google Scholar] [CrossRef] [PubMed]
- Mata, A.T.; Ferreira, J.P.; Oliveira, B.R.; Batoréu, M.C.; Barreto Crespo, M.T.; Pereira, V.J.; Bronze, M.R. Bottled water: Analysis of mycotoxins by LC-MS/MS. Food Chem. 2015, 176, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Galtier, P.; Alvinerie, M.; Charpenteau, J.L. The pharmacokinetic profiles of Ochratoxin A in pigs, rabbits and chickens. Food Cosmet. Toxicol. 1981, 19, 735–738. [Google Scholar] [CrossRef]
- Galtier, P. Contribution of pharmacokinetic studies to mycotoxicology-Ochratoxin A. Vet. Sci. Commun. 1978, 1, 349–358. [Google Scholar] [CrossRef]
- Kumagai, S.; Aibara, K. Intestinal absorption and secretion of Ochratoxin A in the rat. Toxicol. Appl. Pharmacol. 1982, 64, 94–102. [Google Scholar] [CrossRef]
- Roth, A.; Chakor, K.; Creppy, E.E.; Kane, A.; Roschenthaler, R.; Dirheimer, G. Evidence for an enterohepatic circulation of Ochratoxin A in mice. Toxicology 1988, 48, 293–308. [Google Scholar] [CrossRef]
- Berger, V.; Gabriel, A.F.; Sergent, T.; Trouet, A.; Larondelle, Y.; Schneider, Y.J. Interaction of Ochratoxin A with human intestinal Caco-2 cells: Possible implication of a multidrug resistance-associated protein (MRP2). Toxicol. Lett. 2003, 140–141, 465–476. [Google Scholar] [CrossRef]
- Schrickx, J.; Lektarau, Y.; Fink-Gremmels, J. Ochratoxin A secretion by ATP-dependent membrane transporters in Caco-2 cells. Arch. Toxicol. 2006, 80, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M.; Mahfoud, R.; Pfohl-Leszkowicz, A.; Fantini, J. The mycotoxin Ochratoxin A alters intestinal barrier and absorption functions but has no effect on chloride secretion. Toxicol. Appl. Pharmacol. 2001, 176, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Il’ichev, Y.V.; Perry, J.L.; Simon, J.D. Interaction of Ochratoxin A with human serum albumin. Preferential binding of the dianion and pH effects. J. Phys. Chem. B 2002, 106, 452–459. [Google Scholar] [CrossRef]
- Il’ichev, Y.V.; Perry, J.L.; Rüker, F.; Dockal, M.; Simon, J.D. Interaction of Ochratoxin A with human serum albumin. Binding sites localized by competitive interactions with the native protein and its recombinant fragments. Chem. Biol. Interact. 2002, 141, 275–293. [Google Scholar] [CrossRef]
- Perry, J.L.; Goldsmith, M.R.; Peterson, M.A.; Beratan, D.N. Structure of the Ochratoxin A binding site within human serum albumin. J. Phys. Chem. B 2004, 108, 16960–16964. [Google Scholar] [CrossRef]
- Dai, J.; Park, G.; Perry, J.L.; Il'ichev, Y.V.; Bow, D.A.; Pritchard, J.B.; Faucet, V.; Pfohl-Leszkowicz, A.; Manderville, R.A.; Simon, J.D. Molecular aspects of the transport and toxicity of Ochratoxin A. Acc. Chem. Res. 2004, 37, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Il’ichev, Y.V.; Perry, J.L.; Simon, J.D. Interaction of Ochratoxin A with human serum albumin. A common binding site of Ochratoxin A and warfarin in subdomain IIA. J. Phys. Chem. B 2002, 106, 460–465. [Google Scholar] [CrossRef]
- Stojković, R.; Hult, K.; Gamulin, S.; Plestina, R. High affinity binding of Ochratoxin A to plasma constituents. Biochem. Int. 1984, 9, 33–38. [Google Scholar] [PubMed]
- Schwerdt, G.; Freudinger, R.; Silbernagl, S.; Gekle, M. Ochratoxin A—binding proteins in rat organs and plasma and in different cell lines of the kidney. Toxicology 1999, 135, 1–10. [Google Scholar] [CrossRef]
- Belmadani, A.; Tramu, G.; Betbeder, A.M.; Steyn, P.S.; Creppy, E.E. Regional selectivity to Ochratoxin A, distribution and cytotoxicity in rat brain. Arch. Toxicol. 1998, 72, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.Y.; Takeda, M.; Kim, D.K.; Tojo, A.; Narikawa, S.; Yoo, B.S.; Hosoyamada, M.; Cha, S.H.; Sekine, T.; Endou, H. Characterization of Ochratoxin A transport by human organic anion transporters. Life Sci. 2001, 69, 2123–2135. [Google Scholar] [CrossRef]
- Kontaxi, M.; Echkardt, U.; Hagenbuch, B.; Stieger, B.; Meier, P.J.; Petzinger, E. Uptake of the mycotoxin Ochratoxin A in liver cells occurs via the cloned organic anion transporting polypeptide. J. Pharmacol. Exp. Ther. 1996, 279, 1507–1513. [Google Scholar] [PubMed]
- Anzai, N.; Jutabha, P.; Endou, H. Molecular mechanism of Ochratoxin A transport in the kidney. Toxins 2010, 2, 1381–1398. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, A.; Masuda, S.; Saito, H.; Abe, T.; Inui, K. Multispecific substrate recognition of kidney-specific organic anion transporters OAT-K1 and OAT-K2. J. Pharmacol. Exp. Ther. 2001, 299, 261–267. [Google Scholar] [PubMed]
- Zlender, V.; Breljak, D.; Ljubojević, M.; Flajs, D.; Balen, D.; Brzica, H.; Domijan, A.M.; Peraica, M.; Fuchs, R.; Anzai, N.; et al. Low doses of Ochratoxin A upregulate the protein expression of organic anion transporters Oat1, Oat2, Oat3 and Oat5 in rat kidney cortex. Toxicol. Appl. Pharmacol. 2009, 239, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Miraglia, M.; Brera, C.; Corneli, S.; Cava, E.; Montanino, G.; Miraglia, E. Occurence of Ochratoxin A (OTA) in Maternal Serum, Placenta and Funiculum. In Proceedings of the IX International IUPAC Symposium on Mycotoxins and Phycotoxins—Developments in Chemistry, Toxicology and Food Safety, Rome, Italy, 27–31 May 1996; Miraglia, M., van Egmond, H., Brera, C., Gilbert, J., Eds.; Alaken Inc.: Fort Collins, CO, USA; pp. 165–179.
