
Variations of Growth and Toxin Yield in Cylindrospermopsis raciborskii under Different Phosphorus Concentrations

 ,
, 
Abstract
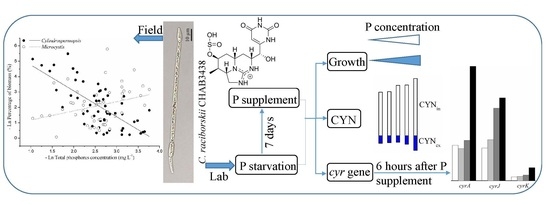
:
1. Introduction
2. Results
2.1. Effects of P Concentration on the Growth and Morphology of C. raciborskii
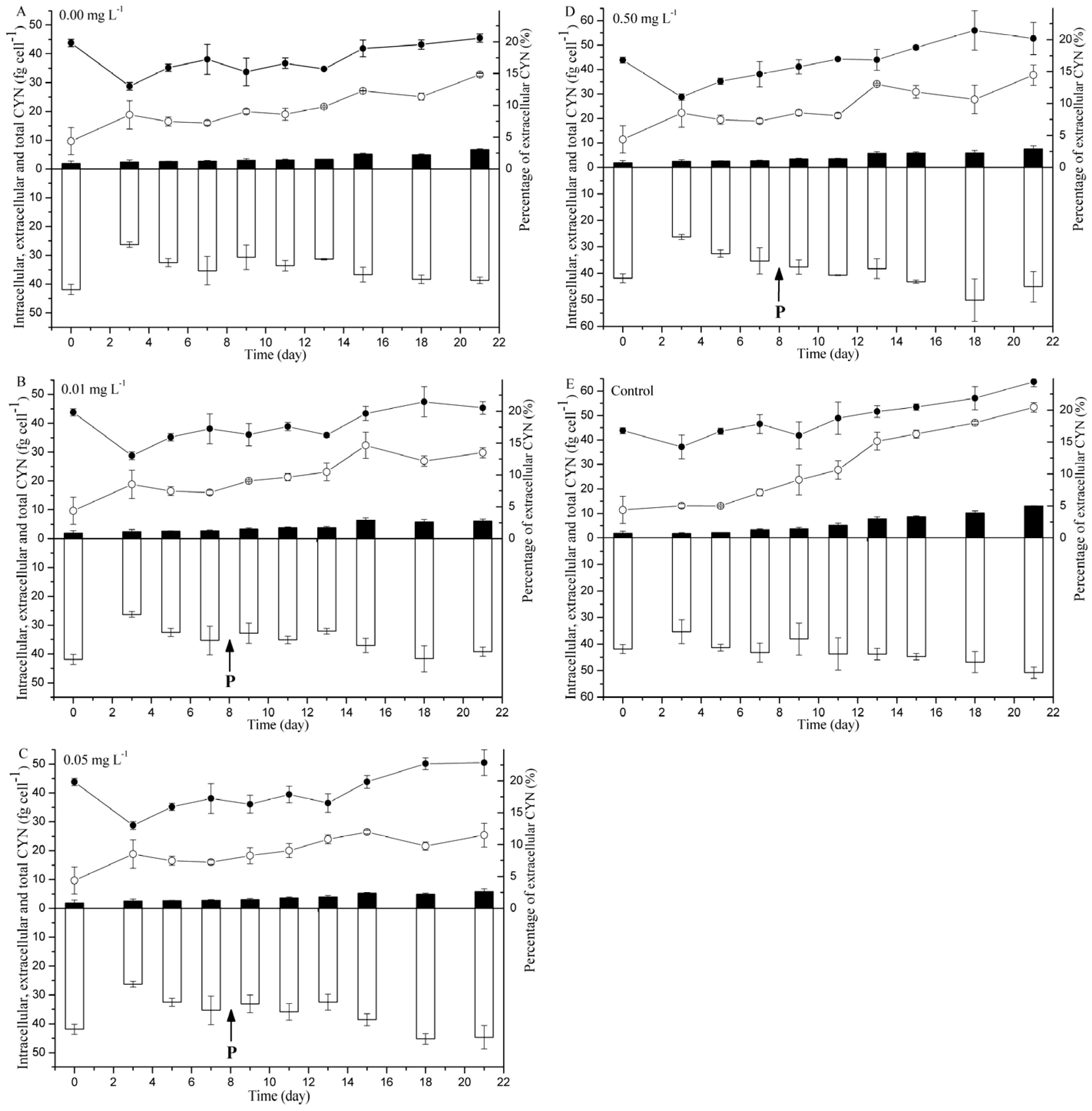
2.2. CYN Measurement
2.3. cyr Gene Expression
2.4. Correlation between the Biomass of Cylindrospermopsis and TP Concentration in Freshwater Bodies
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strain and Culture Conditions
5.2. Phosphorus Starvation and Supplementation
5.3. Measurement of Growth
5.4. Extraction and Quantitation of CYN
5.5. RNA Isolation and cDNA Synthesis
5.6. Quantitative PCR (qPCR) Assay
5.7. Collection of Water Samples
5.8. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sinha, R.; Pearson, L.A.; Davis, T.W.; Burford, M.A.; Orr, P.T.; Neilan, B.A. Increased incidence of Cylindrospermopsis raciborskii in temperate zones—Is climate change responsible? Water Res. 2012, 46, 1408–1419. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G. Harmful cyanobacterial blooms: Causes, consequences, and controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Zurawell, R.W.; Chen, H.; Burke, J.M.; Prepas, E.E. Hepatotoxic cyanobacteria: A review of the biological importance of microcystins in freshwater environments. J. Toxicol. Environ. Health B 2005, 8, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, M.; Krishnamurthy, T.; Beasley, V.; Yu, M.J.; Bunner, D.L.; Moore, R.E.; Eloff, J.N.; Rinehart, K.; Falconer, I.; Runnegar, M.; et al. Naming of cyclic heptapeptide toxins of cyanobacteria (blue–green algae). Toxicon 1988, 26, 971–973. [Google Scholar] [CrossRef]
- Ohtani, I.; Moore, R.E.; Runnegar, M.T.C. Cylindrospermopsin: A potent hepatotoxin from the blue–green alga Cylindrospermopsis raciborskii. J. Am. Chem. Soc. 1992, 114, 7942–7944. [Google Scholar] [CrossRef]
- Schantz, E.J.; Ghazarossian, V.E.; Schnoes, H.K.; Strong, F.M.; Springer, J.P.; Pezzanite, J.O.; Clardy, J. Structure of saxitoxin. J. Am. Chem. Soc. 1975, 97, 1238–1239. [Google Scholar] [CrossRef] [PubMed]
- Devlin, J.P.; Edwards, O.E.; Gorham, P.R.; Hunter, M.R.; Pike, R.K.; Stavric, B. Anatoxin–a, a toxic alkaloid from Anabaena flos–aquae NCR–44h. Can. J. Chem. 1977, 55, 1367–1371. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce β–N–methylamino–l–alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef] [PubMed]
- Cardellina, J.H.; Marner, F.J.; Moore, R.E. Seaweed dermatitis: Structure of lyngbyatoxin A. Science 1979, 204, 193–195. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Scheuer, P.J. Aplysiatoxin and debromoaplysiatoxin, constituents of the marine mollusk Stylocheilus longicauda (Quoy and Gaimard, 1824). J. Am. Chem. Soc. 1974, 96, 2245–2246. [Google Scholar] [CrossRef] [PubMed]
