
The Regulatory Networks That Control Clostridium difficile Toxin Synthesis

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
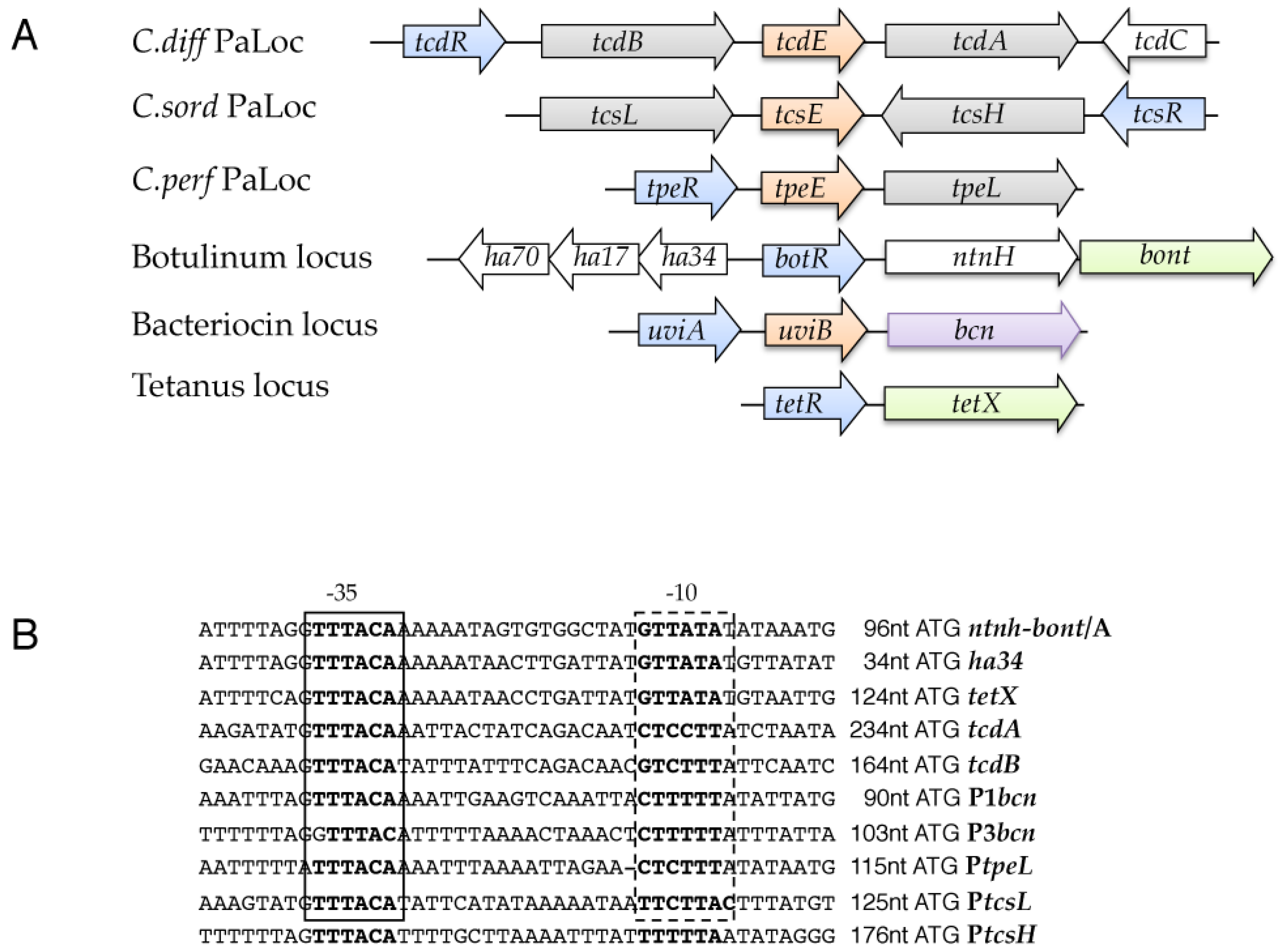
2. The Pathogenicity Loci of C. difficile (PaLoc and CdtLoc)
3. Toxin A and B Genes Are Specifically Transcribed by an Alternative Sigma Factor
4. Toxin Gene Transcription Is Negatively Controlled by TcdC
5. Toxin Expression Is Controlled by the Carbon Catabolite Repression (CCR) System
6. Regulation of Toxin Synthesis in Response to Proline and Cysteine Availability
7. Regulation of Toxin Synthesis by Nutritional Limitation Is Mediated by CodY
8. The Regulatory Network That Controls Transition Phase, Sporulation and Toxin Production in C. difficile
9. Other Regulators That Control Toxin Production
9.1. Toxin Gene Expression Is Controlled by Quorum Sensing
9.2. Control of Toxin Expression by c-di-GMP
9.3. Regulation of Toxin Expression by Flagellar Proteins
9.4. Impact of the Presence of Prophages on Toxin Gene Expression
9.5. Links between the Stress Response and Toxin Gene Expression
10. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Collins, M.D.; Lawson, P.A.; Willems, A.; Cordoba, J.J.; Fernandez-Garayzabal, J.; Garcia, P.; Cai, J.; Hippe, H.; Farrow, J.A. The phylogeny of the genus Clostridium: Proposal of five new genera and eleven new species combinations. Int. J. Syst. Bacteriol. 1994, 44, 812–826. [Google Scholar] [CrossRef] [PubMed]
- Stevens, D.L.; Aldape, M.J.; Bryant, A.E. Life-threatening clostridial infections. Anaerobe 2012, 18, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Yutin, N.; Galperin, M.Y. A genomic update on clostridial phylogeny: Gram-negative spore formers and other misplaced clostridia. Environ. Microbiol. 2013, 15, 2631–2641. [Google Scholar] [CrossRef] [PubMed]
- Borriello, S.P. Pathogenesis of Clostridium difficile infection. J. Antimicrob. Chemother. 1998, 41 (Suppl. SC), 13–19. [Google Scholar] [CrossRef] [PubMed]
- Rupnik, M.; Wilcox, M.H.; Gerding, D.N. Clostridium difficile infection: New developments in epidemiology and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Just, I.; Selzer, J.; Wilm, M.; von Eichel-Streiber, C.; Mann, M.; Aktories, K. Glucosylation of Rho proteins by Clostridium difficile toxin B. Nature 1995, 375, 500–503. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.P.; Rood, J.I.; Lyras, D. The role of toxin A and toxin B in the virulence of Clostridium difficile. Trends Microbiol. 2012, 20, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Eckert, C.; Emirian, A.; Le Monnier, A.; Cathala, L.; de Montclos, H.; Goret, J.; Berger, P.; Petit, A.; de Chevigny, A.; Jean-Pierre, H.; et al. Prevalence and pathogenicity of binary toxin-positive Clostridium difficile strains that do not produce toxins A and B. New Microbes New Infect. 2015, 3, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Gerding, D.N.; Johnson, S.; Rupnik, M.; Aktories, K. Clostridium difficile binary toxin CDT: Mechanism, epidemiology, and potential clinical importance. Gut Microbes 2014, 5, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Papatheodorou, P.; Carette, J.E.; Bell, G.W.; Schwan, C.; Guttenberg, G.; Brummelkamp, T.R.; Aktories, K. Lipolysis-stimulated lipoprotein receptor (LSR) is the host receptor for the binary toxin Clostridium difficile transferase (CDT). Proc. Natl. Acad. Sci. USA 2011, 108, 16422–16427. [Google Scholar] [CrossRef] [PubMed]
- Barth, H.; Aktories, K.; Popoff, M.R.; Stiles, B.G. Binary bacterial toxins: Biochemistry, biology, and applications of common Clostridium and Bacillus proteins. Microbiol. Mol. Biol. Rev. 2004, 68, 373–402. [Google Scholar] [CrossRef] [PubMed]
- Aktories, K.; Schwan, C.; Papatheodorou, P.; Lang, A.E. Bidirectional attack on the actin cytoskeleton. Bacterial protein toxins causing polymerization or depolymerization of actin. Toxicon 2012, 60, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Schwan, C.; Nolke, T.; Kruppke, A.S.; Schubert, D.M.; Lang, A.E.; Aktories, K. Cholesterol- and sphingolipid-rich microdomains are essential for microtubule-based membrane protrusions induced by Clostridium difficile transferase (CDT). J. Biol. Chem. 2011, 286, 29356–29365. [Google Scholar] [CrossRef] [PubMed]
- Schwan, C.; Stecher, B.; Tzivelekidis, T.; van Ham, M.; Rohde, M.; Hardt, W.D.; Wehland, J.; Aktories, K. Clostridium difficile toxin CDT induces formation of microtubule-based protrusions and increases adherence of bacteria. PLoS Pathog. 2009, 5, e1000626. [Google Scholar] [CrossRef] [PubMed]
- Akerlund, T.; Svenungsson, B.; Lagergren, A.; Burman, L.G. Correlation of disease severity with fecal toxin levels in patients with Clostridium difficile-associated diarrhea and distribution of PCR ribotypes and toxin yields in vitro of corresponding isolates. J. Clin. Microbiol. 2006, 44, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Wren, B.; Heard, S.R.; Tabaqchali, S. Association between production of toxins A and B and types of Clostridium difficile. J. Clin. Pathol. 1987, 40, 1397–1401. [Google Scholar] [CrossRef] [PubMed]
- Warny, M.; Pepin, J.; Fang, A.; Killgore, G.; Thompson, A.; Brazier, J.; Frost, E.; McDonald, L.C. Toxin production by an emerging strain of Clostridium difficile associated with outbreaks of severe disease in North America and Europe. Lancet 2005, 366, 1079–1084. [Google Scholar] [CrossRef]
