
Production, Characterisation and Testing of an Ovine Antitoxin against Ricin; Efficacy, Potency and Mechanisms of Action

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
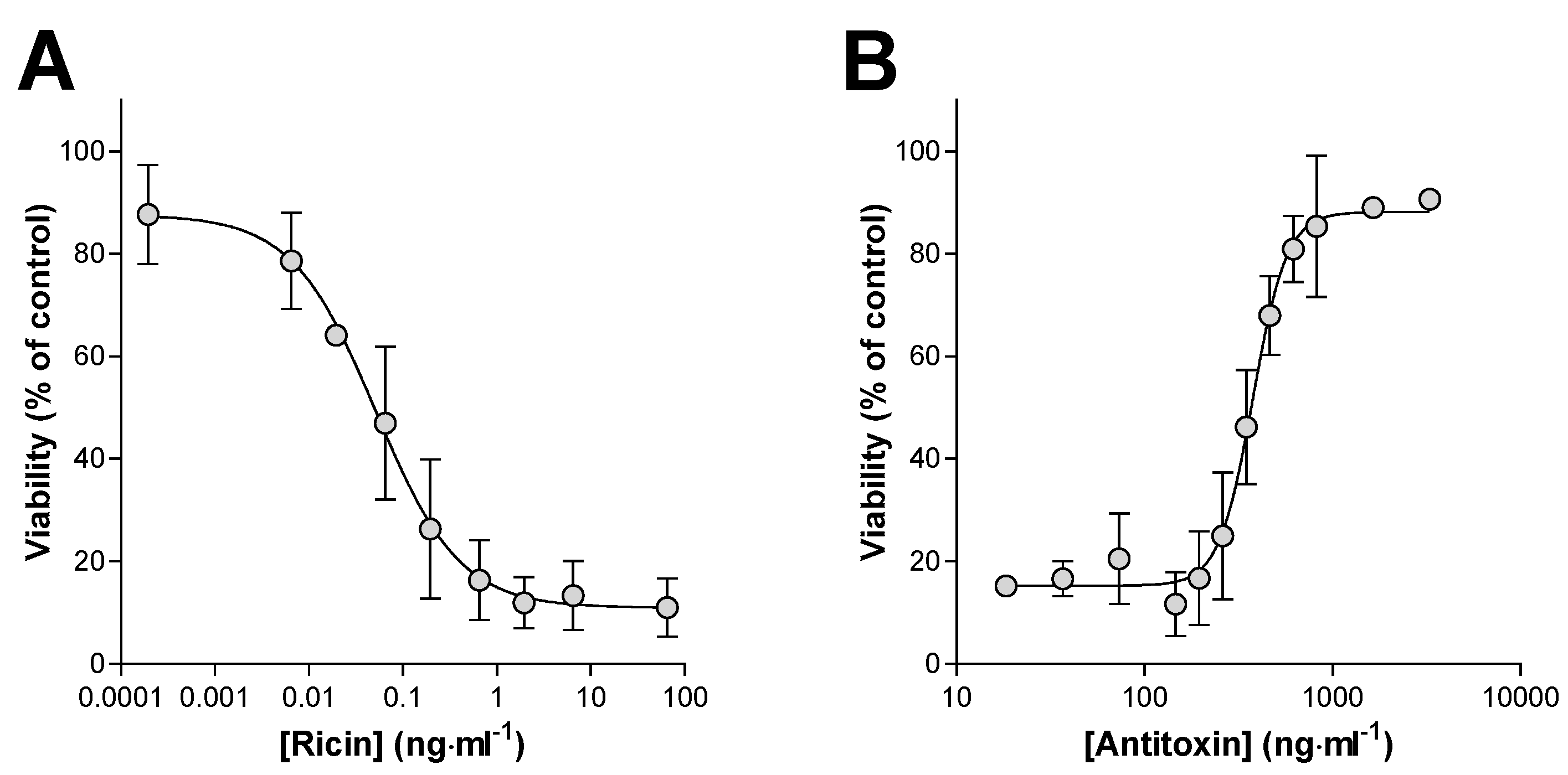
2.1. Determination of the Ricin Neutralising Activity of the Antitoxin In Vitro
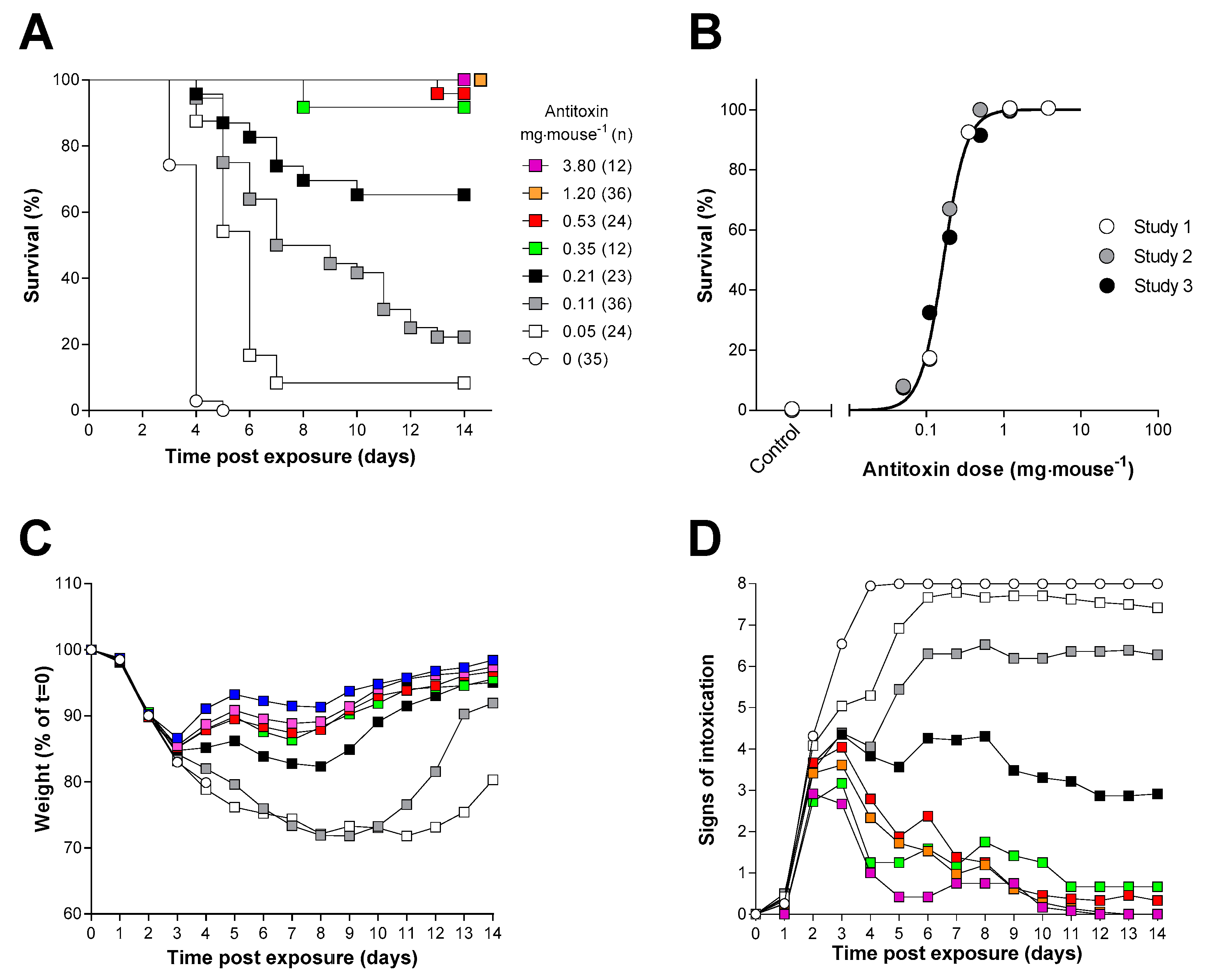
2.2. Determination of the Ricin Neutralising Activity of the Antitoxin In Vivo
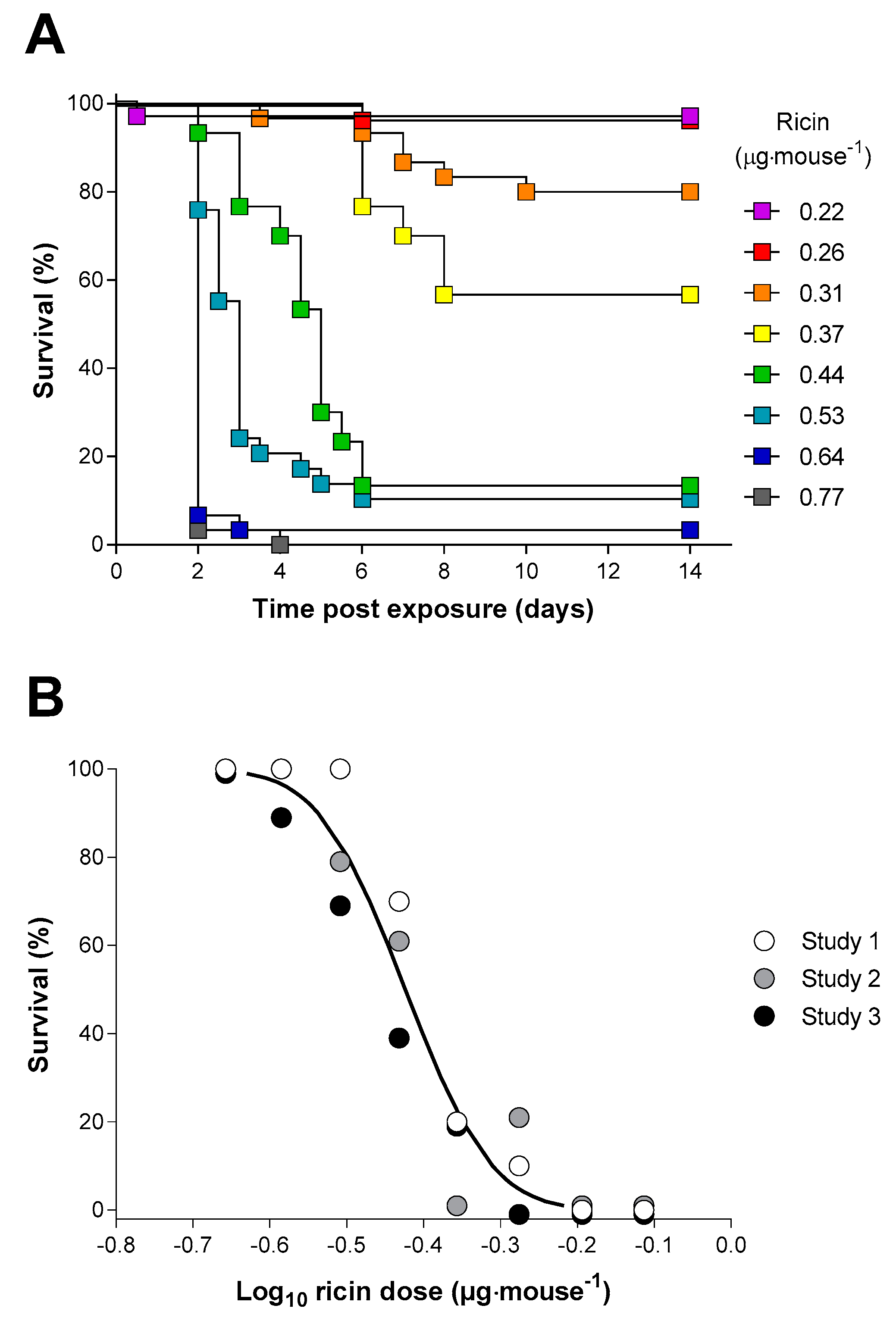
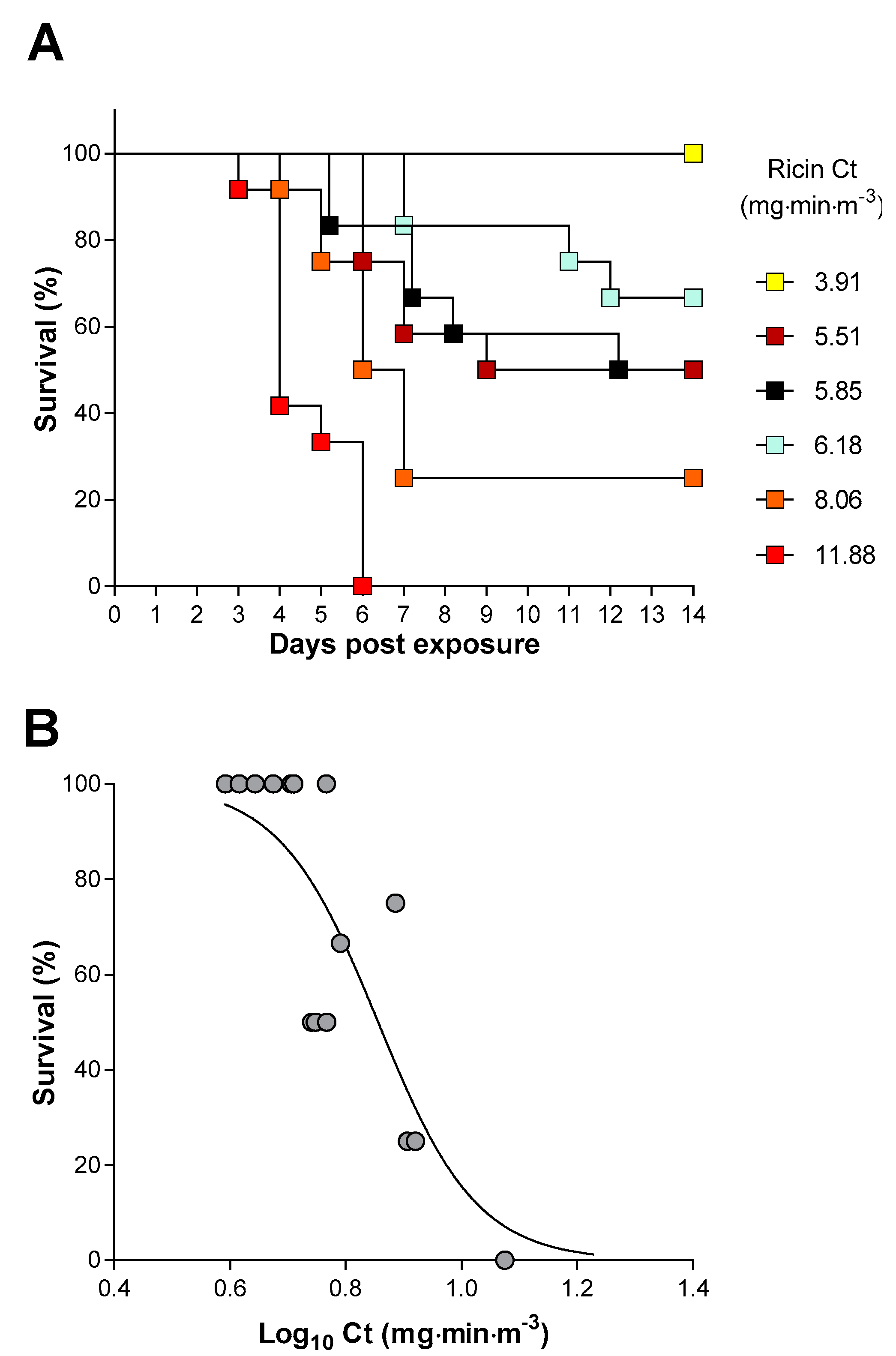
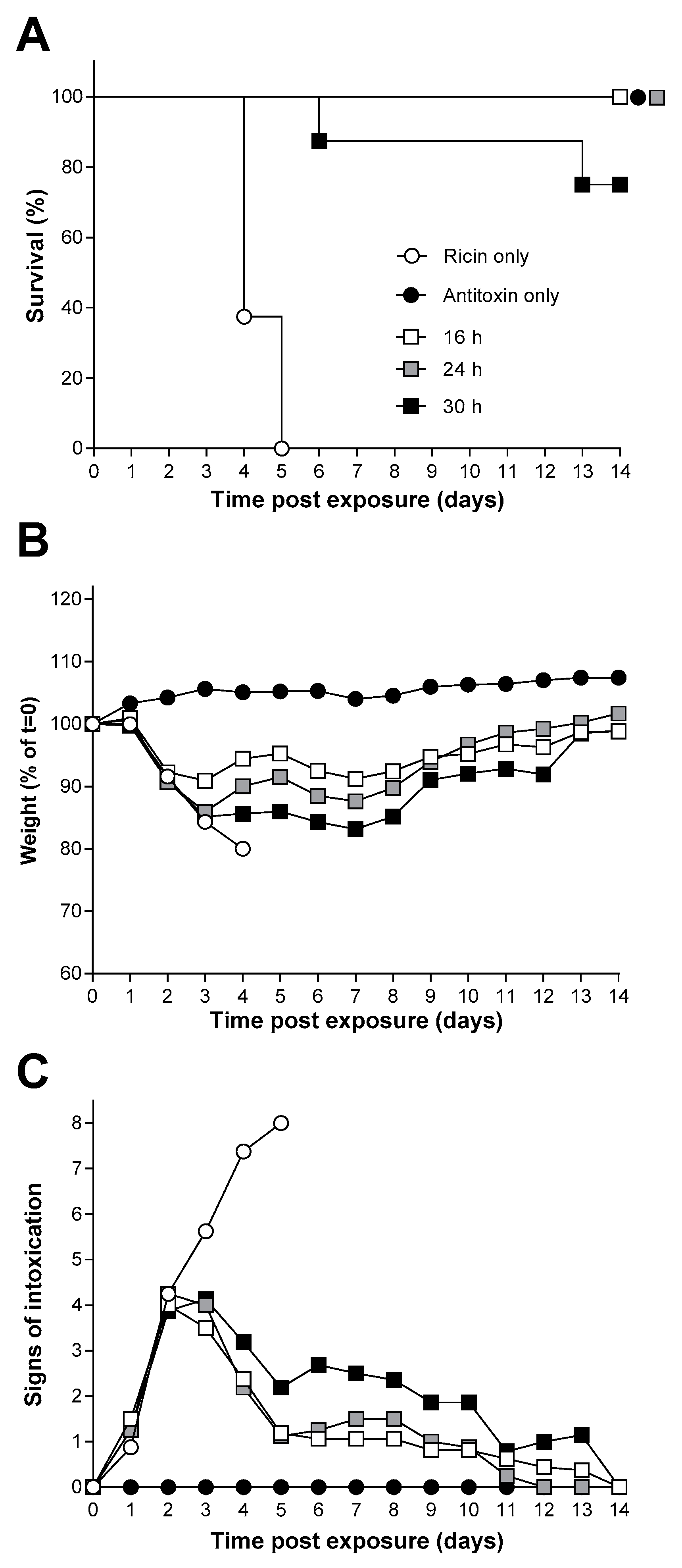
2.3. Inhalation Toxicity of Ricin in Balb/C Mice
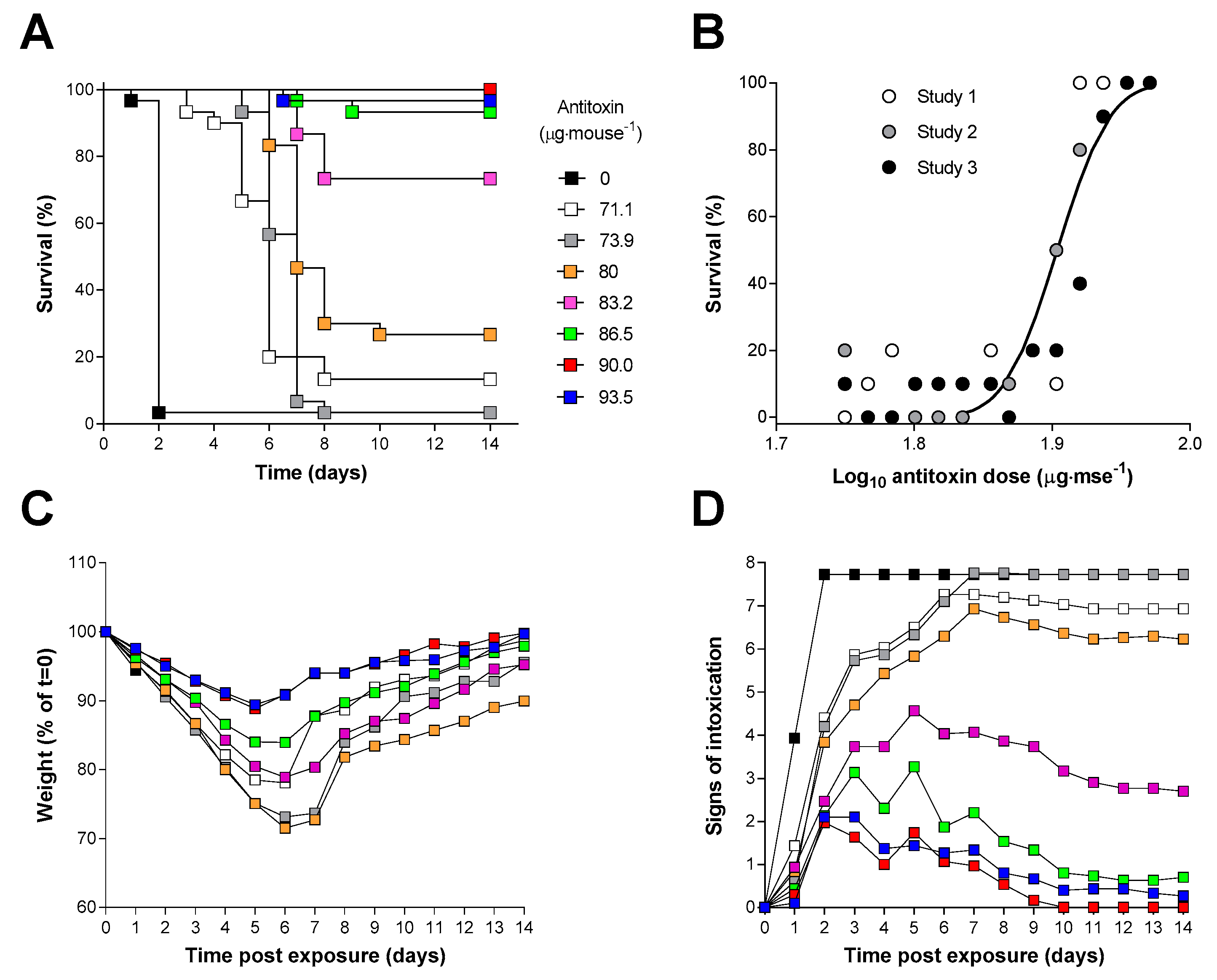
2.4. Antitoxin Protection against an Inhalational Ricin Challenge
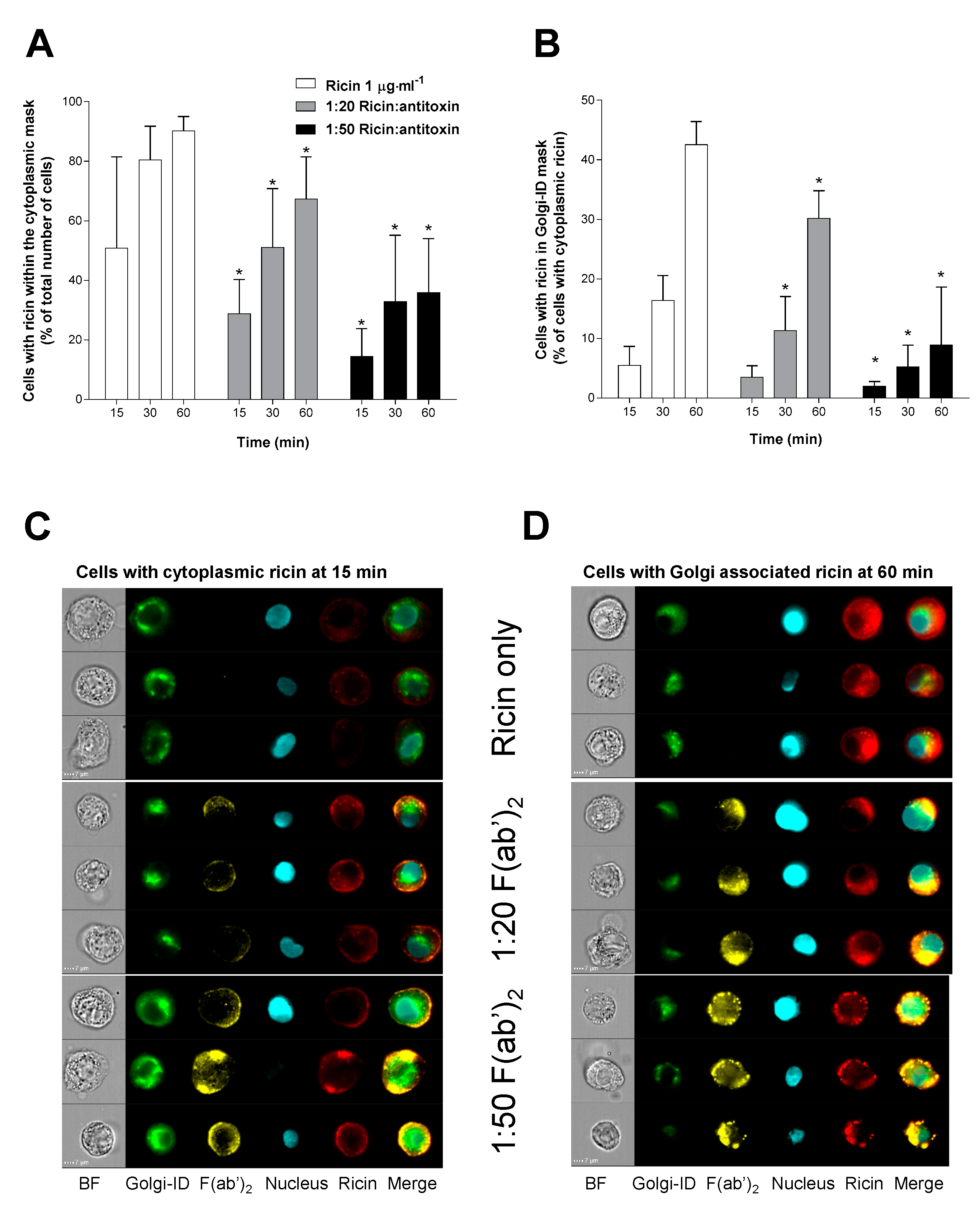
2.5. Antitoxin Can Affect the Uptake and Intracellular Trafficking of Ricin in A549 Cells
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Preparation of Ricin and Ricin Toxoid
5.2. Preparation of Ovine Ricin Antitoxin and Despeciated Antitoxin Fragment F(ab’)2
5.3. In Vitro Cytotoxicity Assay
5.4. In Vitro Neutralising Activity of Anti-Ricin Antitoxin
5.5. Animal Husbandry
5.6. In Vivo Neutralisation Assay in Mice
5.7. Determination of Inhalational Toxicity of Ricin in Mice
5.8. Efficacy of Ricin Antitoxins against Lethal Inhalation Ricin Challenge
5.9. Intracellular Trafficking of Ricin and Fluorochome Labelled F(ab’)2
5.10. Staining of Golgi Using Golgi-ID
5.11. Imaging Flow Cytometry Data Collection and Analysis
5.12. IFC Data Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Franz, D.R.; Jaax, N.K. Ricin toxin. In Medical Aspects of Chemical and Biological Warfare; Borden Institute, Walter Reed Army Medical Center: Washington, DC, USA, 1997; Chapter 32; pp. 631–642. [Google Scholar]
- Shoham, D. Iraqs biological warfare agents: A comprehensive analysis. Crit. Rev. Microbiol. 2000, 26, 179–204. [Google Scholar] [CrossRef] [PubMed]
- Roxas-Duncan, V.I.; Smith, L.A. Ricin: Perspective in bioterrorism. In Bioterrorism; Morse, S., Ed.; InTech: Rijeka, Croatia, 2012; Volume 7, pp. 133–158. [Google Scholar]