- Cha, S.H.; Sekine, T.; Kusushura, H.; Yu, E.; Kim, J.Y.; Kim, D.K.; Sugiyama, Y.; Kanai, Y.; Endou, H. Molecular cloning and characterization of multispecific organic anion transporter 4 expressed in the placenta. J. Biol. Chem. 2000, 275, 4507–4512. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; Bingle, L.E. Comparative Ochratoxin toxicity: A review of the available data. Toxins 2015, 7, 4253–4282. [Google Scholar] [CrossRef] [PubMed]
- Pitout, M.J. The hydrolysis of Ochratoxin A by some proteolytic enzymes. Biochem. Pharmacol. 1969, 18, 485–491. [Google Scholar] [CrossRef]
- Doster, R.C.; Sinnhuber, R.O. Comparative rate of hydrolysis of Ochratoxin A and B in vitro. Food Cosmet. Toxicol. 1972, 10, 389–394. [Google Scholar] [CrossRef]
- Hoehler, D.; Sudekum, K.H.; Wolfram, S.; Frohlich, A.A.; Marquardt, R.R. Metabolism and excretion of Ochratoxin A fed to sheep. J. Anim. Sci. 1999, 77, 1217–1223. [Google Scholar]
- Madhyastha, M.S.; Marquardt, R.R.; Frohlich, A.A. Hydrolysis of Ochratoxin A by the microbial activity of digesta in the gastrointestinal tract. Arch. Environ. Contam. Toxicol. 1992, 23, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Madhyastha, S.; Marquardt, R.R.; Li, S.; Vodela, J.K.; Frohlich, A.A.; Kemppainen, W. Toxicity of OTA, its opened lactone form and several of its analogs: Structure-activity relationships. Toxicol. Appl. Pharmacol. 1996, 137, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Stormer, F.C.; Pederson, J.I. Formation of (4R)- and (4S)-hydroxyochratoxin A from Ochratoxin A by rat liver microsomes. Appl. Environ. Microbiol. 1980, 39, 971–975. [Google Scholar] [PubMed]
- Stormer, F.C.; Storen, O.; Hansen, C.E.; Pedersen, J.I.; Hvistendahl, G.; Aarsen, A.J. Formation of (4R)- and (4S)-hydroxyochratoxin A from Ochratoxin A by liver microsomes from various species. Appl. Environ. Microbiol. 1981, 42, 1051–1056. [Google Scholar] [PubMed]
- Oster, T.; Jayyosi, Z.; Creppy, E.E.; El Amri, H.S.; Batt, A.M. Characterization of pig liver purified cytochrome P-450 isoenzymes for Ochratoxin A metabolism studies. Toxicol. Lett. 1991, 57, 203–214. [Google Scholar] [CrossRef]
- Hutchinson, R.D.; Steyn, P.S.; Thompson, D.L. The isolation and structure of 4-hydroxyochratoxin A and 7-carboxy-3,4-dihydro-8-hydroxy-3-methylisocoumarin from Penicillium viridicatum. Tetrahedron Lett. 1971, 43, 4033–4036. [Google Scholar] [CrossRef]
- Creppy, E.E.; Kern, D.; Steyn, P.S.; Vleggaar, R.; Roschenthaler, R.; Dirheimer, G. Comparative study on the effect of Ochratoxin A analogues on yeast aminoacyl-t-RNA synthetases and on the growth and protein synthesis of hepatoma cells. Toxicol. Lett. 1983, 19, 217–224. [Google Scholar] [CrossRef]
- Xiao, H.; Marquardt, R.R.; Abramson, R.R.; Frohlich, A.A. Metabolites of ochratoxins in rat urine and in a culture of Aspergillus ochraceus. Appl. Environ. Microbiol. 1996, 62, 648–655. [Google Scholar] [PubMed]
- Zepnik, H.; Pahler, A.; Schauer, U.; Dekant, W. Ochratoxin A-Induced tumor formation: Is there a role of reactive Ochratoxin A metabolites? Toxicol. Sci. 2001, 59, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Pinelli, E.; El Adlouni, C.; Pipy, B.; Quartulli, F.; Pfohl-Leskzowicz, A. Roles of cyclooxygenase and lipoygenases in Ochratoxin A genotoxicity in human epithelial lung cells. Environ. Toxicol. Pharmacol. 1999, 7, 95–107. [Google Scholar] [CrossRef]
- El Adlouni, C.; Pinelli, E.; Azemar, B.; Zaoui, D.; Beaune, P.; Leszkowicz, A.P. Phenobarbital increases of DNA adduct and metabolites formed by Ochratoxin A: Role of CYP 2C9 and microsomal glutathione-S-transferase. Environ. Mol. Mutagen. 2000, 35, 123–131. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, H.; de Saeger, S.; de Boevre, M.; Sun, F.; Zhang, S.; Cao, X.; Wang, Z. In vitro and in vivo metabolism of Ochratoxin A: A comparative study using ultra-performance liquid chromatography-quadrupole/time-of-flight hybrid mass spectrometry. Anal. Bioanal. Chem. 2015, 407, 3579–3589. [Google Scholar] [CrossRef] [PubMed]
- Ueno, Y. Biotransformation of mycotoxins in the reconstituted cytochrome P-450 system. Proc. Jpn. Assoc. Mycotoxicol. 1985, 22, 28–30. [Google Scholar] [CrossRef]
- Stormer, F.C.; Storen, O.; Hansen, C.E.; Pedersen, J.I.; Aasen, A.J. Formation of (4R)- and (4S)-4-hydroxyochratoxin A and 10-hydroxyochratoxin A from Ochratoxin A by rabbit liver microsomes. Appl. Environ. Microbiol. 1983, 45, 1183–1187. [Google Scholar] [PubMed]
- Fink-Gremmels, J.; Jahn, A.; Blom, M.J. Toxicity and metabolism of Ochratoxin A. Nat. Toxins 1995, 3, 214–220. [Google Scholar] [CrossRef]
- Simarro Doorten, A.Y.; Bull, S.; van der Doelen, M.A.; Fink-Gremmels, J. Metabolism-mediated cytotoxicity of Ochratoxin A. Toxicol. In Vitro 2004, 18, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Omar, R.F.; Gelboin, H.V.; Rahimtula, A.D. Effect of cytochrome P450 induction on the metabolism and toxicity of Ochratoxin A. Biochem. Pharmacol. 1996, 51, 207–216. [Google Scholar] [CrossRef]
- Grosse, Y.; Monje, M.C.; Mace, K.; Pfeifer, A.; Pfohl-Leszkowicz, A. Use of bronchial epithelial cells expressing human cytochrome P450 for study on metabolism and genotoxicity of Ochratoxin A. In Vitro Toxicol. 1997, 10, 93–102. [Google Scholar]
- Chu, F.S.; Noh, I.; Chang, C.C. Structure requirements for Ochratoxin A intoxication. Life Sci. 1972, 11, 503–508. [Google Scholar]
- Grosse, Y.; Baudrimont, I.; Castegnaro, M.; Creppy, E.E.; Dirheimer, G.; Pfohl-Leskowicz, A. Ochratoxin A metabolites and DNA-adducts formation in monkey kidney cell. Chem. Biol. Interact. 1995, 95, 175–187. [Google Scholar] [CrossRef]
- Kasmuller, S.; Cavin, C.; Chakraborty, A.; Darroudi, F.; Majer, B.J.; Huber, W.W.; Ehrlich, V.A. Structurally related mycotoxins Ochratoxin A, Ochratoxin B, and citrinin differ in their genotoxic activities and in their mode of action in human-derived liver (HepG2) cells: Implication for Risk Assessment. Nutr. Cancer 2004, 50, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Faucet-Marquis, V.; Pont, F.; Størmer, F.C.; Rizk, T.; Castegnaro, M.; Pfohl-Leszkowicz, A. Evidence of a new dechlorinated Ochratoxin A derivative formed in opossum kidney cell cultures after pretreatment by modulators of glutathione pathways: Correlation with DNA-adduct formation. Mol. Nutr. Food Res. 2006, 50, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Gross-Steinmeyer, K.; Weymann, J.; Hege, H.G.; Metzler, M. Metabolism and lack of DNA reactivity of the mycotoxin Ochratoxin A in cultured and human primary hepatocytes. J. Agric. Food Chem. 2002, 50, 935–938. [Google Scholar] [CrossRef]