- Merrigan, M.; Venugopal, A.; Mallozzi, M.; Roxas, B.; Viswanathan, V.K.; Johnson, S.; Gerding, D.N.; Vedantam, G. Human hypervirulent Clostridium difficile strains exhibit increased sporulation as well as robust toxin production. J. Bacteriol. 2010, 192, 4904–4911. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, B.; Sonenshein, A.L. Regulated transcription of Clostridium difficile toxin genes. Mol. Microbiol. 1998, 27, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Hundsberger, T.; Braun, V.; Weidmann, M.; Leukel, P.; Sauerborn, M.; von Eichel-Streiber, C. Transcription analysis of the genes tcdA-E of the pathogenicity locus of Clostridium difficile. Eur. J. Biochem. 1997, 244, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, K.; Karasawa, T.; Ikoma, S.; Nakamura, S. Enhancement of Clostridium difficile toxin production in biotin-limited conditions. J. Med. Microbiol. 1996, 44, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, S.; Burman, L.G.; Akerlund, T. Suppression of toxin production in Clostridium difficile VPI 10463 by amino acids. Microbiology 1999, 145 (Pt 7), 1683–1693. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, S.; Burman, L.G.; Akerlund, T. Induction of toxins in Clostridium difficile is associated with dramatic changes of its metabolism. Microbiology 2008, 154, 3430–3436. [Google Scholar] [CrossRef] [PubMed]
- Antunes, A.; Martin-Verstraete, I.; Dupuy, B. CcpA-mediated repression of Clostridium difficile toxin gene expression. Mol. Microbiol. 2011, 79, 882–899. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, S.; Lindberg, A.; Norin, E.; Burman, L.G.; Akerlund, T. Toxins, butyric acid, and other short-chain fatty acids are coordinately expressed and down-regulated by cysteine in Clostridium difficile. Infect. Immun. 2000, 68, 5881–5888. [Google Scholar] [CrossRef] [PubMed]
- Bouillaut, L.; Self, W.T.; Sonenshein, A.L. Proline-dependent regulation of Clostridium difficile Stickland metabolism. J. Bacteriol. 2013, 195, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, S.; Dupuy, B.; Mukherjee, K.; Norin, E.; Burman, L.G.; Akerlund, T. Expression of Clostridium difficile toxins A and B and their sigma factor TcdD is controlled by temperature. Infect. Immun. 2003, 71, 1784–1793. [Google Scholar] [CrossRef] [PubMed]
- Onderdonk, A.B.; Lowe, B.R.; Bartlett, J.G. Effect of environmental stress on Clostridium difficile toxin levels during continuous cultivation. Appl. Environ. Microbiol. 1979, 38, 637–641. [Google Scholar] [PubMed]
- Honda, T.; Hernadez, I.; Katoh, T.; Miwatani, T. Stimulation of enterotoxin production of Clostridium difficile by antibiotics. Lancet 1983, 1, 655. [Google Scholar] [CrossRef]
- Barc, M.C.; Depitre, C.; Corthier, G.; Collignon, A.; Su, W.J.; Bourlioux, P. Effects of antibiotics and other drugs on toxin production in Clostridium difficile in vitro and in vivo. Antimicrob. Agents Chemother. 1992, 36, 1332–1335. [Google Scholar] [CrossRef] [PubMed]
- Drummond, L.J.; Smith, D.G.; Poxton, I.R. Effects of sub-MIC concentrations of antibiotics on growth of and toxin production by Clostridium difficile. J. Med. Microbiol. 2003, 52, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Aldape, M.J.; Packham, A.E.; Nute, D.W.; Bryant, A.E.; Stevens, D.L. Effects of ciprofloxacin on the expression and production of exotoxins by Clostridium difficile. J. Med. Microbiol. 2013, 62, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Chilton, C.H.; Freeman, J.; Crowther, G.S.; Todhunter, S.L.; Nicholson, S.; Wilcox, M.H. Co-amoxiclav induces proliferation and cytotoxin production of Clostridium difficile ribotype 027 in a human gut model. J. Antimicrob. Chemother. 2012, 67, 951–954. [Google Scholar] [CrossRef] [PubMed]
- Gerber, M.; Walch, C.; Loffler, B.; Tischendorf, K.; Reischl, U.; Ackermann, G. Effect of sub-MIC concentrations of metronidazole, vancomycin, clindamycin and linezolid on toxin gene transcription and production in Clostridium difficile. J. Med. Microbiol. 2008, 57, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Braun, V.; Hundsberger, T.; Leukel, P.; Sauerborn, M.; von Eichel-Streiber, C. Definition of the single integration site of the pathogenicity locus in Clostridium difficile. Gene 1996, 181, 29–38. [Google Scholar] [CrossRef]
- Mani, N.; Dupuy, B. Regulation of toxin synthesis in Clostridium difficile by an alternative RNA polymerase sigma factor. Proc. Natl. Acad. Sci. USA 2001, 98, 5844–5849. [Google Scholar] [CrossRef] [PubMed]
- Matamouros, S.; England, P.; Dupuy, B. Clostridium difficile toxin expression is inhibited by the novel regulator TcdC. Mol. Microbiol. 2007, 64, 1274–1288. [Google Scholar] [CrossRef] [PubMed]
- Govind, R.; Dupuy, B. Secretion of Clostridium difficile toxins A and B requires the holin-like protein TcdE. PLoS Pathog. 2012, 8, e1002727. [Google Scholar] [CrossRef] [PubMed]
- Dingle, K.E.; Elliott, B.; Robinson, E.; Griffiths, D.; Eyre, D.W.; Stoesser, N.; Vaughan, A.; Golubchik, T.; Fawley, W.N.; Wilcox, M.H.; et al. Evolutionary history of the Clostridium difficile pathogenicity locus. Genome Biol. Evol. 2014, 6, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Janezic, S.; Marin, M.; Martin, A.; Rupnik, M. A new type of toxin A-negative, toxin B-positive Clostridium difficile strain lacking a complete tcdA gene. J. Clin. Microbiol. 2015, 53, 692–695. [Google Scholar] [CrossRef] [PubMed]