- Worbs, S.; Köhler, K.; Pauly, D.; Avondet, M.-A.; Schaer, M.; Dorner, M.B.; Dorner, B.G. Ricinus communis intoxications in human and veterinary medicine-a summary of real cases. Toxins 2011, 3, 1332–1372. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, G.D.; Phillips, G.J.; Holley, J.L. Inhalation toxicology of ricin preparations: Animal models, prophylactic and therapeutic approaches to protection. Inhal. Toxicol. 2007, 19, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.M.; Griffiths, G.D. Ricin: Chemistry, sources, exposures, toxicology and medical aspects. In General, Applied and Systems Toxicology; Wiley Ltd.: Chichester, UK, 2009. [Google Scholar] [CrossRef]
- Reisler, R.B.; Smith, L.A. The need for continued development of ricin countermeasures. Adv. Prev. Med. 2012, 1, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, L.; Baltelli, M.; Stirpe, F. Ribosomes-inactivating proteins from plants. Biochem. Biophys. Acta 1993, 1154, 237–282. [Google Scholar] [CrossRef]
- Lord, J.M.; Roberts, L.M.; Robertus, J.D. Ricin: Structure, mode of action and some current application. FASEB J. 1994, 8, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Rutenber, E.; Robertus, J.D. Structure of ricin B-chain at 2.5 A resolution. Proteins Struct. Funct. Genet. 1991, 10, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.; Waring, P. Macrophage adherence prevents apoptosis induced by ricin. Eur. J. Cell Biol. 1993, 62, 406–414. [Google Scholar] [PubMed]
- Simmons, B.M.; Stahl, P.D.; Russell, J.H. Mannose receptor mediated uptake of ricin toxin and ricin A-chain by macrophages. Multiple intracellular airways for A chain translocation. J. Biol. Chem. 1986, 261, 7912–7920. [Google Scholar] [PubMed]
- Magnusson, S.; Berg, T. Endocytosis of ricin by rat liver cells in vivo and in vitro is mainly mediated by mannose receptors on sinusoidal endothelial cells. Biochem. J. 1994, 291, 749–755. [Google Scholar] [CrossRef]
- Sapoznikov, A.; Falach, R.; Mazor, O.; Alcalay, R.; Gal, Y.; Seliger, N.; Sabo, T.; Kronman, C. Diverse profiles of ricin-cell interactions in the lung following intranasal exposure to ricin. Toxins 2015, 7, 4817–4831. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.M.; Smith, D.C.; Roberts, L.M. Toxin entry: How bacterial proteins get into mammalian cells. Cell Microbiol. 1991, 1, 85–91. [Google Scholar] [CrossRef]
