
Neutralization of Botulinum Neurotoxin Type E by a Humanized Antibody

 , , and
, , and
Abstract
:
1. Introduction
2. Results
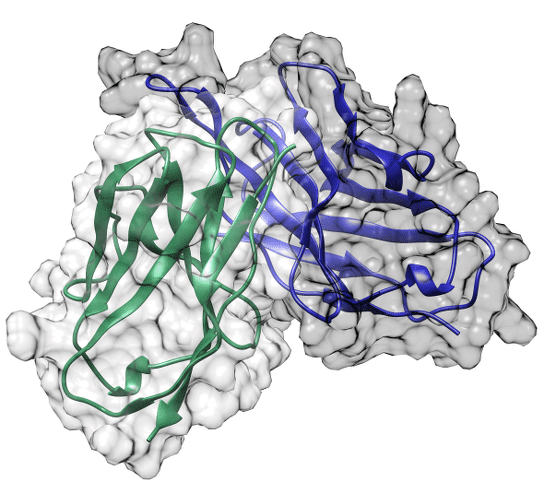
2.1. Comparison between scFv ELC18 and Human Germline Genes
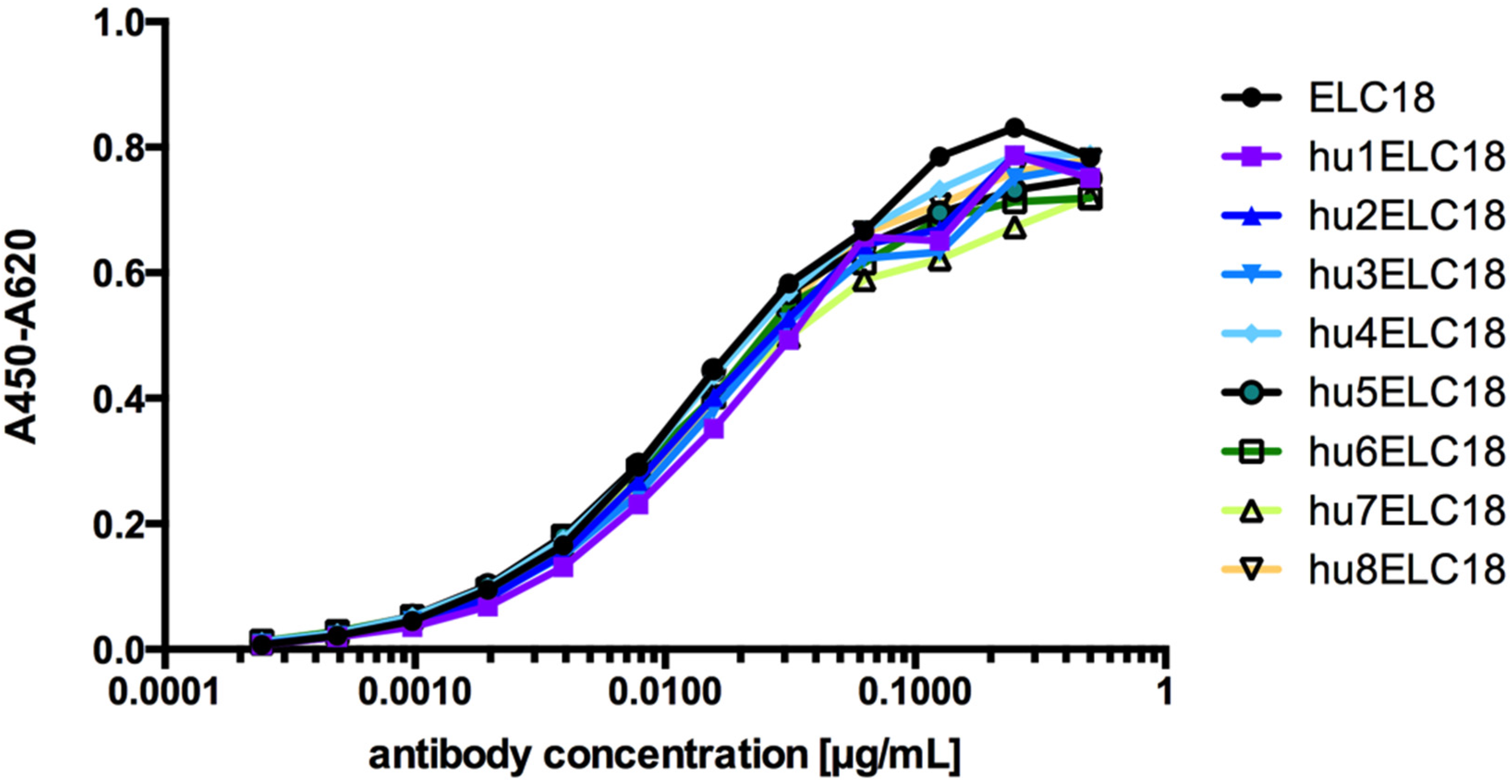
2.2. Humanization of the Macaque scFv ELC18 by Germline-Humanization
2.3. Mouse Protection Assay Performed with IgG hu8ELC18
2.4. Mouse Paralysis Assay Performed with IgG hu8ELC18
2.5. Prophylaxis Assays Performed with IgG hu8ELC18
3. Discussion
4. Materials and Methods
4.1. Ethical Statement and Animal Care
4.2. Humanization of scFv ELC18 by Germline-Humanization
4.3. Production and Purification of scFv-Fc Antibodies
4.4. ELISA Assay
4.5. Cloning of Germline-Humanized IgG hu8ELC18
4.6. Production and Purification of IgG hu8ELC18
4.7. BoNT Toxin Complex Preparation
4.8. Toxicity Titration and Calibration of BoNT Complexes
4.9. Mouse Protection Assays
4.10. Mouse Paralysis Assay
4.11. Prophylaxis Assay
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Arnon, S.S.; Schechter, R.; Inglesby, T.V.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Fine, A.D.; Hauer, J.; Layton, M. Botulinum toxin as a biological weapon: Medical and public health management. JAMA 2001, 285, 1059–1070. [Google Scholar] [CrossRef] [PubMed]
- Sesardic, D.; Jones, R.G.; Leung, T.; Alsop, T.; Tierney, R. Detection of antibodies against botulinum toxins. Mov. Disord. 2004, 19, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Hatheway, C.L. Clostridium botulinum and other clostridia that produce botulinum neurotoxin. In Clostridium botulinum, Ecology and Control in Foods; Hauschild, A.H.W., Doods, K.L., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1993; Volume 13, pp. 3–20. [Google Scholar]
- Lindström, M.; Korkeala, H. Laboratory diagnostics of botulism. Clin. Microbiol. Rev. 2006, 19, 298–314. [Google Scholar] [CrossRef] [PubMed]
- Peck, M.W. Biology and genomic analysis of Clostridium botulinum. Adv. Microb. Physiol. 2009, 55, 183–320. [Google Scholar] [PubMed]
- Hill, K.K.; Smith, T.J. Genetic diversity within Clostridium botulinum serotypes, botulinum neurotoxin gene clusters and toxin subtypes. In Botulinum Neurotoxins; Rummel, A., Binz, T., Eds.; Springer Berlin Heidelberg: Heidelberg, Germany, 2013; Volume 364, pp. 1–20. [Google Scholar]
- Barash, J.R.; Arnon, S.S. A novel strain of Clostridium botulinum that produces type B and type H botulinum toxins. J. Infect. Dis. 2014, 209, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Dover, N.; Barash, J.R.; Hill, K.K.; Xie, G.; Arnon, S.S. Molecular characterization of a novel botulinum neurotoxin type H gene. J. Infect. Dis. 2014, 209, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Maslanka, S.E.; Luquez, C.; Dykes, J.K.; Tepp, W.H.; Pier, C.L.; Pellett, S.; Raphael, B.H.; Kalb, S.R.; Barr, J.R.; Rao, A.; et al. A novel botulinum neurotoxin, previously reported as serotype H, has a hybrid-like structure with regions of similarity to the structures of serotypes A and F and is neutralized with serotype A antitoxin. J. Infect. Dis. 2015, 213, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Korkeala, H.; Aarnikunnas, J.; Lindström, M. Sequencing the botulinum neurotoxin gene and related genes in Clostridium botulinum type E strains reveals orfx3 and a novel type E neurotoxin subtype. J. Bacteriol. 