- Monot, M.; Eckert, C.; Lemire, A.; Hamiot, A.; Dubois, T.; Tessier, C.; Dumoulard, B.; Hamel, B.; Petit, A.; Lalande, V.; et al. Clostridium difficile: New Insights into the Evolution of the Pathogenicity Locus. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, M.S.; Roberts, A.P.; Hussain, H.; Williams, R.J.; Allan, E.; Mullany, P. Horizontal gene transfer converts non-toxigenic Clostridium difficile strains into toxin producers. Nat. Commun. 2013, 4, 2601. [Google Scholar] [CrossRef] [PubMed]
- Sirigi Reddy, A.R.; Girinathan, B.P.; Zapotocny, R.; Govind, R. Identification and characterization of Clostridium sordellii toxin gene regulator. J. Bacteriol. 2013, 195, 4246–4254. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.P.; Larcombe, S.; Li, L.; Jayawardena, D.; Awad, M.M.; Songer, J.G.; Lyras, D. Expression of the large clostridial toxins is controlled by conserved regulatory mechanisms. Int J. Med. Microbiol. 2014, 304, 1147–1159. [Google Scholar] [CrossRef] [PubMed]
- Von Eichel-Streiber, C.; Harperath, U.; Bosse, D.; Hadding, U. Purification of two high molecular weight toxins of Clostridium difficile which are antigenically related. Microb. Pathog. 1987, 2, 307–318. [Google Scholar] [CrossRef]
- Hammond, G.A.; Lyerly, D.M.; Johnson, J.L. Transcriptional analysis of the toxigenic element of Clostridium difficile. Microb. Pathog. 1997, 22, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.P.; Lyras, D.; Allen, D.L.; Mackin, K.E.; Howarth, P.M.; O’Connor, J.R.; Rood, J.I. Binary toxin production in Clostridium difficile is regulated by CdtR, a LytTR family response regulator. J. Bacteriol. 2007, 189, 7290–7301. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.P.; Mackin, K.E.; Rood, J.I.; Lyras, D. Regulation of toxin production in Clostridium difficile. In Regulation of Bacterial Virulence; Vasil, M., Darwin, A., Eds.; ASM Press: Washington, DC, USA, 2013; pp. 295–306. [Google Scholar]
- Moncrief, J.S.; Barroso, L.A.; Wilkins, T.D. Positive regulation of Clostridium difficile toxins. Infect. Immun. 1997, 65, 1105–1108. [Google Scholar] [PubMed]
- Mani, N.; Lyras, D.; Barroso, L.; Howarth, P.; Wilkins, T.; Rood, J.I.; Sonenshein, A.L.; Dupuy, B. Environmental response and autoregulation of Clostridium difficile TxeR, a sigma factor for toxin gene expression. J. Bacteriol. 2002, 184, 5971–5978. [Google Scholar] [CrossRef] [PubMed]
- Raffestin, S.; Dupuy, B.; Marvaud, J.C.; Popoff, M.R. BotR/A and TetR are alternative RNA polymerase sigma factors controlling the expression of the neurotoxin and associated protein genes in Clostridium botulinum type A and Clostridium tetani. Mol. Microbiol. 2005, 55, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, B.; Raffestin, S.; Matamouros, S.; Mani, N.; Popoff, M.R.; Sonenshein, A.L. Regulation of toxin and bacteriocin gene expression in Clostridium by interchangeable RNA polymerase sigma factors. Mol. Microbiol. 2006, 60, 1044–1057. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, B.; Matamouros, S. Regulation of toxin and bacteriocin synthesis in Clostridium species by a new subgroup of RNA polymerase sigma-factors. Res. Microbiol. 2006, 157, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Finn, C.W., Jr.; Silver, R.P.; Habig, W.H.; Hardegree, M.C.; Zon, G.; Garon, C.F. The structural gene for tetanus neurotoxin is on a plasmid. Science 1984, 224, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Garnier, T.; Cole, S.T. Characterization of a bacteriocinogenic plasmid from Clostridium perfringens and molecular genetic analysis of the bacteriocin-encoding gene. J. Bacteriol. 1986, 168, 1189–1196. [Google Scholar] [PubMed]
- Couchman, E.C.; Browne, H.P.; Dunn, M.; Lawley, T.D.; Songer, J.G.; Hall, V.; Petrovska, L.; Vidor, C.; Awad, M.; Lyras, D.; et al. Clostridium sordellii genome analysis reveals plasmid localized toxin genes encoded within pathogenicity loci. BMC Genom. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Gurjar, A.; Li, J.; McClane, B.A. Characterization of toxin plasmids in Clostridium perfringens type C isolates. Infect. Immun. 2010, 78, 4860–4869. [Google Scholar] [CrossRef] [PubMed]
- Monot, M.; Boursaux-Eude, C.; Thibonnier, M.; Vallenet, D.; Moszer, I.; Medigue, C.; Martin-Verstraete, I.; Dupuy, B. Reannotation of the genome sequence of Clostridium difficile strain 630. J. Med. Microbiol. 2011, 60, 1193–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eklund, M.W.; Poysky, F.T.; Reed, S.M.; Smith, C.A. Bacteriophage and the toxigenicity of Clostridium botulinum type C. Science 1971, 172, 480–482. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, B.; Mani, N.; Katayama, S.; Sonenshein, A.L. Transcription activation of a UV-inducible Clostridium perfringens bacteriocin gene by a novel sigma factor. Mol. Microbiol. 2005, 55, 1196–1206. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, M.; Campbell, K.D.; Collins, M.D.; East, A.K. Molecular characterization of the clusters of genes encoding the botulinum neurotoxin complex in Clostridium botulinum (Clostridium argentinense) type G and nonproteolytic Clostridium botulinum type B. Curr. Microbiol. 1997, 35, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Henderson, I.; Whelan, S.M.; Davis, T.O.; Minton, N.P. Genetic characterisation of the botulinum toxin complex of Clostridium botulinum strain NCTC 2916. FEMS Microbiol. Lett. 1996, 140, 151–158. [Google Scholar] [CrossRef] [PubMed]