- Bourke, A.T.C.; Hawes, R.B.; Neilson, A.; Stallman, N.D. An outbreak of hepato–enteritis (the Palm Island mystery disease) possibly caused by algal intoxication. Toxicon 1983, 21, 45–48. [Google Scholar] [CrossRef]
- Hawkins, P.R.; Runnegar, M.T.C.; Jackson, A.R.B.; Falconer, I. Severe hepatotoxicity caused by the tropical cyanobactreium (blue–green alga) Cylindrospermopsis raciborskii (Woloszaynska) Seenaya and Subba Raju isolated from a domestic supply reservoir. Appl. Environ. Microbiol. 1985, 50, 1292–1295. [Google Scholar] [PubMed]
- Wormer, L.; Cirés, A.; Carrasco, D.; Quesada, A. Cylindrospermopsin is not degraded by co–occurring natural bacterial communities during a 40–day study. Harmful Algae 2008, 7, 206–213. [Google Scholar] [CrossRef]
- Runnegar, M.T.; Kong, S.M.; Zhong, Y.Z.; Lu, S.C. Inhibition of reduced glutathione synthesis by cyanobacterial alkaloid cylindrospermopsin in cultured rat hepatocytes. Biochem. Pharmacol. 1995, 49, 219–225. [Google Scholar] [CrossRef]
- Froscio, S.M.; Humpage, A.R.; Burcham, P.C.; Falconer, I.R. Cylindrospermopsin–induced protein synthesis inhibition and its dissociation from acute toxicity in mouse hepatocytes. Environ. Toxicol. 2003, 18, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Terao, K.; Ohmori, S.; Igarashi, K.; Ohtani, I.; Watanabe, M.F.; Harada, K.I.; Ito, E.; Watanabe, M. Electron microscopic studies on experimental poisoning in mice induced by cylindrospermopsin isolated from blue–green alga Umezakia natans. Toxicon 1994, 32, 833–843. [Google Scholar] [CrossRef]
- Banker, R.; Carmeli, S.; Werman, M.; Teltsch, B.; Porat, R.; Sukenik, A. Uracil moiety is required for toxicity of the cyanobacterial hepatotoxin cylindrospermopsin. J. Toxicol. Environ. Health A 2001, 62, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Carmichael, W.W.; Brittain, S.; Eagalesham, G.K.; Shaw, G.R.; Mahakhant, A.; Noparatnarorn, N.; Yongmanitchai, W.; Kaya, K.; Watamabe, M.M. Isolation and identification of the cyanotoxin cylindrospermopsin and deoxy–cylindrosermopsin from a Thailand strain of Cylindrospermopsis raciborskii (Cyanobacteia). Toxicon 2001, 39, 937–980. [Google Scholar] [CrossRef]
- Seifert, M.; McGregor, G.; Eaglesham, G.; Wickramasinghe, W.; Shaw, G. First evidence for the production of cylindrospermopsin and deoxy–cylindrospermopsin by the freshwater benthic cyanobacterium Lyngbya wollei (Farlow ex Gomont) Speziale and Dyck. Harmful Algae 2007, 6, 73–80. [Google Scholar] [CrossRef]
- Mihali, T.K.; Kellmann, R.; Muenchhoff, J.; Barrow, K.D.; Neilan, B.A. Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis. Appl. Environ. Microbiol. 2008, 74, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Nixdorf, B.; Mischke, U.; Rücker, J. Phytoplankton assemblages and steady state in deep and shallow eutrophic lakes—An approach to differentiate the habitat properties of Oscillatoriales. Hydrobiology 2003, 502, 111–121. [Google Scholar] [CrossRef]