2007, 189, 8643–8650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couesnon, A.; Raffestin, S.; Popoff, M.R. Expression of botulinum neurotoxins A and E, and associated non-toxin genes, during the transition phase and stability at high temperature: Analysis by quantitative reverse transcription-PCR. Microbiology 2006, 152, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Froude, J.W.; Stiles, B.G.; Pelat, T.; Thullier, P. Antibodies for biodefense. mAbs 2011, 3, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Rosenau, W. Aum shinrikyo’s biological weapons program: Why did it fail? Stud. Confl. Terror. 2001, 24, 289–301. [Google Scholar] [CrossRef]
- Sakaguchi, G. Clostridium botulinum toxins. Pharmacol. Ther. 1982, 19, 165–194. [Google Scholar] [CrossRef] [PubMed]
- Popoff, M.R.; Bouvet, P. Genetic characteristics of toxigenic clostridia and toxin gene evolution. Toxicon 2013, 75, 63–89. [Google Scholar] [CrossRef] [PubMed]
- Yokosawa, N.; Tsuzuki, K.; Syuto, B.; Oguma, K. Activation of Clostridium botulinum type E toxin purified by two different procedures. J. Gen. Microbiol. 1986, 132, 1981–1988. [Google Scholar] [CrossRef] [PubMed]
- Pellizzari, R.; Rossetto, O.; Schiavo, G.; Montecucco, C. Tetanus and botulinum neurotoxins: Mechanism of action and therapeutic uses. Philos Trans. R. Soc. Lond. B Biol. Sci. 1999, 354, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Montecucco, C. How do tetanus and botulinum toxins bind to neuronal membranes? Trends Biochem. Sci. 1986, 11, 314–317. [Google Scholar] [CrossRef]
- Humeau, Y.; Doussau, F.; Grant, N.J.; Poulain, B. How botulinum and tetanus neurotoxins block neurotransmitter release. Biochimie 2000, 82, 427–446. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L.L. Identification of the major steps in botulinum toxin action. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 167–193. [Google Scholar] [CrossRef] [PubMed]
- Schiavo, G.; Matteoli, M.; Montecucco, C. Neurotoxins affecting neuroexocytosis. Physiol. Rev. 2000, 80, 717–766. [Google Scholar] [PubMed]
- Elias, M.; Al-Saleem, F.; Ancharski, D.M.; Singh, A.; Nasser, Z.; Olson, R.M.; Simpson, L.L. Evidence that botulinum toxin receptors on epithelial cells and neuronal cells are not identical: Implications for development of a non-neurotropic vaccine. J. Pharmacol. Exp. Ther. 2011, 336, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Pirazzini, M.; Tehran, D.A.; Zanetti, G.; Lista, F.; Binz, T.; Shone, C.C.; Rossetto, O.; Montecucco, C. The thioredoxin reductase-Thioredoxin redox system cleaves the interchain disulphide bond of botulinum neurotoxins on the cytosolic surface of synaptic vesicles. Toxicon 