- MacCannell, D.R.; Louie, T.J.; Gregson, D.B.; Laverdiere, M.; Labbe, A.C.; Laing, F.; Henwick, S. Molecular analysis of Clostridium difficile PCR ribotype 027 isolates from Eastern and Western Canada. J. Clin. Microbiol. 2006, 44, 2147–2152. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, B.; Govind, R.; Antunes, A.; Matamouros, S. Clostridium difficile toxin synthesis is negatively regulated by TcdC. J. Med. Microbiol. 2008, 57, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Govind, R.; Vediyappan, G.; Rolfe, R.D.; Fralick, J.A. Evidence that Clostridium difficile TcdC is a membrane-associated protein. J. Bacteriol. 2006, 188, 3716–3720. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, H.C.; Bakker, D.; Steindel, P.; Kuijper, E.J.; Corver, J. Clostridium difficile TcdC protein binds four-stranded G-quadruplex structures. Nucleic Acids Res. 2013, 41, 2382–2393. [Google Scholar] [CrossRef] [PubMed]
- Lipps, H.J.; Rhodes, D. G-quadruplex structures: In vivo evidence and function. Trends Cell Biol. 2009, 19, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Curry, S.R.; Marsh, J.W.; Muto, C.A.; O’Leary, M.M.; Pasculle, A.W.; Harrison, L.H. tcdC genotypes associated with severe TcdC truncation in an epidemic clone and other strains of Clostridium difficile. J. Clin. Microbiol. 2007, 45, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.; Boyd, D.; Levett, P.N.; Mulvey, M.R.; Alfa, M.J. Truncation in the tcdC region of the Clostridium difficile PathLoc of clinical isolates does not predict increased biological activity of Toxin B or Toxin A. BMC Infect. Dis. 2009, 9. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, S.D.; French, G.L. Lack of association of tcdC type and binary toxin status with disease severity and outcome in toxigenic Clostridium difficile. J. Infect. 2011, 62, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.P.; Douce, G.R.; Govind, R.; Howarth, P.M.; Mackin, K.E.; Spencer, J.; Buckley, A.M.; Antunes, A.; Kotsanas, D.; Jenkin, G.A.; et al. The anti-sigma factor TcdC modulates hypervirulence in an epidemic BI/NAP1/027 clinical isolate of Clostridium difficile. PLoS Pathog. 2011, 7, e1002317. [Google Scholar] [CrossRef] [PubMed]
- Cartman, S.T.; Kelly, M.L.; Heeg, D.; Heap, J.T.; Minton, N.P. Precise manipulation of the Clostridium difficile chromosome reveals a lack of association between the tcdC genotype and toxin production. Appl. Environ. Microbiol. 2012, 78, 4683–4690. [Google Scholar] [CrossRef] [PubMed]
- Bakker, D.; Smits, W.K.; Kuijper, E.J.; Corver, J. TcdC does not significantly repress toxin expression in Clostridium difficile 630DeltaErm. PLoS ONE 2012, 7, e43247. [Google Scholar] [CrossRef] [PubMed]
- Neumann-Schaal, M.; Hofmann, J.D.; Will, S.E.; Schomburg, D. Time-resolved amino acid uptake of Clostridium difficile 630Deltaerm and concomitant fermentation product and toxin formation. BMC Microbiol. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Kazamias, M.T.; Sperry, J.F. Enhanced fermentation of mannitol and release of cytotoxin by Clostridium difficile in alkaline culture media. Appl. Environ. Microbiol. 1995, 61, 2425–2427. [Google Scholar] [PubMed]
- Stulke, J.; Hillen, W. Regulation of carbon catabolism in Bacillus species. Annu. Rev. Microbiol. 2000, 54, 849–880. [Google Scholar] [CrossRef] [PubMed]
- Deutscher, J.; Francke, C.; Postma, P.W. How phosphotransferase system-related protein phosphorylation regulates carbohydrate metabolism in bacteria. Microbiol. Mol. Biol. Rev. 2006, 70, 939–1031. [Google Scholar] [CrossRef] [PubMed]
- Deutscher, J.; Saier, M.H., Jr. ATP-dependent protein kinase-catalyzed phosphorylation of a seryl residue in HPr, a phosphate carrier protein of the phosphotransferase system in Streptococcus pyogenes. Proc. Natl. Acad. Sci. USA 1983, 80, 6790–6794. [Google Scholar] [CrossRef] [PubMed]
- Antunes, A.; Camiade, E.; Monot, M.; Courtois, E.; Barbut, F.; Sernova, N.V.; Rodionov, D.A.; Martin-Verstraete, I.; Dupuy, B. Global transcriptional control by glucose and carbon regulator CcpA in Clostridium difficile. Nucleic Acids Res. 2012, 40, 10701–10718. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Stirewalt, V.L.; Melville, S.B. The CcpA protein is necessary for efficient sporulation and enterotoxin gene (cpe) regulation in Clostridium perfringens. J. Bacteriol. 2004, 186, 5221–5229. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.; Calos, M.; Myers, A.; Self, W.T. Analysis of proline reduction in the nosocomial pathogen Clostridium difficile. J. Bacteriol. 2006, 188, 8487–8495. [Google Scholar] [CrossRef] [PubMed]
- Bouillaut, L.; Dubois, T.; Francis, M.B.; Daou, N.; Monot, M.; Sorg, J.A.; Dupuy, B.; Sonenshein, A.L. Role of the global regulator Rex in control of NAD+-regeneration in Clostridium difficile. Mol. Mic. manuscript in preparation.