- Robertus, J. The structure and action of ricin: A cytotoxic N-glycosidase. Cell Biol. 1991, 2, 23–30. [Google Scholar]
- Eiklid, K.; Olsnes, S.; Pihl, A. Entry of lethal doses of abrin, ricin and modeccin into the Cytosol of HeLa cells. Exp. Cell Res. 1980, 126, 321–326. [Google Scholar] [CrossRef]
- Olsnes, S.; Kozlov, J.V. Ricin. Toxicon 2001, 39, 1723–1728. [Google Scholar] [CrossRef]
- Soler-Rodriguez, A.-M.; Ghetie, M.-A.; Oppenheimer-Marks, N.; Uh, J.W.; Vitetta, E.S. Ricin A-chain and ricin A-chain immunotoxins rapidly damage human endothelial cells, Implications for vascular leak syndrome. Exp. Cell Res. 1993, 206, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Lindauer, M.L.; Wong, J.; Iwakura, Y.; Magun, B.E. Pulmonary inflammation triggered by ricin toxin requires macrophages and IL-1 signalling. J. Immunol. 2009, 183, 1419–1426. [Google Scholar] [CrossRef] [PubMed]
- Mabley, J.G.; Pacher, P.; Szabo, C. Activation of cholinergic anti-inflammatory pathway reduces ricin-induced mortality and organ failure in mice. Mol. Med. 2009, 15, 166–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gal, Y.; Mazor, O.; Alcalay, R.; Seliger, N.; Aftalion, M.; Sapoznikov, A.; Falach, R.; Kronman, C.; Sabo, T. Antibody/doxycycline combined therapy for pulmonary ricinosis: Attenuation of inflammation improves survival of ricin-intoxicated mice. Toxicol. Rep. 2014, 1, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Sabo, T.; Gal, Y.; Elhanany, E.; Sapoznikov, A.; Falach, R.; Mazor, O.; Kronman, C. Antibody treatment against pulmonary exposure to abrin confers significantly higher levels of protection than treatment against ricin intoxication. Toxicol. Lett. 2015, 237, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Roy, C.J.; Song, K.; Sivasubramani, S.K.; Gardner, D.J.; Pincus, S.H. Animal models of ricin toxicosis. Curr. Top. Microbiol. Immunol. 2012, 357, 243–257. [Google Scholar] [PubMed]