2015, 107, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; Ochiai, M.; Liu, Y.; Ekong, T.; Sesardic, D. Development of improved SNAP25 endopeptidase immuno-assays for botulinum type A and E toxins. J. Immunol. Methods 2008, 329, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Leclair, D.; Farber, J.M.; Doidge, B.; Blanchfield, B.; Suppa, S.; Pagotto, F.; Austin, J.W. Distribution of Clostridium botulinum type E strains in Nunavik, Northern Quebec, Canada. Appl. Environ. Microbiol. 2013, 79, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Hielm, S.; Hyytiä, E.; Andersin, A.; Korkeala, H. A high prevalence of Clostridium botulinum type E in Finnish freshwater and Baltic Sea sediment samples. J. Appl. Microbiol. 1998, 84, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Hyytiä, E.; Hielm, S.; Björkroth, J.; Korkeala, H. Biodiversity of Clostridium botulinum type E strains isolated from fish and fishery products. Appl. Environ. Microbiol. 1999, 65, 2057–2064. [Google Scholar] [PubMed]
- Hielm, S.; Björkroth, J.; Hyytiä, E.; Korkeala, H. Prevalence of Clostridium botulinum in Finnish trout farms: Pulsed-field gel electrophoresis typing reveals extensive genetic diversity among type E isolates. Appl. Environ. Microbiol. 1998, 64, 4161–4167. [Google Scholar] [PubMed]
- King, L.A.; Niskanen, T.; Junnikkala, M.; Moilanen, E.; Lindström, M.; Korkeala, H.; Korhonen, T.; Popoff, M.; Mazuet, C.; Callon, H.; et al. Botulism and hot-smoked whitefish: A family cluster of type E botulism in France, September 2009. Euro Surveill. 2009, 14, 429–433. [Google Scholar]
- Lindström, M.; Vuorela, M.; Hinderink, K.; Korkeala, H.; Dahlsten, E.; Raahenmaa, M.; Kuusi, M. Botulism associated with vacuum-packed smoked whitefish in Finland, June–July 2006. Euro Surveill 2006. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=3004 (accessed on 23 August 2016). [Google Scholar]
- Mazuet, C.; Sautereau, J.; Legeay, C.; Bouchier, C.; Bouvet, P.; Popoff, M.R. An atypical outbreak of food-borne botulism due to clostridium botulinum types B and E from ham. J. Clin. Microbiol. 2015, 53, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L.L.; DasGupta, B.R. Botulinum neurotoxin type E: Studies on mechanism of action and on structure-activity relationships. J. Pharmacol. Exp. Ther. 1983, 224, 135–140. [Google Scholar] [PubMed]
- Arnon, S.S. Human Tetanus and Human Botulism. In Clostridia: Molecular Biology and Pathogenesis; Rood, J., Mc Clane, B., Songer, J., Titball, R., Eds.; Academic Press: San Diego, CA, USA, 1997; pp. 95–111. [Google Scholar]
- Cherington, M. Clinical spectrum of botulism. Muscle Nerve 1998, 21, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Meunier, F.A.; Schiavo, G.; Molgó, J. Botulinum neurotoxins: From paralysis to recovery of functional neuromuscular transmission. J. Physiol. Paris 2002, 96, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Arnon, S.S.; Schechter, R.; Maslanka, S.E.; Jewell, N.P.; Hatheway, C.L. Human botulism immune globulin for the treatment of infant botulism. N. Engl. J. Med. 2006, 354, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Investigational heptavalent botulinum antitoxin (HBAT) to replace licensed botulinum antitoxin AB and investigational botulinum antitoxin E. MMWR Morb. Mortal. Wkly. Rep. 2010, 59, 299. [Google Scholar]