- Paerl, H.W.; Hall, N.S.; Calandrino, E.S. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic–induced change. Sci. Total Environ. 2011, 409, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- Figueredo, C.C.; Giani, A. Phytoplankton community in the tropical lake of Lagoa Santa (Brazil): Conditions favoring a persistent bloom of Cylindrospermopsis raciborskii. Limnologica 2009, 39, 264–272. [Google Scholar] [CrossRef]
- Vidal, L.; Kruk, C. Cylindrospermopsis raciborskii (Cyanobacteria) extends its distribution to Latitude 34°53′ S: Taxonomical and ecological features in Uruguayan eutrophic lakes. Pan–Am. J. Aquat. Sci. 2008, 3, 142–151. [Google Scholar]
- Everson, S.; Fabbro, L.; Kinner, S.; Wright, P. Extreme differences in akinete, heterocyte and cylindrospermopsin concentrations with depth in a successive bloom involving Aphanizomenon ovalisporum (Forti) and Cylindrospermopsis raciborskii (Woloszynska) Seenaya and Subba Raju. Harmful Algae 2011, 10, 265–276. [Google Scholar] [CrossRef]
- Stüken, A.; Rucker, J.; Endrulat, T.; Preussel, K.; Hemm, M.; Nixdorf, B.; Karsten, U.; Wiedner, C. Distribution of three alien cyanobacterial species (Nostocales) in northeast Germany: Cylindrospermopsis raciborskii, Anabaena bergii and Aphanizomenon aphanizomenoides. Phycologia 2006, 45, 696–703. [Google Scholar] [CrossRef]
- Rzymski, P.; Poniedziałek, B. In search of environmental role of cylindrospermopsin: A review on global distribution and ecology of its producers. Water Res. 2014, 66, 320–337. [Google Scholar] [CrossRef] [PubMed]
- Burford, M.A.; Beardall, J.; Willis, A.; Orr, P.T.; Magalhaes, V.F.; Rangel, L.; Azevedo, S.M.F.O.E.; Neilan, B.A. Understanding the winning strategies used by the toxic cyanobacterium Cylindrospermopsis raciborskii. Harmful Algae 2016, 54, 44–53. [Google Scholar] [CrossRef]
- Briand, J.F.; Leboulange, C.; Humbert, J.F.; Bernard, C.; Dufour, P. Cylidrospermopsis raciborskii (Cyanobacteria) invasion at mid–latitudes: Selection, wide physiological tolerance, or global warming? J. Phycol. 2004, 40, 231–238. [Google Scholar] [CrossRef]
- Padisák, J. Cylindrospermopsis raciborskii (Woloszynska) Seenaya et Subba Raju, an expanding, highly adaptive cyanobacterium: Worldwide distribution and review of its ecology. Arch. Hydrobiol. 1997, 107, 563–593. [Google Scholar]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Smith, V.H. Low nitrogen to phosphorus ratios favor dominance by blue–green algae in lake phytoplankton. Science 1983, 221, 669–671. [Google Scholar] [CrossRef] [PubMed]
- Valiei, A.I. Producers and processes involved in primary production. In Marine Ecological Processes, 2nd ed.; Springer: New York, NY, USA, 1995; pp. 3–35. [Google Scholar]