- Zepnik, H.; Volkel, W.; Dekant, W. Toxicokinetics of the mycotoxin Ochratoxin A in F 344 rats after oral administration. Toxicol. Appl. Pharmacol. 2003, 192, 36–44. [Google Scholar] [CrossRef]
- Malaveille, C.; Brun, G.; Bartsch, H. Structure-activity studies in E. coli strains on Ochratoxin A (OTA) and its analogues implicate a genotoxic free radical and a cytosolic thiol derivative as reactive metabolites. Mutat. Res. 1994, 307, 141–147. [Google Scholar] [CrossRef]
- Dai, J.; Park, G.; Wright, M.W.; Adams, M.; Akman, S.A.; Manderville, R.A. Detection and characterization of a glutathione conjugate of Ochratoxin A. Chem. Res. Toxicol. 2002, 15, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Tozlovanu, M.; Canadas, D.; Pfohl-Leszkowicz, A.; Frenette, C.; Paugh, R.J.; Manderville, R.A. Glutathione conjugates of Ochratoxin A as biomarkers of exposure. Arh. Hig. Rada Toksikol. 2012, 63, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Pfohl-Leszkowicz, A.; Pinelli, E.; Bartsch, H.; Mohr, U.; Castegnaro, M. Sex and strain differences in Ochratoxin A metabolism and DNA adduction in two strains of rats. Mol. Carcinog. 1998, 23, 76–83. [Google Scholar] [CrossRef]
- Petkova-Bocharova, T.; El Adlouni, C.; Faucet, V.; Pfohl-Leszkowicz, A.; Mantle, P. Analysis for DNA adducts, Ochratoxin A content and enzymes expression in kidneys of pigs exposed to mild experimental chronic ochratoxicosis. Facta Univ. Ser. Med. Biol. 2003, 10, 111–115. [Google Scholar]
- Pfohl-Leszkowicz, A.; Tozlovanu, M.; Manderville, R.; Peraica, M.; Castegnaro, M.; Stefanovic, V. New molecular and field evidences for the implication of mycotoxins but not aristolochic acid in human nephropathy and urinary tract tumor. Mol. Nutr. Food Res. 2007, 51, 1131–1146. [Google Scholar] [CrossRef] [PubMed]
- Gekle, M.; Silbernagl, S. The role of the proximal tubule in Ochratoxin A nephrotoxicity in vivo. Toxicodynamics and toxicokinetics aspects. Ren. Physiol. Biochem. 1994, 17, 40–49. [Google Scholar] [PubMed]
- Groves, C.E.; Morales, M.; Wright, S.H. Peritubular transport of Ochratoxin A in rabbit renal proximal tubules. J. Pharmacol. Exp. Ther. 1998, 284, 943–948. [Google Scholar] [PubMed]
- Kusushura, H.; Sekine, T.; Utsunomiya-Tate, N.M.; Tsuda, M.; Kojima, R.; Cha, S.H.; Sugiyama, Y.; Kanai, Y.; Endou, H. Molecular cloning and characterization of a new multispecific organic anion transporter from rat brain. J. Biol. Chem. 1999, 274, 13675–13680. [Google Scholar] [CrossRef]
- Bahnemann, E.; Kerling, H.P.; Ensminger, S.; Scwerdt, G.; Silbernagl, S.; Gekle, M. Renal transepithelial secretion of Ochratoxin A in the non-filtering toad kidney. Toxicology 1997, 120, 11–17. [Google Scholar] [CrossRef]
- Leier, I.; Hummel-Eisenbeiss, J.; Cui, Y.; Keppler, D. ATP-dependent para-aminohippurate transport by apical multidrug resistance protein MRP2. Kidney Int. 2000, 57, 1636–1642. [Google Scholar] [CrossRef] [PubMed]
- Dahlman, A.; Dantzker, W.H.; Silbernagl, S.; Gekle, M. Detailed mapping of Ochratoxin A reabsorption along the rate nephron in vivo: The nephrotoxin can be reabsorbed in all nephron segments by different mechanisms. J. Pharmacol. Exp. Ther. 1998, 286, 157–162. [Google Scholar]
- Sreemannarayana, O.; Frohlich, A.A.; Vitti, T.G.; Marquardt, R.R.; Abramson, D.; Phillips, G.D. Studies of the tolerance and disposition of Ochratoxin A in young calves. J. Anim. Sci. 1988, 66, 1703–1711. [Google Scholar] [PubMed]
- Suzuki, S.; Satoh, T.; Yamakazi, M. The pharmacokinetics of Ochratoxin A in rats. Jpn. J. Pharmacol. 1977, 27, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Nip, W.K.; Chu, F.S. The fate of Ochratoxin A in goats. J. Environ. Sci. Health B 1979, 14, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Marquardt, R.R.; Frohlich, A.A.; Philips, G.D.; Vitti, T.G. Effect of hay and a grain diet on the rate of hydrolysis of Ochratoxin A in the rumen sheep. J. Anim. Sci. 1991, 69, 3706–3714. [Google Scholar] [PubMed]
- Fuchs, R.; Radic, B.; Peraica, M.; Hult, K.; Plestina, R. Enterohepatic circulation of Ochratoxin A in rats. Period. Biol. 1988, 90, 39–42. [Google Scholar]
- Gareis, M.; Martlbauer, E.; Bauer, J.; Gedek, B. Determination of Ochratoxin A in breast milk. Zeitschr. Lebensm. Undersch. Forsch. 1988, 186, 114–117. [Google Scholar] [CrossRef]
- Micco, C.; Miraglia, M.; Brera, C.; Corneli, S.; Ambruzi, A. Evaluation of Ochratoxin A level in human milk in Italy. Food Addit. Contam. 1995, 12, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Skaug, M.A.; Helland, I.; Solvoll, K.; Saugstad, O.D. Presence of Ochratoxin A in human milk in relation to dietary intake. Food Addit. Contam. 2001, 18, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, K.; Blaszkewicz, M.; Campos, V.; Vega, M.; Degen, G.H. Exposure of infants to Ochratoxin A with breast milk. Arch. Toxicol. 2014, 88, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Creppy, E.E.; Röschenthaler, R.; Dirheimer, G. Inhibition of protein synthesis in mice by Ochratoxin A and its prevention by phenylalanine. Food Chem. Toxicol. 1984, 22, 883–886. [Google Scholar] [CrossRef]
- Dirheimer, G.; Creppy, E.E. Mechanism of action of Ochratoxin A. IARC Sci. Publ. 1991, 115, 171–186. [Google Scholar] [PubMed]
- Bruinink, A.; Rasony, T.; Sidler, C. Reduction of Ochratoxin A toxicity by heat-induced epimerization. In vitro effects of ochratoxins on embryonic chick meningeal and other cell cultures. Toxicology 1997, 118, 205–210. [Google Scholar] [CrossRef]
- Bruinink, A.; Sidler, C. The neurotoxic effect of Ochratoxin A are reduced by protein binding but not affected byl-phenylalanine. Toxicol. Appl. Pharmacol. 1997, 146, 173–179. [Google Scholar] [CrossRef] [PubMed]
- McMaster, D.R.; Vedani, A. Ochratoxin A binding to phenlylalanine-tRNA synthetatse: Computational approach to the mechanism of ochratoxicoses and its antagonism. J. Med. Chem. 1999, 42, 3075–3086. [Google Scholar] [CrossRef] [PubMed]