- Black, R.E.; Gunn, R.A. Hypersensitivity reactions associated with botulinal antitoxin. Am. J. Med. 1980, 69, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Hibbs, R.G.; Weber, J.T.; Corwin, A.; Allos, B.M.; Abd el Rehim, M.S.; Sharkawy, S.E.; Sarn, J.E.; McKee, K.T., Jr. Experience with the use of an investigational F(ab’)2 heptavalent botulism immune globulin of equine origin during an outbreak of type E botulism in Egypt. Clin. Infect. Dis. 1996, 23, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Pelat, T.; Thullier, P. Non-human primate immune libraries combined with germline humanization: An (almost) new and powerful approach for the isolation of therapeutic antibodies. mAbs 2009, 1, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.; Mitchell, D.A.; Buss, T.N.; Holmes, M.A.; Anasetti, C.; Foote, J. “Superhumanized” antibodies: Reduction of immunogenic potential by complementarity-determining region grafting with human germline sequences: Application to an anti-CD28. J. Immunol. 2002, 169, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.T.; Jolly, C.J.; Köhler, J.; Neuberger, M.S. The contribution of somatic hypermutation to the diversity of serum immunoglobulin: Dramatic increase with age. Immunity 2000, 13, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Pelat, T.; Bedouelle, H.; Rees, A.R.; Crennell, S.J.; Lefranc, M.; Thullier, P. Germline humanization of a non-human primate antibody that neutralizes the anthrax toxin, by in vitro and in silico engineering. J. Mol. Biol. 2008, 384, 1400–1407. [Google Scholar] [CrossRef] [PubMed]
- Robert, R.; Streltsov, V.A.; Newman, J.; Pearce, L.A.; Wark, K.L.; Dolezal, O. Germline humanization of a murine Aβ antibody and crystal structure of the humanized recombinant Fab fragment. Protein Sci. 2010, 19, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Miethe, S.; Rasetti-Escargueil, C.; Avril, A.; Liu, Y.; Chahboun, S.; Korkeala, H.; Mazuet, C.; Popoff, M.R.; Pelat, T.; Thullier, P.; et al. Development of human-like scFv-fc neutralizing botulinum neurotoxin E. PLoS ONE 2015, 10, e0139905. [Google Scholar] [CrossRef] [PubMed]
- Rosenwasser, L.J.; Busse, W.W.; Lizambri, R.G.; Olejnik, T.A.; Totoritis, M.C. Allergic asthma and an anti-CD23 mAb (IDEC-152): Results of a phase I, single-dose, dose-escalating clinical trial. J. Allergy Clin. Immunol. 2003, 112, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Miethe, S.; Mazuet, C.; Liu, Y.; Tierney, R.; Rasetti-Escargueil, C.; Avril, A.; Frenzel, A.; Thullier, P.; Pelat, T.; Urbain, R.; et al. Development of germline-humanized antibodies neutralizing botulinum neurotoxin A and B. PLoS ONE 2016, in press. [Google Scholar] [CrossRef] [PubMed]