- Pagels, M.; Fuchs, S.; Pane-Farre, J.; Kohler, C.; Menschner, L.; Hecker, M.; McNamarra, P.J.; Bauer, M.C.; von Wachenfeldt, C.; Liebeke, M.; et al. Redox sensing by a Rex-family repressor is involved in the regulation of anaerobic gene expression in Staphylococcus aureus. Mol. Microbiol. 2010, 76, 1142–1161. [Google Scholar] [CrossRef] [PubMed]
- Brekasis, D.; Paget, M.S. A novel sensor of NADH/NAD+ redox poise in Streptomyces coelicolor A3 (2). EMBO J. 2003, 22, 4856–4865. [Google Scholar] [CrossRef] [PubMed]
- Sickmier, E.A.; Brekasis, D.; Paranawithana, S.; Bonanno, J.B.; Paget, M.S.; Burley, S.K.; Kielkopf, C.L. X-ray structure of a Rex-family repressor/NADH complex insights into the mechanism of redox sensing. Structure 2005, 13, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Wietzke, M.; Bahl, H. The redox-sensing protein Rex, a transcriptional regulator of solventogenesis in Clostridium acetobutylicum. Appl. Microbiol. Biotechnol. 2012, 96, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Gyan, S.; Shiohira, Y.; Sato, I.; Takeuchi, M.; Sato, T. Regulatory loop between redox sensing of the NADH/NAD+ ratio by Rex (YdiH) and oxidation of NADH by NADH dehydrogenase Ndh in Bacillus subtilis. J. Bacteriol. 2006, 188, 7062–7071. [Google Scholar] [CrossRef] [PubMed]
- Schau, M.; Chen, Y.; Hulett, F.M. Bacillus subtilis YdiH is a direct negative regulator of the cydABCD operon. J. Bacteriol. 2004, 186, 4585–4595. [Google Scholar] [CrossRef] [PubMed]
- Ravcheev, D.A.; Li, X.; Latif, H.; Zengler, K.; Leyn, S.A.; Korostelev, Y.D.; Kazakov, A.E.; Novichkov, P.S.; Osterman, A.L.; Rodionov, D.A. Transcriptional regulation of central carbon and energy metabolism in bacteria by redox-responsive repressor Rex. J. Bacteriol. 2012, 194, 1145–1157. [Google Scholar] [CrossRef] [PubMed]
- Andre, G.; Haudecoeur, E.; Monot, M.; Ohtani, K.; Shimizu, T.; Dupuy, B.; Martin-Verstraete, I. Global regulation of gene expression in response to cysteine availability in Clostridium perfringens. BMC Microbiol. 2010, 10. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, J.A.; Nazario-Larrieu, J.; Sarwar, J.; Alexander, P.; Blake, M.S. Bordetella pertussis autoregulates pertussis toxin production through the metabolism of cysteine. Infect. Immun. 2001, 69, 6823–6830. [Google Scholar] [CrossRef] [PubMed]
- Dubois, T.; Dancer-Thibonnier, M.; Monot, M.; Hamiot, A.; Bouillaut, L.; Soutourina, O.; Martin-Verstraete, I.; Dupuy, B. Control of Clostridium difficile physiopathology in response to cysteine availability. Infect. Immun. submitted for publication. 2016. [Google Scholar]
- Dineen, S.S.; Villapakkam, A.C.; Nordman, J.T.; Sonenshein, A.L. Repression of Clostridium difficile toxin gene expression by CodY. Mol. Microbiol. 2007, 66, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Saujet, L.; Monot, M.; Dupuy, B.; Soutourina, O.; Martin-Verstraete, I. The key sigma factor of transition phase, SigH, controls sporulation, metabolism, and virulence factor expression in Clostridium difficile. J. Bacteriol. 2011, 193, 3186–3196. [Google Scholar] [CrossRef] [PubMed]
- Dalet, K.; Briand, C.; Cenatiempo, Y.; Hechard, Y. The rpoN gene of Enterococcus faecalis directs sensitivity to subclass IIa bacteriocins. Curr. Microbiol. 2000, 41, 441–443. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Okada, N.; Makino, S.; Asakura, H.; Yamamoto, S.; Igimi, S. The sigma factor RpoN (sigma54) is involved in osmotolerance in Listeria monocytogenes. FEMS Microbiol. Lett. 2006, 263, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Molle, V.; Nakaura, Y.; Shivers, R.P.; Yamaguchi, H.; Losick, R.; Fujita, Y.; Sonenshein, A.L. Additional targets of the Bacillus subtilis global regulator CodY identified by chromatin immunoprecipitation and genome-wide transcript analysis. J. Bacteriol. 2003, 185, 1911–1922. [Google Scholar] [CrossRef] [PubMed]
- Bergara, F.; Ibarra, C.; Iwamasa, J.; Patarroyo, J.C.; Aguilera, R.; Marquez-Magana, L.M. CodY is a nutritional repressor of flagellar gene expression in Bacillus subtilis. J. Bacteriol. 2003, 185, 3118–3126. [Google Scholar] [CrossRef] [PubMed]
- Ratnayake-Lecamwasam, M.; Serror, P.; Wong, K.W.; Sonenshein, A.L. Bacillus subtilis CodY represses early-stationary-phase genes by sensing GTP levels. Genes Dev. 2001, 15, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Sonenshein, A.L. CodY, a global regulator of stationary phase and virulence in Gram-positive bacteria. Curr. Opin. Microbiol. 2005, 8, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Shivers, R.P.; Sonenshein, A.L. Activation of the Bacillus subtilis global regulator CodY by direct interaction with branched-chain amino acids. Mol. Microbiol. 2004, 53, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Dineen, S.S.; McBride, S.M.; Sonenshein, A.L. Integration of metabolism and virulence by Clostridium difficile CodY. J. Bacteriol. 2010, 192, 5350–5362. [Google Scholar] [CrossRef] [PubMed]
- Bordeleau, E.; Fortier, L.C.; Malouin, F.; Burrus, V. c-di-GMP turn-over in Clostridium difficile is controlled by a plethora of diguanylate cyclases and phosphodiesterases. PLoS Genet. 2011, 7, e1002039. [Google Scholar] [CrossRef] [PubMed]