- Creppy, E.E.; Chakor, K.; Fisher, M.; Dirheimer, G. The mycotoxin Ochratoxin A is a substrate for phenylalanine hydroxylase in isolated rat hepatocytes and in vivo. Arch. Toxicol. 1990, 64, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Vettorazzi, A.; van Delft, J.; de Cerain, A.L. A review on Ochratoxin A transcriptomic studies. Food Chem. Toxicol. 2013, 59, 766–783. [Google Scholar] [CrossRef] [PubMed]
- Poór, M.; Veres, B.; Jakus, P.B.; Antus, C.; Montskó, G.; Zrínyi, Z.; Vladimir-Knežević, S.; Petrik, J.; Kőszegi, T. Flavonoid diosmetin increases ATP levels in kidney cells and relieves ATP depleting effect of Ochratoxin A. J. Photochem. Photobiol. B 2014, 132, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Aleo, M.D.; Wyat, R.D.; Schnellmann, R.G. Mitochondrial dysfunction is an early event in Ochratoxin A but not oosporein toxicity to rat renal proximal tubules. Toxicol. Appl. Pharmacol. 1991, 107, 73–80. [Google Scholar] [CrossRef]
- Meisner, H.; Meisner, P. Ochratoxin A an in vivo inhibitor of renal phosphoenolpyruvate carboxykinase. Arch. Biochem. Biophys. 1981, 208, 146–153. [Google Scholar] [CrossRef]
- Meisner, H.; Krogh, P. Phosphoenolpyruvate carboxykinase as a selective indicator of Ochratoxin A induced nephropathy. Dev. Toxicol. Environ. Sci. 1986, 14, 199–206. [Google Scholar] [PubMed]
- Meisner, H.; Polsinelli, P. Changes of renal mRNA species abundance by Ochratoxin A. Biochem. Pharmacol. 1986, 35, 661–665. [Google Scholar] [CrossRef]
- Thekkumkarra, T.J.; Patel, M.S. Ochratoxin A decrease the activity of phosphoenolpyruvate carboxykinase and its mRNA content in primary cultures of rat kidney proximal convoluted tubular cells. Biochem. Biophys. Res. Commun. 1989, 162, 916–920. [Google Scholar] [CrossRef]
- Suzuki, S.; Kozuka, Y.; Satoh, T.; Yamazaki, M. Studies on the nephrotoxicity of Ochratoxin A in rats. Toxicol. Appl. Pharmacol. 1975, 34, 479–490. [Google Scholar] [CrossRef]
- Meisner, H.; Chan, S. Ochratoxin A, an inhibitor of mitochondrial transport systems. Biochemistry 1974, 13, 2795–2800. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.H.; Lu, T.N.; Wei, R.D. Effect of Ochratoxin A on rat liver mitochondrial respiration and oxidative phosphorilation. Toxicology 1985, 36, 119–123. [Google Scholar] [CrossRef]
- Manderville, R.A.; Pfohl-Leszkowicz, A. Genotoxicity of chlorophenols and Ochratoxin A. Adv. Mol. Toxicol. 2006, 1, 73–118. [Google Scholar]
- Waidyanatha, S.; Lin, P.H.; Rappaport, S.M. Characterization of chlorinated adducts of hemoglobin and albumin following administration of pentachlorophenol to rats. Chem. Res. Toxicol. 1996, 9, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, V.G.; Villalta, P.W.; Sturla, S.J. Nucleobase-dependent reactivity of a quinone metabolite of pentachlorophenol. Chem. Res. Toxicol. 2007, 20, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Tozlovanu, M.; Faucet-Marquis, V.; Pfohl-Leszkowicz, A.; Manderville, R.A. Genotoxicity of the hydroquinone metabolite of Ochratoxin A: Structure-activity relationships for covalent DNA adduction. Chem. Res. Toxicol. 2006, 19, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Gillman, I.G.; Clark, T.N.; Manderville, R.A. Oxidation of Ochratoxin A by an Fe-porphyrin system: Model for enzymatic activation and DNA cleavage. Chem. Res. Toxicol. 1999, 12, 1066–1076. [Google Scholar] [CrossRef] [PubMed]
- Reljic, Z.; Zlatovic, M.; Savic-Radojevic, A.; Pekmezovic, T.; Djukanovic, L.; Matic, M.; Pljesa-Ercegovac, M.; Mimic-Oka, J.; Opsenica, D.; Simic, T. Is increased susceptibility to Balkan endemic nephropathy in carriers of common GSTA1 (*A/*B) polymorphism linked with the catalytic role of GSTA1 in Ochratoxin A biotransformation? Serbian case control study and in silico analysis. Toxins 2014, 6, 2348–2362. [Google Scholar] [CrossRef] [PubMed]
- Mally, A.; Zepnik, H.; Wanek, P.; Eder, E.; Dingley, K.; Ihmels, H.; Völkel, W.; Dekant, W. Ochratoxin A: Lack of formation of covalent DNA adducts. Chem. Res. Toxicol. 2004, 17, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Samokyszyn, V.M.; Freeman, J.P.; Maddipati, K.R.; Lloyd, R.V. Peroxidase-catalyzed oxidation of pentachlorophenol. Chem. Res. Toxicol. 1995, 8, 349–355. [Google Scholar] [CrossRef] [PubMed]
- El Adlouni, C.; Pinelli, E.; Azémar, B.; Zaoui, D.; Beaune, P.; Pfohl-Leszkowicz, A. Role of CYP 2C and microsomal glutathione-S-transferase in modulating susceptibility to Ochratoxin A genotoxicity. Environ. Mol. Mutagen. 2000, 35, 123–131. [Google Scholar] [CrossRef]
- Stoyanovosky, D.A.; Goldman, R.; Jonnalagadda, S.S.; Day, B.W.; Claycamp, H.G.; Kagan, V.E. Detection and characterization of the electron paramagnetic resonance-silent glutathionyl-5,5-dimethyl-1-pyrroline N-oxide adduct derived from redox cycling of phenoxyl radicals in model systems and HL-60 cells. Arch. Biochem. Biophys. 1996, 330, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Murray, A.R.; Kisin, E.; Castranova, V.; Kommineni, C.; Gunther, M.R.; Shvedova, A.A. Phenol-induced in vivo oxidative stress in skin: Evidence for enhanced free radical generation, thiol oxidation, and antioxidant depletion. Chem. Res. Toxicol. 2007, 20, 1769–1777. [Google Scholar] [CrossRef] [PubMed]
- Kamp, H.G.; Eisenbrand, G.; Schlatter, J.; Würth, K.; Janzowski, C. Ochratoxin A: Induction of (oxidative) DNA damage, cytotoxicity and apoptosis in mammalian cell lines and primary cells. Toxicology 2005, 206, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Wright, M.W.; Manderville, R.A. An oxygen-bonded C8-deoxyguanosine nucleoside adduct of pentachlorophenol by peroxidase activation: Evidence for ambident C8 reactivity by phenoxyl radicals. Chem. Res. Toxicol. 2003, 16, 817–821. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Sloat, A.L.; Wright, M.W.; Manderville, R.A. Role of phenoxyl radicals in DNA adduction by chlorophenol xenobiotics following peroxidase activation. Chem. Res. Toxicol. 2005, 18, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Mohn, W.W.; Kennedy, K.J. Reductive dehalogenation of chlorophenols by Desulfomonile tiedjei DCB-1. Appl. Environ. Microbiol. 1992, 58, 1367–1370. [Google Scholar] [PubMed]