- Godal, A.; Fodstad, O.; Ingebrigtsen, K.; Pihl, A. Pharmacological studies of ricin in mice and humans. Cancer Chemother. Pharmacol. 1984, 13, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Fodstad, O.; Olsnes, S.; Pihl, A. Toxicity, distribution and elimination of the cancerostatic lectins abrin and ricin after parenteral injections into mice. Br. J. Cancer 1976, 32, 418–425. [Google Scholar] [CrossRef]
- Griffiths, G.D.; Leek, M.D.; Gee, D.J. The toxic plant proteins ricin and abrin induce apoptotic changes in mammalian lymphoid tissues and intestines. J. Pathol. 1987, 151, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Balint, G.A. Ricin: The toxic protein of castor oil seeds. Toxicology 1974, 2, 77–102. [Google Scholar] [CrossRef]
- Muldoon, D.F.; Stoh, S.J. Modulation of ricin toxicity in mice by biologically active substances. J. Appl. Toxicol. 1994, 14, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, G.D.; Rice, P.; Allenby, A.C.; Bailey, S.C.; Upshall, D.G. Inhalation toxicology and histopathology of ricin and abrin toxins. Inhal. Toxicol. 1995, 7, 269–288. [Google Scholar] [CrossRef]
- Bhaskaran, M.; Didier, P.J.; Sivasubramani, S.K.; Doyle, L.A.; Holley, J.; Roy, C.J. Pathology of lethal and sublethal doses of aerosolized ricin in Rhesus Macaques. Toxicol. Pathol. 2014, 42, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Mayers, C.N.; Holley, J.L.; Brooks, T. Antitoxin therapy for botulinum intoxication. Rev. Med. Microbiol. 2001, 12, 1–9. [Google Scholar] [CrossRef]
- Casadevall, A. Passive antibody administration as a specific defence against biological weapons. Emerg. Infect. Dis. 2002, 8, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P.; Goyffon, M. Venoms, anti-venoms and immunotherapy. Toxicon 1998, 36, 823–846. [Google Scholar] [CrossRef]
- Sedlacek, H.H.; Gronski, P.; Hofstaetter, E.J.; Kanzy, E.J.; Schlorlemmer, H.U.; Seiler, F.R. The biological properties of Immunoglobulin G and its split products (Fab’)2 and Fab. Klin. Wochenschr. 1983, 61, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Nydegger, U.E.; Sturzenegger, M. Adverse effects of intravenous immunoglobulinum therapy. Drug Saf. 1999, 21, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Morais, V.M.; Massaldi, H. Snake antivenoms: Adverse reactions and production technology. Venom. Anim. Toxins Incl. Trop. Dis. 2009, 15, 2–18. [Google Scholar] [CrossRef]