- Koenigsknecht, M.J.; Theriot, C.M.; Bergin, I.L.; Schumacher, C.A.; Schloss, P.D.; Young, V.B. Dynamics and establishment of Clostridium difficile infection in the murine gastrointestinal tract. Infect. Immun. 2015, 83, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Pettit, L.J.; Browne, H.P.; Yu, L.; Smits, W.K.; Fagan, R.P.; Barquist, L.; Martin, M.J.; Goulding, D.; Duncan, S.H.; Flint, H.J.; et al. Functional genomics reveals that Clostridium difficile Spo0A coordinates sporulation, virulence and metabolism. BMC Genom. 2014, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackin, K.E.; Carter, G.P.; Howarth, P.; Rood, J.I.; Lyras, D. Spo0A Differentially Regulates Toxin Production in Evolutionarily Diverse Strains of Clostridium difficile. PLoS ONE 2013, 8, e79666. [Google Scholar] [CrossRef] [PubMed]
- Deakin, L.J.; Clare, S.; Fagan, R.P.; Dawson, L.F.; Pickard, D.J.; West, M.R.; Wren, B.W.; Fairweather, N.F.; Dougan, G.; Lawley, T.D. The Clostridium difficile spo0A gene is a persistence and transmission factor. Infect. Immun. 2012, 80, 2704–2711. [Google Scholar] [CrossRef] [PubMed]
- Sonenshein, A.L. Control of sporulation initiation in Bacillus subtilis. Curr. Opin. Microbiol. 2000, 3, 561–566. [Google Scholar] [CrossRef]
- Edwards, A.N.; McBride, S.M. Initiation of sporulation in Clostridium difficile: A twist on the classic model. FEMS Microbiol. Lett. 2014, 358, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Underwood, S.; Guan, S.; Vijayasubhash, V.; Baines, S.D.; Graham, L.; Lewis, R.J.; Wilcox, M.H.; Stephenson, K. Characterization of the sporulation initiation pathway of Clostridium difficile and its role in toxin production. J. Bacteriol. 2009, 191, 7296–7305. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.; Dworkin, J. Recent progress in Bacillus subtilis sporulation. FEMS Microbiol. Rev. 2012, 36, 131–148. [Google Scholar] [CrossRef] [PubMed]
- Phillips, Z.E.; Strauch, M.A. Bacillus subtilis sporulation and stationary phase gene expression. Cell. Mol. Life Sci. 2002, 59, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Britton, R.A.; Eichenberger, P.; Gonzalez-Pastor, J.E.; Fawcett, P.; Monson, R.; Losick, R.; Grossman, A.D. Genome-wide analysis of the stationary-phase sigma factor (Sigma-H) regulon of Bacillus subtilis. J. Bacteriol. 2002, 184, 4881–4890. [Google Scholar] [CrossRef] [PubMed]
- Paredes, C.J.; Alsaker, K.V.; Papoutsakis, E.T. A comparative genomic view of clostridial sporulation and physiology. Nat. Rev. Microbiol. 2005, 3, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Steiner, E.; Dago, A.E.; Young, D.I.; Heap, J.T.; Minton, N.P.; Hoch, J.A.; Young, M. Multiple orphan histidine kinases interact directly with Spo0A to control the initiation of endospore formation in Clostridium acetobutylicum. Mol. Microbiol. 2011, 80, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Molle, V.; Fujita, M.; Jensen, S.T.; Eichenberger, P.; Gonzalez-Pastor, J.E.; Liu, J.S.; Losick, R. The Spo0A regulon of Bacillus subtilis. Mol. Microbiol. 2003, 50, 1683–1701. [Google Scholar] [CrossRef] [PubMed]
- Rosenbusch, K.E.; Bakker, D.; Kuijper, E.J.; Smits, W.K. C. difficile 630Deltaerm Spo0A regulates sporulation, but does not contribute to toxin production, by direct high-affinity binding to target DNA. PLoS ONE 2012, 7, e48608. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.N.; Tamayo, R.; McBride, S.M. A Novel Regulator Controls Clostridium difficile Sporulation, Motility and Toxin Production. Mol. Microbiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Slamti, L.; Perchat, S.; Huillet, E.; Lereclus, D. Quorum sensing in Bacillus thuringiensis is required for completion of a full infectious cycle in the insect. Toxins 2014, 6, 2239–2255. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, S.; Borriello, S.P. A non-haemagglutinating form of Clostridium difficile toxin A. J. Med. Microbiol. 1992, 36, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Harry, K.H.; Zhou, R.; Kroos, L.; Melville, S.B. Sporulation and enterotoxin (CPE) synthesis are controlled by the sporulation-specific sigma factors SigE and SigK in Clostridium perfringens. J. Bacteriol. 2009, 191, 2728–2742. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Melville, S.B. Identification and characterization of sporulation-dependent promoters upstream of the enterotoxin gene (cpe) of Clostridium perfringens. J. Bacteriol. 1998, 180, 136–142. [Google Scholar] [PubMed]
- Fimlaid, K.A.; Bond, J.P.; Schutz, K.C.; Putnam, E.E.; Leung, J.M.; Lawley, T.D.; Shen, A. Global analysis of the sporulation pathway of Clostridium difficile. PLoS Genet. 2013, 9, e1003660. [Google Scholar] [CrossRef] [PubMed]
- Saujet, L.; Pereira, F.C.; Serrano, M.; Soutourina, O.; Monot, M.; Shelyakin, P.V.; Gelfand, M.S.; Dupuy, B.; Henriques, A.O.; Martin-Verstraete, I. Genome-Wide Analysis of Cell Type-Specific Gene Transcription during Spore Formation in Clostridium difficile. PLoS Genet. 2013, 9, e1003756. [Google Scholar] [CrossRef] [PubMed]
- Lawley, T.D.; Croucher, N.J.; Yu, L.; Clare, S.; Sebaihia, M.; Goulding, D.; Pickard, D.J.; Parkhill, J.; Choudhary, J.; Dougan, G. Proteomic and genomic characterization of highly infectious Clostridium difficile 630 spores. J. Bacteriol. 2009, 191, 5377–5386. [Google Scholar] [CrossRef] [PubMed]
- Abhyankar, W.; Hossain, A.H.; Djajasaputra, A.; Permpoonpattana, P.; Ter Beek, A.; Dekker, H.L.; Cutting, S.M.; Brul, S.; de Koning, L.J.; de Koster, C.G. In pursuit of protein targets: Proteomic characterization of bacterial spore outer layers. J. Proteome Res. 2013, 12, 4507–4521. [Google Scholar] [CrossRef] [PubMed]
- Cassona, C.P.; Ramalhete, S.; Antunes, W.; Dupuy, B.; Serrano, M.; Henriques, A.O. The link between toxin production and spore formation in the intestinal pathogen Clostridium difficile. In Proceedings of the 9th international conference on the molecular biology and pathogenesis of the clostridia, Freiburg, Germany, 10 September 2015.