- Lawson, T.; Gannett, P.M.; Yau, W.M.; Dalal, N.S.; Toth, B. Different patterns of mutagenicity of arenediazonium ions in V79 cells and Salmonella typhimurium TA102: Evidence for different mechanisms of action. J. Agric. Food Chem. 1995, 43, 2627–2635. [Google Scholar] [CrossRef]
- Hiramoto, K.; Kaku, M.; Sueyoshi, A.; Fujise, M.; Kikugawa, K. DNA base and deoxyribose modification by the carbon-centered radical generated from 4-(hydroxymethyl)-benzenediazonium salt, a carcinogen in mushroom. Chem. Res. Toxicol. 1995, 8, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Gannett, P.M.; Powell, J.H.; Rao, R.; Shi, X.; Lawson, T.; Kolar, C.; Toth, B. C8-Arylguanine and C8-aryladenine formation in calf thymus DNA from arenediazonium ions. Chem. Res. Toxicol. 1999, 12, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Wright, M.W.; Manderville, R.A. Ochratoxin a forms a carbon-bonded C8-deoxyguanosine nucleoside adduct: Implications for C8 reactivity by a phenolic radical. J. Am. Chem. Soc. 2003, 125, 3716–3717. [Google Scholar] [CrossRef] [PubMed]
- Il'ichev, Y.V.; Perry, J.L.; Manderville, R.A.; Chignell, C.F.; Simon, J.D. The pH-dependent primary photoreactions of Ochratoxin A. J. Phys. Chem. B 2001, 105, 11369–11376. [Google Scholar] [CrossRef]
- Faucet, V.; Pfohl-Leszkowicz, A.; Dai, J.; Castegnaro, M.; Manderville, R.A. Evidence for covalent DNA adduction by Ochratoxin A following chronic exposure to rat and subacute exposure to pig. Chem. Res. Toxicol. 2004, 17, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Pfohl-Leszkowicz, A.; Gabryelski, W.; Manderville, R.A. Formation of 2′-deoxyguanosine-carbon 8-bound Ochratoxin A adduct in rat kidney DNA. Mol. Nutr. Food Res. 2009, 53, 154–155. [Google Scholar] [CrossRef] [PubMed]
- Pfohl-Leszkowicz, A.; Castegnaro, M. Further arguments in favour of direct covalent binding of Ochratoxin A (OTA) after metabolic biotransformation. Food Addit. Contam. 2005, 22 (Suppl. 1), 75–87. [Google Scholar] [CrossRef] [PubMed]
- Mantle, P.G.; Faucet-Marquis, V.; Manderville, R.A.; Squillaci, B.; Pfohl-Leszkowicz, A. Structures of covalent adducts between DNA and Ochratoxin A: A new factor in debate about genotoxicity and human risk assessment. Chem. Res. Toxicol. 2010, 23, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Mally, A.; Pepe, G.; Ravoori, S.; Fiore, M.; Gupta, R.C.; Dekant, W.; Mosesso, P. Ochratoxin A causes DNA damage and cytogenetic effects but no DNA adducts in rats. Chem. Res. Toxicol. 2005, 18, 1253–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delatour, T.; Mally, A.; Richoz, J.; Ozden, S.; Dekant, W.; Ihmels, H.; Otto, D.; Gasparutto, D.; Marin-Kuan, M.; Schilter, B.; et al. Absence of 2′-deoxyguanosine-carbon 8-bound Ochratoxin A adduct in rat kidney DNA monitored by isotope dilution LC-MS/MS. Mol. Nutr. Food Res. 2008, 52, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Mally, A. Ochratoxin A and mitotic disruption: Mode of action analysis of renal tumor formation by Ochratoxin A. Toxicol. Sci. 2012, 127, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Stiborová, M.; Bárta, F.; Levová, K.; Hodek, P.; Frei, E.; Arlt, V.M.; Schmeiser, H.H. The influence of Ochratoxin A on DNA adduct formation by the carcinogen aristolochic acid in rats. Arch. Toxicol. 2015, 89, 2141–2158. [Google Scholar] [CrossRef] [PubMed]
- Sorrenti, V.; di Giacomo, C.; Acquaviva, R.; Barbagallo, I.; Bognanno, M.; Galvano, F. Toxicity of Ochratoxin A and its modulation by antioxidants: A review. Toxins 2013, 5, 1742–1766. [Google Scholar] [CrossRef] [PubMed]
- Rahimtula, A.D.; Béréziat, J.C.; Bussacchini-Griot, V.; Bartsch, H. Lipid peroxidation as a possible cause of Ochratoxin A toxicity. Biochem. Pharmacol. 1988, 37, 4469–4477. [Google Scholar] [CrossRef]
- Omar, R.F.; Hasinoff, B.B.; Mejilla, F.; Rahimtula, A.D. Mechanism of Ochratoxin A stimulated lipid peroxidation. Biochem. Pharmacol. 1990, 40, 1183–1191. [Google Scholar] [CrossRef]
- Ranaldi, G.; Caprini, V.; Sambuy, Y.; Perozzi, G.; Murgia, C. Intracellular zinc stores protect the intestinal epithelium from Ochratoxin A toxicity. Toxicol. In Vitro 2009, 23, 1516–1521. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhang, Y.; Xu, W.; Luo, Y.; Hao, J.; Shen, X.L.; Yang, X.; Li, X.; Huang, K. Zinc protects HepG2 cells against the oxidative damage and DNA damage induced by Ochratoxin A. Toxicol. Appl. Pharmacol. 2013, 268, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Poór, M.; Kuzma, M.; Matisz, G.; Li, Y.; Perjési, P.; Kunsági-Máté, S.; Kőszegi, T. Further aspects of Ochratoxin A-cation interactions: Complex formation with zinc ions and a novel analytical application of Ochratoxin A-magnesium interaction in the HPLC-FLD system. Toxins 2014, 6, 1295–1307. [Google Scholar] [CrossRef] [PubMed]
- Marin-Kuan, M.; Nestler, S.; Verguet, C.; Bezençon, C.; Piguet, D.; Mansourian, R.; Holzwarth, J.; Grigorov, M.; Delatour, T.; Mantle, P.; et al. A toxicogenomics approach to identify new plausible epigenetic mechanisms of Ochratoxin A carcinogenicity in rat. Toxicol. Sci. 2006, 89, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Cavin, C.; Delatour, T.; Marin-Kuan, M.; Holzhäuser, D.; Higgins, L.; Bezençon, C.; Guignard, G.; Junod, S.; Richoz-Payot, J.; Gremaud, E.; et al. Reduction in antioxidant defenses may contribute to Ochratoxin A toxicity and carcinogenicity. Toxicol. Sci. 2007, 96, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Boesch-Saadatmandi, C.; Loboda, A.; Jozkowicz, A.; Huebbe, P.; Blank, R.; Wolffram, S.; Dulak, J.; Rimbach, G. Effect of Ochratoxin A on redox-regulated transcription factors, antioxidant enzymes and glutathione-S-transferase in cultured kidney tubulus cells. Food Chem. Toxicol. 2008, 46, 2665–2671. [Google Scholar] [CrossRef] [PubMed]