- Trimbee, A.M.; Prepas, E.E. Evaluation of total phosphorus as a predictor of the relative biomass of blue-green-algae with emphasis on Alberta lakes. Can. J. Fish. Aquat. Sci. 1987, 44, 1337–1342. [Google Scholar] [CrossRef]
- Heisler, J.; Glibert, P.M.; Burkholder, J.M.; Anderson, D.M.; Cochlan, W.; Dennison, W.C.; Dortch, Q.; Gobler, C.J.; Heil, C.A.; Lewitus, A.; et al. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, Y.; Lin, D.; Shan, K.; Luo, Y.; Zhao, L.; Tan, Z.; Song, L. The relationships of meteorological factors and nutrient levels with phytoplankton biomass in a shallow eutrophic lake dominated by cyanobacteria, Lake Dianchi from 1991 to 2013. Environ. Sci. Pollut. Res. 2016, 23, 15616–15626. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tang, L.; Chen, L.; Li, J. Spatial and temporal variations of phosphorus in Dianchi Lake. J. Agro-Environ. Sci. 2005, 24, 1145–1151. (In Chinese) [Google Scholar]
- Wang, Z.; Shi, S.; Li, S. The constituent distribution of phosphorus and its effects on phytoplankton in Jiulong River estuary in spring. Adm. Tech. Environ. Monit. 2008, 20, 62–65, (In Chinese with English Abstract). [Google Scholar]
- Dyhrman, S.T.; Ammerman, J.W.; Van Mooy, B.A.S. Microbes and the marine phosphorus cycle. Oceanography 2007, 20, 110–116. [Google Scholar] [CrossRef]
- Harke, M.J.; Berry, D.L.; Ammerman, J.W.; Gobler, C.J. Molecular response of the bloom-forming cyanobacterium, Microcystis aeruginosa, to phosphorus limitation. Microb. Ecol. 2012, 63, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Köhler, J.; Hilt, S.; Adrian, R.; Nicklisch, A.; Kozerski, H.P.; Walz, N. Long–term response of a shallow, moderately flushed lake to reduced external phosphorus and nitrogen loading. Freshw. Biol. 2005, 50, 1639–1650. [Google Scholar] [CrossRef]
- Willis, A.; Chuang, A.W.; Burford, M.A. Nitrogen fixation by the diazotroph Cylindrospermopsis raciborskii (Cyanophyceae). J. Phycol. 2016, 52, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Chislock, M.F.; Sharp, K.L.; Wilson, A.E. Cylindrospermopsis raciborskii dominates under very low and high nitrogen-to-phosphorus ratios. Water Res. 2014, 49, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Bar-Yosef, Y.; Sukenik, A.; Hadas, O.; Viner-Mozzini, Y.; Kaplan, A. Enslavement in the water body by toxic Aphanizomenon ovalisporum, inducing alkaline phosphatase in phytoplanktons. Curr. Biol. 2010, 20, 1557–1561. [Google Scholar] [CrossRef] [PubMed]
- Chiswell, R.K.; Shaw, G.R.; Eaglesham, G.; Smith, M.J.; Norris, R.L.; Seawright, A.A.; Moore, M.R. Stability of cylindrospermopsin, the toxin from the cyanobacterium Cylindrospermopsis raciborskii: Effect of pH, temperature, and sunlight on decomposition. Environ. Toxicol. 1999, 14, 155–161. [Google Scholar] [CrossRef]