- Arya, R.; Princy, S.A. An insight into pleiotropic regulators Agr and Sar: Molecular probes paving the new way for antivirulent therapy. Future Microbiol. 2013, 8, 1339–1353. [Google Scholar] [CrossRef] [PubMed]
- Stabler, R.A.; He, M.; Dawson, L.; Martin, M.; Valiente, E.; Corton, C.; Lawley, T.D.; Sebaihia, M.; Quail, M.A.; Rose, G.; et al. Comparative genome and phenotypic analysis of Clostridium difficile 027 strains provides insight into the evolution of a hypervirulent bacterium. Genome Biol. 2009, 10. [Google Scholar] [CrossRef] [PubMed]
- Marsden, G.L.; Davis, I.J.; Wright, V.J.; Sebaihia, M.; Kuijper, E.J.; Minton, N.P. Array comparative hybridisation reveals a high degree of similarity between UK and European clinical isolates of hypervirulent Clostridium difficile. BMC Genom. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Darkoh, C.; DuPont, H.L.; Norris, S.J.; Kaplan, H.B. Toxin synthesis by Clostridium difficile is regulated through quorum signaling. MBio 2015, 6, e02569. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.J.; Clare, S.; Goulding, D.; Faulds-Pain, A.; Barquist, L.; Browne, H.P.; Pettit, L.; Dougan, G.; Lawley, T.D.; Wren, B.W. The agr locus regulates virulence and colonization genes in Clostridium difficile 027. J. Bacteriol. 2013, 195, 3672–3681. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; McClane, B.A. Role of the Agr-like quorum-sensing system in regulating toxin production by Clostridium perfringens type B strains CN1793 and CN1795. Infect. Immun. 2012, 80, 3008–3017. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Rood, J.I.; McClane, B.A. Epsilon-toxin production by Clostridium perfringens type D strain CN3718 is dependent upon the agr operon but not the VirS/VirR two-component regulatory system. MBio 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, J.; Vidal, J.E.; McClane, B.A. The Agr-like quorum-sensing system regulates sporulation and production of enterotoxin and beta2 toxin by Clostridium perfringens type A non-food-borne human gastrointestinal disease strain F5603. Infect. Immun. 2011, 79, 2451–2459. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Yuan, Y.; Hassan, S.; Wang, R.; Wang, Y.; Shimizu, T. Virulence gene regulation by the agr system in Clostridium perfringens. J. Bacteriol. 2009, 191, 3919–3927. [Google Scholar] [CrossRef] [PubMed]
- Vidal, J.E.; Ma, M.; Saputo, J.; Garcia, J.; Uzal, F.A.; McClane, B.A. Evidence that the Agr-like quorum sensing system regulates the toxin production, cytotoxicity and pathogenicity of Clostridium perfringens type C isolate CN3685. Mol. Microbiol. 2012, 83, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Steiner, E.; Scott, J.; Minton, N.P.; Winzer, K. An agr quorum sensing system that regulates granulose formation and sporulation in Clostridium acetobutylicum. Appl. Environ. Microbiol. 2012, 78, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- Cooksley, C.M.; Davis, I.J.; Winzer, K.; Chan, W.C.; Peck, M.W.; Minton, N.P. Regulation of neurotoxin production and sporulation by a Putative agrBD signaling system in proteolytic Clostridium botulinum. Appl. Environ. Microbiol. 2010, 76, 4448–4460. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.S.; Thompson, J.A.; Xavier, K.B. AI-2-mediated signalling in bacteria. FEMS Microbiol. Rev. 2013, 37, 156–181. [Google Scholar] [CrossRef] [PubMed]
- Hullo, M.F.; Auger, S.; Soutourina, O.; Barzu, O.; Yvon, M.; Danchin, A.; Martin-Verstraete, I. Conversion of methionine to cysteine in Bacillus subtilis and its regulation. J. Bacteriol. 2007, 189, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S.; Song, K.P. LuxS/autoinducer-2 quorum sensing molecule regulates transcriptional virulence gene expression in Clostridium difficile. Biochem. Biophys. Res. Commun. 2005, 335, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.P.; Purdy, D.; Williams, P.; Minton, N.P. Quorum sensing in Clostridium difficile: Analysis of a luxS-type signalling system. J. Med. Microbiol. 2005, 54, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.; Oh, S.; Song, M.; Hong, Y.S.; Park, S.; Park, D.J.; Griffiths, M.W.; Oh, S. Inhibitory Effect of Epigallocatechin Gallate on the Virulence of Clostridium difficile PCR Ribotype 027. J. Food Sci. 2015, 80, M2925–M2931. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Hayashi, H.; Shimizu, T. The luxS gene is involved in cell-cell signalling for toxin production in Clostridium perfringens. Mol. Microbiol. 2002, 44, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Romling, U.; Galperin, M.Y.; Gomelsky, M. Cyclic di-GMP: The first 25 years of a universal bacterial second messenger. Microbiol. Mol. Biol. Rev. 2013, 77, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Dong, X.; Subramanian, S.; Matthews, P.M.; Cooper, C.A.; Kearns, D.B.; Dann, C.E., 3rd. Engineering of Bacillus subtilis strains to allow rapid characterization of heterologous diguanylate cyclases and phosphodiesterases. Appl. Environ. Microbiol. 2014, 80, 6167–6174. [Google Scholar] [CrossRef] [PubMed]
- Soutourina, O.A.; Monot, M.; Boudry, P.; Saujet, L.; Pichon, C.; Sismeiro, O.; Semenova, E.; Severinov, K.; Le Bouguenec, C.; Coppee, J.Y.; et al. Genome-wide identification of regulatory RNAs in the human pathogen Clostridium difficile. PLoS Genet. 2013, 9, e1003493. [Google Scholar] [CrossRef] [PubMed]
- Peltier, J.; Shaw, H.A.; Couchman, E.C.; Dawson, L.F.; Yu, L.; Choudhary, J.S.; Kaever, V.; Wren, B.W.; Fairweather, N.F. Cyclic diGMP regulates production of sortase substrates of Clostridium difficile and their surface exposure through ZmpI protease-mediated cleavage. J. Biol. Chem. 2015, 290, 24453–24469. [Google Scholar] [CrossRef] [PubMed]
- Bordeleau, E.; Purcell, E.B.; Lafontaine, D.A.; Fortier, L.C.; Tamayo, R.; Burrus, V. Cyclic di-GMP riboswitch-regulated type IV pili contribute to aggregation of Clostridium difficile. J. Bacteriol. 2015, 197, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Purcell, E.B.; McKee, R.W.; Bordeleau, E.; Burrus, V.; Tamayo, R. Regulation of Type IV Pili Contributes to Surface Behaviors of Historical and Epidemic Strains of Clostridium difficile. J. Bacteriol. 2015, 198, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Purcell, E.B.; McKee, R.W.; McBride, S.M.; Waters, C.M.; Tamayo, R. Cyclic diguanylate inversely regulates motility and aggregation in Clostridium difficile. J. Bacteriol. 2012, 194, 3307–3316. [Google Scholar] [CrossRef] [PubMed]
- Hensbergen, P.J.; Klychnikov, O.I.; Bakker, D.; Dragan, I.; Kelly, M.L.; Minton, N.P.; Corver, J.; Kuijper, E.J.; Drijfhout, J.W.; van Leeuwen, H.C. Clostridium difficile secreted Pro-Pro endopeptidase PPEP-1 (ZMP1/CD2830) modulates adhesion through cleavage of the collagen binding protein CD2831. FEBS Lett. 2015, 589, 3952–3958. [Google Scholar] [CrossRef] [PubMed]