- Byrd, J.C.; O’Brien, S.; Flinn, I.W.; Kipps, T.J.; Weiss, M.; Rai, K.; Lin, T.S.; Woodworth, J.; Wynne, D.; Reid, J.; et al. Phase 1 study of lumiliximab with detailed pharmacokinetic and pharmacodynamic measurements in patients with relapsed or refractory chronic lymphocytic leukemia. Clin. Cancer Res. 2007, 13, 4448–4455. [Google Scholar] [CrossRef] [PubMed]
- Hust, M.; Meyer, T.; Voedisch, B.; Rülker, T.; Thie, H.; El-Ghezal, A.; Kirsch, M.I.; Schütte, M.; Helmsing, S.; Meier, D.; et al. A human scFv antibody generation pipeline for proteome research. J. Biotechnol. 2011, 152, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Thyagarajan, B. Antidotes to botulinum neurotoxin. In Biological Toxins and Bioterrorism; Gopalakrishnakone, P., Balali-Mood, M., Ram Singh, B., Llewellyn, L., Eds.; Springer: Heidelberg, Germany, 2015; pp. 376–397. [Google Scholar]
- Smith, L.A. Botulism and vaccines for its prevention. Vaccine 2009, 27, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Rusnak, J.M.; Smith, L.A. Botulinum neurotoxin vaccines: Past history and recent developments. Hum. Vaccines 2009, 5, 794–805. [Google Scholar] [CrossRef]
- Foster, K.A. Molecular Aspects of Botulinum Neurotoxin; Springer: New York, NY, USA, 2014. [Google Scholar]
- Fagan, R.P.; Neil, K.P.; Sasich, R.; Luquez, C.; Asaad, H.; Maslanka, S.; Khalil, W. Initial recovery and rebound of type F intestinal colonization botulism after administration of investigational heptavalent botulinum antitoxin. Clin. Infect. Dis. 2011, 53, e125–e128. [Google Scholar] [CrossRef] [PubMed]
- IMGT/V-QUEST Online Tool. Available online: http://www.imgt.org (accessed on 26 August 2016).
- Jäger, V.; Büssow, K.; Wagner, A.; Weber, S.; Hust, M.; Frenzel, A.; Schirrmann, T. High level transient production of recombinant antibodies and antibody fusion proteins in HEK293 cells. BMC Biotechnol. 2013, 13, 52–71. [Google Scholar] [CrossRef] [PubMed]
- Sibéril, S.; Dutertre, C.; Boix, C.; Bonnin, E.; Ménez, R.; Stura, E.; Jorieuxb, S.; Fridmana, W.; Teillaud, J. Molecular aspects of human FcγR interactions with IgG: Functional and therapeutic consequences. Immunol. Lett. 2006, 106, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, K.; Steindler, K.A.; Harrison, S.J. Limulus amoebocyte lysate assay for detection and quantitation of endotoxin in a small-volume parenteral product. Appl. Environ. Microbiol. 1980, 40, 533–538. [Google Scholar] [PubMed]
- Guttman, A. On the separation mechanism of capillary sodium dodecyl sulfate-gel electrophoresis of proteins. Electrophoresis 1995, 16, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Szabo, Z.; Guttman, A.; Bones, J.; Karger, B.L. Rapid high-resolution characterization of functionally important monoclonal antibody N-glycans by capillary electrophoresis. Anal. Chem. 2011, 83, 5329–5336. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.G.; Alsop, T.A.; Hull, R.; Tierney, R.; Holley, J.; Sesardic, D. Botulinum type A toxin neutralisation by specific IgG and its fragments: A comparison of mouse systemic toxicity and local flaccid paralysis assays. Toxicon 2006, 48, 246–254. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Framework Region | Total Number of AAs | Number of Identical AAs | Number of Very Similar AAs | Number of Similar AAs | Number of Dissimilar AAs | Number of Very Dissimilar AAs | |