- O’Hara, J.M.; Whaley, K.; Pauly, M.; Zeitlin, L.C.; Mantis, N. Plant-based expression of a partially humanized neutralizing monoclonal IgG directed against an immunodominant epitope on the ricin toxin A subunit. Vaccine 2012, 30, 1239–1243. [Google Scholar] [CrossRef] [PubMed]
- Sully, E.K.; Whaley, K.J.; Bohorova, N.; Goodman, C.; Kim, D.H.; Pauly, M.H.; Velasco, J.; Hiatt, E.; Morton, J.; Swope, K.; et al. Chimeric plantibody passively protects mice against aerosolized ricin challenge. Clin. Vaccine Immunol. 2014, 21, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Van Slyke, G.; Sully, E.K.; Bohorova, N.; Bohorov, O.; Kim, D.; Pauly, M.; Whaley, K.; Zeitlin, L.; Mantis, N.J. A humanized monoclonal antibody that passively protects mice 3 against systemic and intranasal ricin toxin challenge. Clin. Vaccine Immunol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Pratt, T.S.; Pincus, S.H.; Hale, M.L.; Moreira, A.L.; Roy, C.J.; Tchou-Wong, K.-M. Oropharyngeal aspiration of ricin as a lung challenge model for evaluation of the therapeutic index of antibodies against ricin A chain for post-exposure treatment. Exp. Lung Res. 2007, 33, 459–481. [Google Scholar] [CrossRef] [PubMed]
- Noy-Porat, T.; Alcalay, R.; Epstein, E.; Sabo, T.; Kronman, C. Extended therapeutic window for post exposure treatment of ricin intoxication conferred by the use of high-affinity antibodies. Toxicon 2017, 127, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Miner, N.A.; Koehler, J. Intraperitoneal injection of mice. Appl. Microbiol. 1968, 16, 1418–1419. [Google Scholar]
- Noy-Porat, T.; Rosenfeld, R.; Ariel, N.; Epstein, E.; Alcalay, R.; Zvi, A.; Kronman, C.; Ordentlich, A.; Mazor, O. Isolation of anti-ricin protective antibodies exhibiting high affinity from immunized non-human primates. Toxins 2016, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, G.D.; Lindsay, C.D.; Upshall, D.G. Examination of the toxicity of several protein toxins of plant origin using bovine pulmonary endothelial cells. Toxicology 1994, 90, 11–27. [Google Scholar] [CrossRef]