- McKee, R.W.; Mangalea, M.R.; Purcell, E.B.; Borchardt, E.K.; Tamayo, R. The second messenger cyclic Di-GMP regulates Clostridium difficile toxin production by controlling expression of sigD. J. Bacteriol. 2013, 195, 5174–5185. [Google Scholar] [CrossRef] [PubMed]
- El Meouche, I.; Peltier, J.; Monot, M.; Soutourina, O.; Pestel-Caron, M.; Dupuy, B.; Pons, J.L. Characterization of the SigD regulon of C. difficile and its positive control of toxin production through the regulation of tcdR. PLoS ONE 2013, 8, e83748. [Google Scholar]
- Aubry, A.; Hussack, G.; Chen, W.; KuoLee, R.; Twine, S.M.; Fulton, K.M.; Foote, S.; Carrillo, C.D.; Tanha, J.; Logan, S.M. Modulation of toxin production by the flagellar regulon in Clostridium difficile. Infect. Immun. 2012, 80, 3521–3532. [Google Scholar] [CrossRef] [PubMed]
- Baban, S.T.; Kuehne, S.A.; Barketi-Klai, A.; Cartman, S.T.; Kelly, M.L.; Hardie, K.R.; Kansau, I.; Collignon, A.; Minton, N.P. The role of flagella in Clostridium difficile pathogenesis: Comparison between a non-epidemic and an epidemic strain. PLoS ONE 2013, 8, e73026. [Google Scholar] [CrossRef] [PubMed]
- Dingle, T.C.; Mulvey, G.L.; Armstrong, G.D. Mutagenic analysis of the Clostridium difficile flagellar proteins, FliC and FliD, and their contribution to virulence in hamsters. Infect. Immun. 2011, 79, 4061–4067. [Google Scholar] [CrossRef] [PubMed]
- Twine, S.M.; Reid, C.W.; Aubry, A.; McMullin, D.R.; Fulton, K.M.; Austin, J.; Logan, S.M. Motility and flagellar glycosylation in Clostridium difficile. J. Bacteriol. 2009, 191, 7050–7062. [Google Scholar] [CrossRef] [PubMed]
- Brussow, H.; Hendrix, R.W. Phage genomics: Small is beautiful. Cell 2002, 108, 13–16. [Google Scholar] [CrossRef]
- Brussow, H.; Canchaya, C.; Hardt, W.D. Phages and the evolution of bacterial pathogens: From genomic rearrangements to lysogenic conversion. Microbiol. Mol. Biol. Rev. 2004, 68, 560–602. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, K.R.; Clokie, M.R. A Taxonomic Review of Clostridium difficile Phages and Proposal of a Novel Genus, “Phimmp04likevirus”. Viruses 2015, 7, 2534–2541. [Google Scholar] [CrossRef] [PubMed]
- Fortier, L.C.; Sekulovic, O. Importance of prophages to evolution and virulence of bacterial pathogens. Virulence 2013, 4, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Govind, R.; Vediyappan, G.; Rolfe, R.D.; Dupuy, B.; Fralick, J.A. Bacteriophage-mediated toxin gene regulation in Clostridium difficile. J. Virol. 2009, 83, 12037–12045. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.; Meader, E.; Mayer, M.; Narbad, A.; Roberts, A.P.; Mullany, P. Determination of the attP and attB sites of phage CD27 from Clostridium difficile NCTC 12727. J. Med. Microbiol. 2013, 62, 1439–1443. [Google Scholar] [CrossRef] [PubMed]
- Sekulovic, O.; Meessen-Pinard, M.; Fortier, L.C. Prophage-stimulated toxin production in Clostridium difficile NAP1/027 lysogens. J. Bacteriol. 2011, 193, 2726–2734. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.; Chang, B.J.; Riley, T.V. Effect of phage infection on toxin production by Clostridium difficile. J. Med. Microbiol. 2005, 54, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Erill, I.; Campoy, S.; Barbe, J. Aeons of distress: An evolutionary perspective on the bacterial SOS response. FEMS Microbiol. Rev. 2007, 31, 637–656. [Google Scholar] [CrossRef] [PubMed]
- Butala, M.; Zgur-Bertok, D.; Busby, S.J. The bacterial LexA transcriptional repressor. Cell. Mol. Life Sci. 2009, 66, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Walter, B.M.; Cartman, S.T.; Minton, N.P.; Butala, M.; Rupnik, M. The SOS Response Master Regulator LexA Is Associated with Sporulation, Motility and Biofilm Formation in Clostridium difficile. PLoS ONE 2015, 10, e0144763. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Ledesma, K.R.; Chang, K.T.; Tam, V.H. Impact of recA on levofloxacin exposure-related resistance development. Antimicrob. Agents Chemother. 2010, 54, 4262–4268. [Google Scholar] [CrossRef] [PubMed]
- Pultz, N.J.; Donskey, C.J. Effect of antibiotic treatment on growth of and toxin production by Clostridium difficile in the cecal contents of mice. Antimicrob. Agents Chemother. 2005, 49, 3529–3532. [Google Scholar] [CrossRef] [PubMed]
- Walter, B.M.; Rupnik, M.; Hodnik, V.; Anderluh, G.; Dupuy, B.; Paulic, N.; Zgur-Bertok, D.; Butala, M. The LexA regulated genes of the Clostridium difficile. BMC Microbiol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Selby, C.P.; Witkin, E.M.; Sancar, A. Escherichia coli mfd mutant deficient in “mutation frequency decline” lacks strand-specific repair: In vitro complementation with purified coupling factor. Proc. Natl. Acad. Sci. USA 1991, 88, 11574–11578. [Google Scholar] [CrossRef] [PubMed]
- Savery, N.J. The molecular mechanism of transcription-coupled DNA repair. Trends Microbiol. 2007, 15, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Saxowsky, T.T.; Doetsch, P.W. RNA polymerase encounters with DNA damage: Transcription-coupled repair or transcriptional mutagenesis? Chem. Rev. 2006, 106, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Willing, S.E.; Richards, E.J.; Sempere, L.; Dale, A.G.; Cutting, S.M.; Fairweather, N.F. Increased toxin expression in a Clostridium difficile mfd mutant. BMC Microbiol. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Robleto, E.A.; Martin, H.A.; Pedraza-Reyes, M. Mfd and transcriptional derepression cause genetic diversity in Bacillus subtilis. Front. Biosci. 2012, 4, 1246–1254. [Google Scholar] [CrossRef]
- Belitsky, B.R.; Sonenshein, A.L. Roadblock repression of transcription by Bacillus subtilis CodY. J. Mol. Biol. 2011, 411, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Zalieckas, J.M.; Wray, L.V., Jr.; Ferson, A.E.; Fisher, S.H. Transcription-repair coupling factor is involved in carbon catabolite repression of the Bacillus subtilis hut and gnt operons. Mol. Microbiol. 1998, 27, 1031–1038. [Google Scholar] [CrossRef] [PubMed]



© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Verstraete, I.; Peltier, J.; Dupuy, B. The Regulatory Networks That Control Clostridium difficile Toxin Synthesis. Toxins 2016, 8, 153. https://doi.org/10.3390/toxins8050153
Martin-Verstraete I, Peltier J, Dupuy B. The Regulatory Networks That Control Clostridium difficile Toxin Synthesis. Toxins. 2016; 8(5):153. https://doi.org/10.3390/toxins8050153
Chicago/Turabian StyleMartin-Verstraete, Isabelle, Johann Peltier, and Bruno Dupuy. 2016. "The Regulatory Networks That Control Clostridium difficile Toxin Synthesis" Toxins 8, no. 5: 153. https://doi.org/10.3390/toxins8050153
APA StyleMartin-Verstraete, I., Peltier, J., & Dupuy, B. (2016). The Regulatory Networks That Control Clostridium difficile Toxin Synthesis. Toxins, 8(5), 153. https://doi.org/10.3390/toxins8050153