|---|---|---|---|---|---|---|---|
| VH | |||||||
| FR1-IMGT | 25 | 20 | 0 | 2 | 2 | 1 | |
| FR2-IMGT | 17 | 16 | 0 | 0 | 1 | 0 | |
| FR3-IMGT | 38 | 33 | 0 | 2 | 2 | 1 | |
| FR4-IMGT | 11 | 9 | 0 | 0 | 1 | 1 | |
| FR-IMGT | 91 | 78 | 0 | 4 | 6 | 3 | |
| GI | 87.5% | 100% | 0% | 4.4% | 6.6% | 3.3% | |
| VL | |||||||
| FR1-IMGT | 26 | 23 | 1 | 1 | 0 | 1 | |
| FR2-IMGT | 17 | 15 | 1 | 0 | 1 | 0 | |
| FR3-IMGT | 36 | 33 | 0 | 0 | 2 | 1 | |
| FR4-IMGT | 10 | 9 | 0 | 1 | 0 | 0 | |
| FR-IMGT | 89 | 80 | 2 | 2 | 3 | 2 | |
| GI | 89.9% | 100% | 2.2% | 2.2% | 3.4% | 2.2% | |
| VH and VL | 180 | 158 | 2 | 6 | 9 | 5 | |
| GI | 87.8% | 100% | 1.1% | 3.3% | 5.0% | 2.8% | |
| Variant | VH Variant | VL Variant | Total GI |
|---|---|---|---|
| ELC18 | Macaque VH | Macaque VL | 87.8% |
| hu1ELC18 | Macaque VH | hu1VL | 90.1% |
| hu2ELC18 | Macaque VH | hu2VL | 91.8% |
| hu3ELC18 | hu1VH | Macaque VL | 90.0% |
| hu4ELC18 | hu1VH | hu1VL | 92.3% |
| hu5ELC18 | hu1VH | hu2VL | 94.0% |
| hu6ELC18 | hu2VH | Macaque VL | 93.3% |
| hu7ELC18 | hu2VH | hu1VL | 95.6% |
| hu8ELC18 | hu2VH | hu1VL | 97.3% |
| hu8ELC18 IgG Concentration (µg) | Number of Survived/Total Mice |
|---|---|
| 25 | 4/4 |
| 2.5 | 4/4 |
| 0.25 | 4/4 |
| 0.025 | 2/2 |
| 0.0025 | 2/2 |
| 0 | 0/4 |
| BoNT/E Injection after hu8ELC18 Administration (Days) | Number of Survived Mice/Total Mice |
|---|---|
| 1 | 3/3 |
| 3 | 4/4 |
| 7 | 4/4 |
| 14 | 4/4 |
| Control (no antibody) | 0/4 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derman, Y.; Selby, K.; Miethe, S.; Frenzel, A.; Liu, Y.; Rasetti-Escargueil, C.; Avril, A.; Pelat, T.; Urbain, R.; Fontayne, A.; et al. Neutralization of Botulinum Neurotoxin Type E by a Humanized Antibody. Toxins 2016, 8, 257. https://doi.org/10.3390/toxins8090257
Derman Y, Selby K, Miethe S, Frenzel A, Liu Y, Rasetti-Escargueil C, Avril A, Pelat T, Urbain R, Fontayne A, et al. Neutralization of Botulinum Neurotoxin Type E by a Humanized Antibody. Toxins. 2016; 8(9):257. https://doi.org/10.3390/toxins8090257
Chicago/Turabian StyleDerman, Yağmur, Katja Selby, Sebastian Miethe, André Frenzel, Yvonne Liu, Christine Rasetti-Escargueil, Arnaud Avril, Thibaut Pelat, Remi Urbain, Alexandre Fontayne, and et al. 2016. "Neutralization of Botulinum Neurotoxin Type E by a Humanized Antibody" Toxins 8, no. 9: 257. https://doi.org/10.3390/toxins8090257
APA StyleDerman, Y., Selby, K., Miethe, S., Frenzel, A., Liu, Y., Rasetti-Escargueil, C., Avril, A., Pelat, T., Urbain, R., Fontayne, A., Thullier, P., Sesardic, D., Lindström, M., Hust, M., & Korkeala, H. (2016). Neutralization of Botulinum Neurotoxin Type E by a Humanized Antibody. Toxins, 8(9), 257. https://doi.org/10.3390/toxins8090257