- Yermakova, A.; Mantis, N.J. Protective immunity to ricin toxin conferred by antibodies against the toxin’s binding subunit (RTB). Vaccine 2011, 29, 7925–7935. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Hu, W.-G.; Yin, J.; Chau, D.; Hu, C.C.; Lillico, D.; Yu, J.; Negrych, L.M.; Cherwonogrodzky, J.W. Conformation-dependent high-affinity potent ricin-neutralizing monoclonal antibodies. Biomed. Res. Int. 2013. [Google Scholar] [CrossRef]
- Hu, W.G.; Yin, J.; Chau, D.; Negreych, L.M.; Cherwonogrodzky, J.W. Humanization and characterization of an anti-ricin neutralization monoclonal antibody. PLoS ONE 2012, 7, e45595. [Google Scholar] [CrossRef] [PubMed]
- Prigent, J.; Panigai, L.; Lamourette, P.; Sauvaire, D.; Devilliers, K.; Plaisance, M.; Volland, H.; Creminon, C.; Simon, S. Neutralising antibodies against ricin toxin. PLoS ONE 2009, 6, e20166. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.; Klokk, T.; Cole, R.; Sandvig, K.; Mantis, N.J. A bispecific antibody promotes aggregation of ricin toxin on cell surfaces and alters dynamics of toxin internalization and trafficking. PLoS ONE 2016, 11, e0156893. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Mize, R.R.; Marrero, L.; Corti, M.; Kirk, J.M.; Pincus, S.H. Antibody to ricin A chain hinders intracellular routing of toxin and protects cells even after toxin has been internalized. PLoS ONE 2013, 8, e62417. [Google Scholar] [CrossRef] [PubMed]
- Yermakova, A.; Klokk, T.I.; Cole, R.; Sandvig, K.; Mantis, N.J. Antibody-mediated inhibition of ricin toxin retrograde transport. mBio 2014, 5, e00995. [Google Scholar] [CrossRef] [PubMed]
- Yermakova, A.; Klokk, T.I.; O’Hara, J.M.; Cole, R.; Sandvig, K.; Mantis, N.J. Neutralizing monoclonal antibodies against disparate epitopes on ricin toxin’s enzymatic subunit interfere with intracellular toxin transport. Sci. Rep. 2016, 6, 22721. [Google Scholar] [CrossRef] [PubMed]
- Gawarammana, I.; Keyler, D. Dealing with adverse reactions to snake venom. Ceylon Med. J. 2011, 56, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Theakston, R.D.G.; Smith, D.C. Therapeutic antibodies to snake venoms. In Therapeutic Antibodies; Landon, J., Chard, T., Eds.; Springer: London, UK, 1995; Chapter 6; pp. 109–133. [Google Scholar]