- Rücker, J.; Stüken, A.; Nixdorf, B.; Fastner, J.; Chorus, I.; Wiedner, C. Concentrations of particulate and dissolved cylindrospermopsin in 21 Aphanizomenon–dominated temperate lakes. Toxicon 2007, 50, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Xiao, P.; Yu, G.; Shao, J.; Liu, D.; Azevedo, S.M.; Li, R. Sporadic distribution and distinctive variations of cylindrospermopsin genes in cyanobacterial strains and environmental samples from Chinese freshwater bodies. Appl. Environ. Microbiol. 2014, 80, 5219–5230. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, P.R.; Putt, E.; Falconer, I.; Humpage, A. Phenotypical variation in a toxic strain of the phytoplankter, Cylindrospermopsis raciborskii (Nostocales Cyanophyceae) during batch culture. Environ. Toxicol. 2001, 16, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Saker, M.L.; Griffiths, D.J. The effect of temperature on growth and cylindrospermopsin content of seven isolates of Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) from water bodies in northern Australia. Phycologia 2000, 39, 349–354. [Google Scholar] [CrossRef]
- Bormans, M.; Lengronne, M.; Brient, L.; Duval, C. Cylindrospermopsin accumulation and release by the benthic cyanobacterium Oscillatoria sp. PCC 6506 under different light conditions and growth phases. Bull. Environ. Contam. Toxicol. 2014, 92, 243–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyble, J.; Tester, P.A.; Litaker, R.W. Effects of light intensity on cylindrospermopsin production in the cyanobacterial HAB species Cylindrospermopsis raciborskii. Afr. J. Mar. Sci. 2006, 28, 309–312. [Google Scholar] [CrossRef]
- Davis, T.W.; Orr, P.T.; Boyer, G.L.; Burford, M.A. Investigating the production and release of cylindrospermopsin and deoxy–cylindrospermopsin by Cylindrosermopsis raciborskii over a natural growth cycle. Harmful Algae 2014, 31, 18–25. [Google Scholar] [CrossRef]
- Istvánovics, V.; Shafik, H.M.; Présing, M.; Juhos, S. Growth and phosphate uptake kinetics of the cyanobacterium, Cylindrospermopsis raciborskii (Cyanophyceae) in throughflow cultures. Freshw. Biol. 2000, 43, 257–275. [Google Scholar] [CrossRef]
- Wu, Z.; Zeng, B.; Li, R.; Song, L. Physiological regulation of Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria) in response to inorganic phosphorus limitation. Harmful Algae 2012, 15, 53–58. [Google Scholar] [CrossRef]
- Amaral, V.; Bonilla, S.; Aubriot, L. Growth optimization of the invasive cyanobacterium Cylindrospermopsis raciborskii in response to phosphate fluctuations. Eur. J. Phycol. 2014, 49, 134–141. [Google Scholar] [CrossRef]
- Steffen, M.M.; Dearth, S.P.; Dill, B.D.; Li, Z.; Larsen, K.M.; Campagna, S.R.; Wilhelm, S.W. Nutrients drive transcriptional changes that maintain metabolic homeostasis but alter genome architecture in Microcystis. ISME J. 2014, 8, 2080–2092. [Google Scholar] [CrossRef] [PubMed]