- Limonciel, A.; Jennings, P. A review of the evidence that Ochratoxin A is an Nrf2 inhibitor: Implications for nephrotoxicity and renal carcinogenicity. Toxins 2014, 6, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Cavin, C.; Delatour, T.; Marin-Kuan, M.; Fenaille, F.; Holzhäuser, D.; Guignard, G.; Bezençon, C.; Piguet, D.; Parisod, V.; Richoz-Payot, J.; et al. Ochratoxin A-mediated DNA and protein damage: Roles of nitrosative and oxidative stresses. Toxicol. Sci. 2009, 110, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Yu, T.; Zhu, L.; Gao, J.; He, X.; Huang, K.; Luo, Y.; Xu, W. Ochratoxin A induces rat renal carcinogenicity with limited induction of oxidative stress responses. Toxicol. Appl. Pharmacol. 2014, 280, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Taniai, E.; Yafune, A.; Nakajima, M.; Hayashi, S.M.; Nakane, F.; Itahashi, M.; Shibutani, M. Ochratoxin A induces karyomegaly and cell cycle aberrations in renal tubular cells without relation to induction of oxidative stress responses in rats. Toxicol. Lett. 2014, 224, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Gekle, M.; Schwerdt, G.; Freudinger, R.; Mildenberg, S.; Wilflingseder, D.; Pollack, V.; Dander, M.; Schramek, H. Ochratoxin A induces JNK activation and apoptosis in MDKC-C7 cells at nanomolar concentrations. J. Pharmacol. Exp. Ther. 2000, 293, 837–844. [Google Scholar] [PubMed]
- Lühe, A.; Hildebrandt, H.; Bach, U.; Dingerman, T.; Ahr, H.J. A new approach to studying Ochratoxin A (OTA)-induced nephrotoxicity: Expressing profiling in vivo and in vitro employing cDNA microarrays. Toxicol. Sci. 2003, 73, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K.; Hibi, D.; Ishii, Y.; Yokoo, Y.; Takasu, S.; Kijima, A.; Matsushita, K.; Masumura, K.; Kodama, Y.; Yanai, T.; et al. Role of p53 in the progression from ochratoxin A-induced DNA damage to gene mutations in the kidneys of mice. Toxicol. Sci. 2015, 144, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Horvath, A.; Upham, B.L.; Ganev, V.; Trosko, J.E. Determination of the epigenetic effects of ochratoxin in a human kidney and a rat liver epithelial cell line. Toxicon 2002, 40, 273–282. [Google Scholar] [CrossRef]
- Sauvant, C.; Holzinger, H.; Gekle, M. The nephrotoxin Ochratoxin A induces key parameters of chronic interstitial nephropathy in renal proximal tubular cells. Cell Physiol. Biochem. 2005, 15, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Özcan, Z.; Gül, G.; Yaman, I. Ochratoxin A activates opposing c-MET/PI3K/Akt and MAPK/ERK 1–2 pathways in human proximal tubule HK-2 cells. Arch. Toxicol. 2015, 89, 1313–1327. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Shen, X.L.; Zhang, B.; Li, Y.; Xu, W.; Zhao, C.; Luo, Y.; Huang, K. Apoptosis signal-regulating kinase 1 promotes Ochratoxin A-induced renal cytotoxicity. Sci. Rep. 2015, 5, 8078. [Google Scholar] [CrossRef] [PubMed]
- Atroshi, F.; Biese, I.; Saloniemi, H. Significance of apoptosis and its relationship to antioxidants after Ochratoxin A administration in mice. J. Pharm. Pharm. Sci. 2000, 3, 281–291. [Google Scholar] [PubMed]
- Petrik, J.; Zanic-Grubisic, T.; Barisic, K.; Pepeljnjak, S.; Radic, B.; Ferencic, Z.; Cepelak, I. Apoptosis and oxidative stress induced by Ochratoxin A in rat kidney. Arch. Toxicol. 2003, 77, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Weidenbach, A.; Schuh, K.; Failing, K.; Petzinger, E. Ochratoxin A induced TNF-release from the isolated and blood-free perfused rat liver. Mycotoxin Res. 2000, 16, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Al-Anati, L.; Katz, N.; Petzinger, E. Interference of arachidonic acid and its metabolites with TNFα release by Ochratoxin A from rat liver. Toxicology 2005, 208, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Petzinger, E.; Weidenbach, A. Mycotoxins in the food chain: The role of ochratoxins. Livest. Prod. Sci. 2002, 76, 245–250. [Google Scholar] [CrossRef]
- Rached, E.; Pfeiffer, E.; Dekant, W.; Mally, A. Ochratoxin A: Apoptosis and aberrant exit from mitosis due to perturbation of microtubule dynamics? Toxicol. Sci. 2006, 92, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Mally, A.; Dekant, W. Mycotoxins and the kidney: Modes of action for renal tumor formation by Ochratoxin A in rodents. Mol. Nutr. Food Res. 2009, 53, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Adler, M.; Müller, K.; Rached, E.; Dekant, W.; Mally, A. Modulation of key regulators of mitosis linked to chromosomal instability is an early event in Ochratoxin A carcinogenicity. Carcinogenesis 2009, 30, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Czakai, K.; Müller, K.; Mosesso, P.; Pepe, G.; Schulze, M.; Gohla, A.; Patnaik, D.; Dekant, W.; Higgins, J.M.G.; Mally, A. Perturbation of mitosis through inhibition of histone acetyltransferases: The key to Ochratoxin A toxicity and carcinogenicity? Toxicol. Sci. 2011, 122, 317–329. [Google Scholar] [CrossRef] [PubMed]
- O'Brien, E.; Heussner, A.H.; Dietrich, D.R. Species-, sex-, and cell type-specific effects of Ochratoxin A and B. Toxicol. Sci. 2001, 63, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Palma, N.; Cinelli, S.; Sapora, O.; Wilson, S.H.; Dogliotti, E. Ochratoxin A-induced mutagenesis in mammalian cells is consistent with the production of oxidative stress. Chem. Res. Toxicol. 2007, 20, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, J.; Cui, J.; Xing, L.; Wang, J.; Yan, X.; Zhang, X. ERK and p38 MAPK signaling pathways are involved in Ochratoxin A-induced G2 phase arrest in human gastric epithelium cells. Toxicol. Lett. 2012, 209, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Liu, J.; Wu, S.; Wang, Y.; Shen, H.; Xing, L.; Wang, J.; Yan, X.; Zhang, X. Oxidative DNA damage is involved in Ochratoxin A-induced G2 arrest through ataxia telangiectasia-mutated (ATM) pathways in human gastric epithelium GES-1 cells in vitro. Arch. Toxicol. 2013, 87, 1829–1840. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Y.; Cui, J.; Xing, L.; Shen, H.; Wu, S.; Lian, H.; Wang, J.; Yan, X.; Zhang, X. Ochratoxin A induces oxidative DNA damage and G1 phase arrest in human peripheral blood mononuclear cells in vitro. Toxicol. Lett. 2012, 211, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; He, X.; Li, X.; Xu, W.; Luo, Y.; Yang, X.; Wang, Y.; Li, Y.; Huang, K. DNA damage and S phase arrest induced by Ochratoxin A in human embryonic kidney cells (HEK 293). Mutat. Res. 2014, 765, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Shi, L.; Huang, K.; Xu, W. Protective effect of N-acetylcysteine against DNA damage and S-phase arrest induced by Ochratoxin A in human embryonic kidney cells (HEK-293). Food Chem Toxicol. 2014, 70, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Martin, M.; Bartsch, H.; Rahimtula, A.D. Perturbation of liver microsomal calcium homeostasis by Ochratoxin A. Biochem. Pharmacol. 1989, 38, 67–72. [Google Scholar] [CrossRef]