- Mayers, C.N.; Veall, S.; Bedford, R.J.; Holley, J.L. Anti-immunoglobulin responses to IgG, F(ab’)2 and Fab botulinum antitoxins in mice. Immunopharmacol. Immunotoxicol. 2003, 25, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Holley, J.L.; Poole, S.J.C.; Cooper, I.A.M.; Griffiths, G.D.; Simpson, A.J. The production and evaluation of ricin antitoxin. In Defence against the Effects of Chemical Hazards: Toxicology, Diagnosis and Medical Countermeasures; NATO Meeting Proceedings RTO-MP-HFM-149; NATO: Neuilly-sur-Seine, France, 2007; pp. 12-1–12-8. [Google Scholar]
- Sesardic, D.; Winsnes, R.; Rigsby, P.; Beh-Gross, M.-E. Collaborative study for the calibration of serological methods for potency testing of diphtheria toxoid vaccines. Extended study: Correlation of serology with in vivo toxin neutralisation. Pharmeuropa Bio 2003, 2, 69–75. [Google Scholar]
- First WHO International Standard for Human Diphtheria Antitoxin; WHO Expert Committee on Biological Standardization: 63rd Report; World Health Organization: Geneva, Switzerland, 2013; Section 4.1.3; p. 29.
- Stechmann, B.; Bai, S.K.; Gobbo, E.; Lopez, R.; Merer, G.; Pinchard, S.; Panigai, L.; Tenza, D.; Raposo, G.; Beaumelle, B.; et al. Inhibition of retrograde transport protects mice from lethal ricin challenge. Cell 2010, 141, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Barbier, J.; Bouclier, C.; Johannes, L.; Gillet, D. Inhibitors of the cellular trafficking of ricin. Toxins 2012, 4, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Pons, V.; Noel, R.; Buisson, D.A.; Uisson, D.A.; Michau, A.; Johannes, L.; Gillet, D.; Barbier, J.; Cintrat, J.-C. (S)-N-Methyldihydroquinazolinones are the Active Enantiomers of Retro-2 Derived Compounds against toxins. ACS Med. Chem. Lett. 2014, 5, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Gal, Y.; Sapoznikov, A.; Falach, R.; Ehrlich, S.; Aftalion, M.; Sabo, T.; Kronman, C. Potent antiedematous and protective effects of ciprofloxacin in pulmonary ricinosis. Antimicrob. Agents Chemother. 2016, 60, 7153–7158. [Google Scholar] [PubMed]