- Kenesi, G.; Shafik, H.M.; Kovács, A.W.; Herodek, S.; Présing, M. Effect of nitrogen forms on growth, cell composition and N2 fixation of Cylindrospermopsis raciborskii in phosphorus-limited chemostat cultures. Hydrobiologia 2009, 623, 191–202. [Google Scholar] [CrossRef]
- Bonilla, S.; Aubriot, L.; Soares, M.C.S.; González–Piana, M.; Fabre, A.; Huszar, V.L.; Lürling, M.; Antoniades, D.; Padisák, J.; Kruk, C. What drives the distribution of the bloom–forming cyanobacteria Planktothrix agardhii and Cylindrospermopsis raciborskii? FEMS Microbiol. Ecol. 2012, 79, 594–607. [Google Scholar] [CrossRef] [PubMed]
- Bácsi, I.; Vasas, G.; Surányi, G.; Mhamvas, M.; Máthé, C.; Tóth, E.; Grigorszky, I.; Gáspár, A.; Tóth, S.; Borbely, G. Alteration of cylindrospermopsin production in sulfate- or phosphate-starved cyanobacterium Aphanizomenon ovalisporum. FEMS Microbiol. Lett. 2006, 259, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, Z.A.; Al-Shehri, A.M. Assessment of cylindrospermopsin toxin in an arid Saudi lake containing dense cyanobacterial bloom. Environ. Monit. Assess. 2013, 185, 2157–2166. [Google Scholar] [CrossRef] [PubMed]
- Burford, M.A.; Davis, T.W.; Orr, P.T.; Sinha, R.; Willis, A.; Neilan, B.A. Nutrient-related changes in the toxicity of field blooms of the cyanobacterium, Cylindrospermopsis raciborskii. FEMS Microbiol. Ecol. 2014, 89, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Preußel, K.; Chorus, I.; Fastner, J. Nitrogen limitation promotes accumulation and suppresses release of cylindrospermopsins in cells of Aphanizomenon sp. Toxins 2014, 6, 2932–2947. [Google Scholar] [CrossRef] [PubMed]
- Pierangelini, M.; Sinha, R.; Willis, A.; Burford, M.A.; Orr, P.T.; Beardall, J.; Neilan, B.A. Constitutive cylindrospermopsin pool size in Cylindrospermopsis raciborskii under different light and CO2 partial pressure conditions. Appl. Environ. Microbiol. 2015, 81, 3069–3076. [Google Scholar] [CrossRef] [PubMed]
- Willis, A.; Adams, M.P.; Chuang, A.W.; Orr, P.T.; O’Brien, K.R.; Burford, M.A. Constitutive toxin production under various nitrogen and phosphorus regimes of three ecotypes of Cylindrospermopsis raciborskii ((Wołoszyńska) Seenayya et Subba Raju). Harmful Algae 2015, 47, 27–34. [Google Scholar] [CrossRef]
- Orr, P.T.; Rasmussen, J.P.; Burford, M.A.; Eaglesham, G.K.; Lennox, S.M. Evaluation of quantitative real-time PCR to characterise spatial and temporal variations in cyanobacteria, Cylindrospermopsis raciborskii (Woloszynska) Seenaya et Subba Raju and cylindrospermopsin concentrations in three subtropical Australian reservoirs. Harmful Algae 2010, 9, 243–254. [Google Scholar] [CrossRef]