- Dopp, E.; Müller, J.; Hahnel, C.; Schiffmann, D. Induction of genotoxic effects and modulation of the intracellular calcium level in Syrian Hamster Embryo (SHE) fibroblasts caused by Ochratoxin A. Food Chem. Toxicol. 1999, 37, 713–721. [Google Scholar] [CrossRef]
- Hoehler, D.; Marquardt, R.R.; McIntosh, A.R.; Xiao, H. Free radical generation as induced by Ochratoxin A and its analogs in bacteria (Bacillus brevis). J. Biol. Chem. 1996, 271, 27388–27394. [Google Scholar] [CrossRef] [PubMed]
- Hoehler, D.; Marquardt, R.R.; McIntosh, A.R.; Hatch, G.M. Induction of free radicals in hepatocytes, mitochondria and microsomes of rats by Ochratoxin A and its analogs. Biochim. Biophys. Acta 1997, 1357, 225–233. [Google Scholar] [CrossRef]
- Benesic, A.; Mildenberger, S.; Gekle, M. Nephritogenic Ochratoxin A interferes with hormonal signalling in immortalized human kidney epithelial cells. Pflügers Arch. 2000, 439, 278–287. [Google Scholar] [CrossRef] [PubMed]
- García, A.R.; Avila, E.; Rosiles, R.; Petrone, V.M. Evaluation of two mycotoxin binders to reduce toxicity of broiler diets containing Ochratoxin A and T-2 toxin contaminated grain. Avian Dis. 2003, 47, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Trailović, J.N.; Stefanović, S.; Trailović, S.M. In vitro and in vivo protective effects of three mycotoxin adsorbents against Ochratoxin A in broiler chickens. Br. Poult. Sci. 2013, 54, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Madhyastha, M.S.; Frohlich, A.A.; Marquardt, R.R. Effect of dietary cholestyramine on the elimination pattern of Ochratoxin A in rats. Food Chem. Toxicol. 1992, 30, 709–714. [Google Scholar] [CrossRef]
- Kerkadi, A.; Barriault, C.; Marquardt, R.R.; Frohlich, A.A.; Yousef, I.M.; Zhu, X.X.; Tuchweber, B. Cholestyramine protection against Ochratoxin A toxicity: Role of Ochratoxin A sorption by the resin and bile acid enterohepatic circulation. J. Food Prot. 1999, 62, 1461–1465. [Google Scholar] [PubMed]
- Yong, S.; Albassam, M.; Prior, M. Protective effects of sodium bicarbonate on murine ochratoxicosis. J. Environ. Sci. Health B 1987, 22, 455–470. [Google Scholar] [CrossRef] [PubMed]
- Baudrimont, I.; Murn, M.; Betbeder, A.M.; Guilcher, J.; Creppy, E.E. Effect of piroxicam on the nephrotoxicity induced by Ochratoxin A in rats. Toxicology 1995, 95, 147–154. [Google Scholar] [CrossRef]
- Moroi, K.; Suzuki, S.; Kuga, T.; Yamazaki, M.; Kanisawa, M. Reduction of Ochratoxin A toxicity in mice treated with phenylalanine and phenobarbital. Toxicol. Lett. 1985, 25, 1–5. [Google Scholar] [PubMed]
- Chakor, K.; Creppy, E.E.; Dirheimer, G. In vivo Studies on the relationship between hepatic metabolism and toxicity of Ochratoxin A. Arch. Toxicol. 1988, 12, 201–204. [Google Scholar]
- Suzuki, S.; Moroi, K.; Kanisawa, M.; Satoh, T. Effect of drug metabolizing enzyme inducers on carcinogenesis and toxicity of Ochratoxin A in mice. Toxicol. Lett. 1986, 31, 206. [Google Scholar]
- Creppy, E.E.; Schlegel, M.; Röschenthaler, R.; Dirheimer, G. Phenylalanine prevents acute poisoning by ochratoxina in mice. Toxicol. Lett. 1980, 6, 77–80. [Google Scholar] [CrossRef]
- Baudrimont, I.; Betbeder, A.M.; Creppy, E.E. Reduction of the Ochratoxin A-induced cytotoxicity in Vero cells by aspartame. Arch. Toxicol. 1997, 71, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Baudrimont, I.; Sostaric, B.; Yenot, C.; Betbeder, A.M.; Dano-Djedje, S.; Sanni, A.; Steyn, P.S.; Creppy, E.E. Aspartame prevents the karyomegaly induced by Ochratoxin A in rat kidney. Arch. Toxicol. 2001, 75, 176–183. [Google Scholar] [PubMed]
- Stoev, S.D. Studies on carcinogenic and toxic effects of Ochratoxin A in chicks. Toxins 2010, 2, 649–664. [Google Scholar] [CrossRef] [PubMed]
- Creppy, E.E.; Baudrimont, I.; Belmadani, A.; Betbeder, A.M. Aspartame as a preventive agent of chronic toxic effects of Ochratoxin A in experimental animals. Toxin Rev. 1996, 15, 207–221. [Google Scholar] [CrossRef]
- Pfohl-Leszkowicz, A.; Bartsch, H.; Azémar, B.; Mohr, U.; Estève, J.; Castegnaro, M. MESNA protects rats against nephrotoxicity but not carcinogenicity induced by Ochratoxin A, implicating two separate pathways. Facta Univ. Ser. Med. Biol. 2002, 9, 57–63. [Google Scholar]
- Kumari, D.; Sinha, S.P. Effect of retinol on ochratoxin-produced genotoxicity in mice. Food Chem. Toxicol. 1994, 32, 471–475. [Google Scholar] [CrossRef]
- Hoehler, D.; Marquardt, R.R. Influence of vitamins E and C on the toxic effects of Ochratoxin A and T-2 toxin in chicks. Poult. Sci. 1996, 75, 1508–1515. [Google Scholar] [CrossRef] [PubMed]
- Grosse, Y.; Chekir-Ghedira, L.; Huc, A.; Obrecht-Pflumio, S.; Dirheimer, G.; Bacha, H.; Pfohl-Leszkowicz, A. Retinol, ascorbic acid and alpha-tocopherol prevent DNA adduct formation in mice treated with the mycotoxins Ochratoxin A and zearalenone. Cancer Lett. 1997, 114, 225–229. [Google Scholar] [CrossRef]
- Baldi, A.; Losio, M.N.; Cheli, F.; Rebucci, R.; Sangalli, L.; Fusi, E.; Bertasi, B.; Pavoni, E.; Carli, S.; Politis, I. Evaluation of the protective effects of alpha-tocopherol and retinol against Ochratoxin A cytotoxicity. Br. J. Nutr. 2004, 91, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Utan, A.; Cervellati, R.; Speroni, E.; Guerra, M.C. Catechins: Natural free-radical scavengers against Ochratoxin A-induced cell damage in a pig kidney cell line (LLC-PK1). Food Chem. Toxicol. 2007, 45, 1910–1917. [Google Scholar] [CrossRef] [PubMed]
- Renzulli, C.; Galvano, F.; Pierdomenico, L.; Speroni, E.; Guerra, M.C. Effects of rosmarinic acid against aflatoxin B1 and Ochratoxin-A-induced cell damage in a human hepatoma cell line (Hep G2). J. Appl. Toxicol. 2004, 24, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; la Fauci, L.; Acquaviva, R.; Campisi, A.; Raciti, G.; Scifo, C.; Renis, M.; Galvano, G.; Vanella, A.; Galvano, F. Ochratoxin A-induced DNA damage in human fibroblast: Protective effect of cyanidin 3-O-beta-d-glucoside. J. Nutr. Biochem. 2005, 16, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Guerra, M.C.; Galvano, F.; Bonsi, L.; Speroni, E.; Costa, S.; Renzulli, C.; Cervellati, R. Cyanidin-3-O-beta-glucopyranoside, a natural free-radical scavenger against aflatoxin B1- and Ochratoxin A-induced cell damage in a human hepatoma cell line (Hep G2) and a human colonic adenocarcinoma cell line (CaCo-2). Br. J. Nutr. 2005, 94, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Sorrenti, V.; di Giacomo, C.; Acquaviva, R.; Bognanno, M.; Grilli, E.; D'Orazio, N.; Galvano, F. Dimethylarginine dimethylaminohydrolase/nitric oxide synthase pathway in liver and kidney: Protective effect of cyanidin 3-O-β-d-glucoside on Ochratoxin-A toxicity. Toxins 2012, 4, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Essid, E.; Dernawi, Y.; Petzinger, E. Apoptosis induction by OTA and TNF-α in cultured primary rat hepatocytes and prevention by silibinin. Toxins 2012, 4, 1139–1156. [Google Scholar] [CrossRef] [PubMed]