- Thullier, P.; Griffiths, G.D. Broad recognition of ricin toxins prepared from a range of Ricinus cultivars using immunochomatographic tests. Clin. Toxicol. 2009, 47, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Pugsley, H.R. Quantifying autophagy: Measuring LC3 puncta and autolysosome formation in cells using multispectral imaging flow cytometry. Methods 2017, 112, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Rajan, R.; Karbowniczek, M.; Pugsley, H.R.; Sabnani, M.K.; Astrinidis, A.; La-Beck, N.M. Quantifying autophagosomes and autolysosomes in cells using imaging flow cytometry. Cytometry A 2015, 87, 451–458. [Google Scholar] [CrossRef] [PubMed]







© Crown copyright (2017), Dstl. This material is licensed under the terms of the Open Government Licence except where otherwise stated. To view this licence, visit: http://www.nationalarchives.gov.uk/doc/open-government-licence/version/3 or write to the Information Policy Team, The National Archives, Kew, London TW9 4DU, or email: [email protected].
Share and Cite
Whitfield, S.J.C.; Griffiths, G.D.; Jenner, D.C.; Gwyther, R.J.; Stahl, F.M.; Cork, L.J.; Holley, J.L.; Green, A.C.; Clark, G.C. Production, Characterisation and Testing of an Ovine Antitoxin against Ricin; Efficacy, Potency and Mechanisms of Action. Toxins 2017, 9, 329. https://doi.org/10.3390/toxins9100329
Whitfield SJC, Griffiths GD, Jenner DC, Gwyther RJ, Stahl FM, Cork LJ, Holley JL, Green AC, Clark GC. Production, Characterisation and Testing of an Ovine Antitoxin against Ricin; Efficacy, Potency and Mechanisms of Action. Toxins. 2017; 9(10):329. https://doi.org/10.3390/toxins9100329
Chicago/Turabian StyleWhitfield, Sarah J. C., Gareth D. Griffiths, Dominic C. Jenner, Robert J. Gwyther, Fiona M. Stahl, Lucy J. Cork, Jane L. Holley, A. Christopher Green, and Graeme C. Clark. 2017. "Production, Characterisation and Testing of an Ovine Antitoxin against Ricin; Efficacy, Potency and Mechanisms of Action" Toxins 9, no. 10: 329. https://doi.org/10.3390/toxins9100329
APA StyleWhitfield, S. J. C., Griffiths, G. D., Jenner, D. C., Gwyther, R. J., Stahl, F. M., Cork, L. J., Holley, J. L., Green, A. C., & Clark, G. C. (2017). Production, Characterisation and Testing of an Ovine Antitoxin against Ricin; Efficacy, Potency and Mechanisms of Action. Toxins, 9(10), 329. https://doi.org/10.3390/toxins9100329