- Stucken, K.; John, U.; Cembella, A.; Soto–Liebe, K.; Vásquez, M. Impact of nitrogen sources on gene expression and toxin production in the diazotroph Cylindrospermopsis raciborskii CS-505 and non-diazotroph Raphidiopsis brookii D9. Toxins 2014, 6, 1896–1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichimura, T. Isolation and culture methods of algae (Sôrui no bunri to baiyôhô. 2.5.B. Tansui sôrui). In Methods in Phycological Studies (Sôrui Kenkyûhô); Nishizawa, K., Chihara, M., Eds.; Kyoritu Shuppan: Tokyo, Japan, 1979; pp. 294–305, (In Japanese without English Title). [Google Scholar]
- Pierangelini, M.; Stojkovic, S.; Orr, P.T.; Beardall, J. Photosynthetic characteristics of two Cylindrospermopsis raciborskii strains differing in their toxicity. J. Phycol. 2014, 50, 292–302. [Google Scholar] [CrossRef] [PubMed]
- China Environmental Protection Administration. Water and Wastewater Monitoring and Analysis Methods, 4th ed.China Environmental Science Press: Beijing, China, 2006; pp. 670–671.
- Wormer, L.; Carrasco, D.; Cirés, S.; Quesada, A. Advances in solid-phase extraction of the cyanobacterial toxin cylindrospermopsin. Limnol. Oceanogr. Methods 2009, 7, 568–575. [Google Scholar] [CrossRef]
- Wu, S.K.; Xie, P.; Liang, G.D.; Wang, S.B.; Liang, X.M. Relationships between microcystins and environmental parameters in 30 subtropical shallow lakes along the Yangtze River, China. Freshw. Biol. 2006, 51, 2309–2319. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′–3′) | Product Size (bp) | Efficiency (%) |
|---|---|---|---|---|
| cyrA | qcyrAF193 | GAGGAGTTGAATGGGCTGGTA | 136 | 98.94 |
| qcyrAR328 | GTGGGCAGACCGCACAATA | |||
| cyrJ | qcyrJF375 | TCTGATTCGCCAACCCAAAG | 133 | 98.63 |
| qcyrJR507 | CGGGATTACTCCGCTCGTT | |||
| cyrK | qcyrKF345 | CGGGAAATAGCCAACACG | 106 | 102.02 |
| qcyrKR450 | AAAGGGAAAGGAGCCACA | |||
| 16S rDNA | q16SF1029 | GTGTCGTGAGATGTTGGGTT | 182 | 101.17 |
| q16SR1210 | CCTCTGTCCGTAGCATTGTAG |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Jiang, Y.; Li, X.; Li, H.; Chen, Y.; Xie, J.; Cai, F.; Li, R. Variations of Growth and Toxin Yield in Cylindrospermopsis raciborskii under Different Phosphorus Concentrations. Toxins 2017, 9, 13. https://doi.org/10.3390/toxins9010013
Yang Y, Jiang Y, Li X, Li H, Chen Y, Xie J, Cai F, Li R. Variations of Growth and Toxin Yield in Cylindrospermopsis raciborskii under Different Phosphorus Concentrations. Toxins. 2017; 9(1):13. https://doi.org/10.3390/toxins9010013
Chicago/Turabian StyleYang, Yiming, Yongguang Jiang, Xiaochuang Li, Hua Li, Youxin Chen, Jinlin Xie, Fangfang Cai, and Renhui Li. 2017. "Variations of Growth and Toxin Yield in Cylindrospermopsis raciborskii under Different Phosphorus Concentrations" Toxins 9, no. 1: 13. https://doi.org/10.3390/toxins9010013
APA StyleYang, Y., Jiang, Y., Li, X., Li, H., Chen, Y., Xie, J., Cai, F., & Li, R. (2017). Variations of Growth and Toxin Yield in Cylindrospermopsis raciborskii under Different Phosphorus Concentrations. Toxins, 9(1), 13. https://doi.org/10.3390/toxins9010013