- Cariddi, L.N.; Sabini, M.C.; Escobar, F.M.; Montironi, I.; Manas, F.; Iglesias, D.; Comini, L.R.; Sabini, L.I.; Dalcero, A.M. Polyphenols as possible bioprotectors against cytotoxicity and DNA damage induced by Ochratoxin A. Environ. Toxicol. Pharmacol. 2015, 39, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Ramyaa, P.; Padma, V.V. Ochratoxin-induced toxicity, oxidative stress and apoptosis ameliorated by quercetin—Modulation by Nrf2. Food Chem. Toxicol. 2013, 62, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Ramyaa, P.; Krishnaswamy, R.; Padma, V.V. Quercetin modulates OTA-induced oxidative stress and redox signalling in HepG2 cells—up regulation of Nrf2 expression and down regulation of NF-κB and COX-2. Biochim. Biophys. Acta 2014, 1840, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S.; Palabiyik, S.S.; Erkekoglu, P.; Sahin, G.; Başaran, N.; Giray, B.K. The carotenoid lycopene protects rats against DNA damage induced by Ochratoxin A. Toxicon 2013, 73, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Palabiyik, S.S.; Erkekoglu, P.; Zeybek, N.D.; Kizilgun, M.; Baydar, D.E.; Sahin, G.; Giray, B.K. Protective effect of lycopene against Ochratoxin A induced renal oxidative stress and apoptosis in rats. Exp. Toxicol. Pathol. 2013, 65, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahhab, M.A.; Abdel-Azim, S.H.; El-Nekeety, A.A. Inula crithmoides extract protects against Ochratoxin A-induced oxidative stress, clastogenic and mutagenic alterations in male rats. Toxicon 2008, 52, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, D.; Verma, R. Ameliorative effect of Emblica officinalis aqueous extract on ochratoxin-induced lipid peroxidation in the kidney and liver of mice. Int. J. Occup. Med. Environ. Health 2010, 23, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Gan, F.; Xue, H.; Huang, Y.; Pan, C.; Huang, K. Selenium alleviates porcine nephrotoxicity of Ochratoxin A by improving selenoenzyme expression in vitro. PLoS ONE 2015, 10, e119808. [Google Scholar]
- Mantle, P.; Kilic, M.A.; Mor, F.; Ozmen, O. Contribution of organ vasculature in rat renal analysis for Ochratoxin A: Relevance to toxicology of nephrotoxins. Toxins 2015, 7, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Obrecht-Pflumio, S.; Grosse, Y.; Pfohl-Leszkowicz, A.; Dirheimer, G. Protection by indomethacin and aspirin against genotoxicity of Ochratoxin A, particularly in the urinary bladder and kidney. Arch. Toxicol. 1996, 70, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Baudrimont, I.; Ahouandjivo, R.; Creppy, E.E. Prevention of lipid peroxidation induced by Ochratoxin A in Vero cells in culture by several agents. Chem. Biol. Interact. 1997, 104, 29–40. [Google Scholar] [CrossRef]
- Galtier, P.; Camguilhem, R.; Bodin, G. Evidence for in vitro and in vivo interaction between Ochratoxin A and three acidic drugs. Food Cosmet. Toxicol. 1980, 18, 493–496. [Google Scholar] [CrossRef]
- Creppy, E.E.; Baudrimont, I.; Betbeder, A.M. Prevention of nephrotoxicity of Ochratoxin A, a food contaminant. Toxicol. Lett. 1995, 82–83, 869–877. [Google Scholar] [CrossRef]
- Hong, S.S.; Seo, K.; Lim, S.C.; Han, H.K. Interaction characteristics of flavonoids with human organic anion transporter 1 (hOAT1) and 3 (hOAT3). Pharmacol. Res. 2007, 56, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.C.; Botting, N.P.; Orfila, C.; Al-Maharik, N.; Williamson, G. Flavonoid conjugates interact with organic anion transporters (OATs) and attenuate cytotoxicity of adefovir mediated by organic anion transporter 1 (OAT1/SLC22A6). Biochem. Pharmacol. 2011, 81, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wolkoff, A.W.; Morris, M.E. Flavonoids as a novel class of human organic anion-transporting polypeptide OATP1B1 (OATP-C) modulators. Drug Metab. Dispos. 2005, 33, 1666–1672. [Google Scholar] [CrossRef] [PubMed]
- Sergent, T.; Garsou, S.; Schaut, A.; Saeger, S.D.; Pussemier, L.; Peteghem, C.V.; Larondelle, Y.; Schneider, Y.J. Differential modulation of Ochratoxin A absorption across Caco-2 cells by dietary polyphenols, used at realistic intestinal concentrations. Toxicol. Lett. 2005, 159, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Brand, W.; Schutte, M.E.; Williamson, G.; van Zanden, J.J.; Cnubben, N.H.P.; Groten, J.P.; van Bladeren, P.J.; Rietjens, I.M.C.M. Flavonoid-mediated inhibition of intestinal ABC transporters may affect the oral bioavailability of drugs, food-borne toxic compounds and bioactive ingredients. Biomed. Pharmacother. 2006, 60, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Dangles, O.; Dufour, C.; Manach, C.; Morand, C.; Remesy, C. Binding of flavonoids to plasma proteins. Methods Enzymol. 2001, 335, 319–333. [Google Scholar] [PubMed]




© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kőszegi, T.; Poór, M. Ochratoxin A: Molecular Interactions, Mechanisms of Toxicity and Prevention at the Molecular Level. Toxins 2016, 8, 111. https://doi.org/10.3390/toxins8040111
Kőszegi T, Poór M. Ochratoxin A: Molecular Interactions, Mechanisms of Toxicity and Prevention at the Molecular Level. Toxins. 2016; 8(4):111. https://doi.org/10.3390/toxins8040111
Chicago/Turabian StyleKőszegi, Tamás, and Miklós Poór. 2016. "Ochratoxin A: Molecular Interactions, Mechanisms of Toxicity and Prevention at the Molecular Level" Toxins 8, no. 4: 111. https://doi.org/10.3390/toxins8040111
APA StyleKőszegi, T., & Poór, M. (2016). Ochratoxin A: Molecular Interactions, Mechanisms of Toxicity and Prevention at the Molecular Level. Toxins, 8(4), 111. https://doi.org/10.3390/toxins8040111