
Rapid Assessment of the Toxicity of Fungal Compounds Using Luminescent Vibrio qinghaiensis sp. Q67




Abstract
:1. Introduction
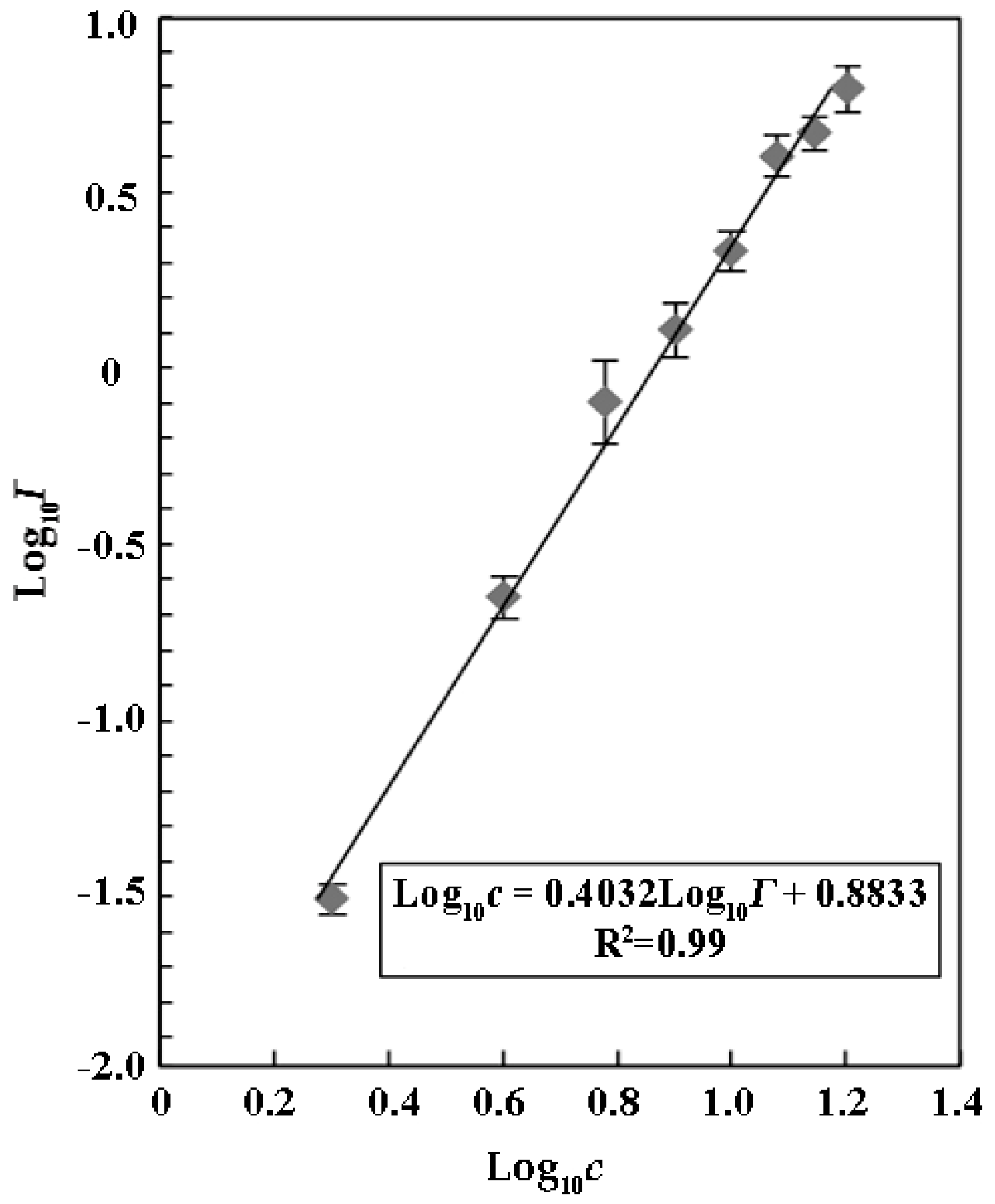
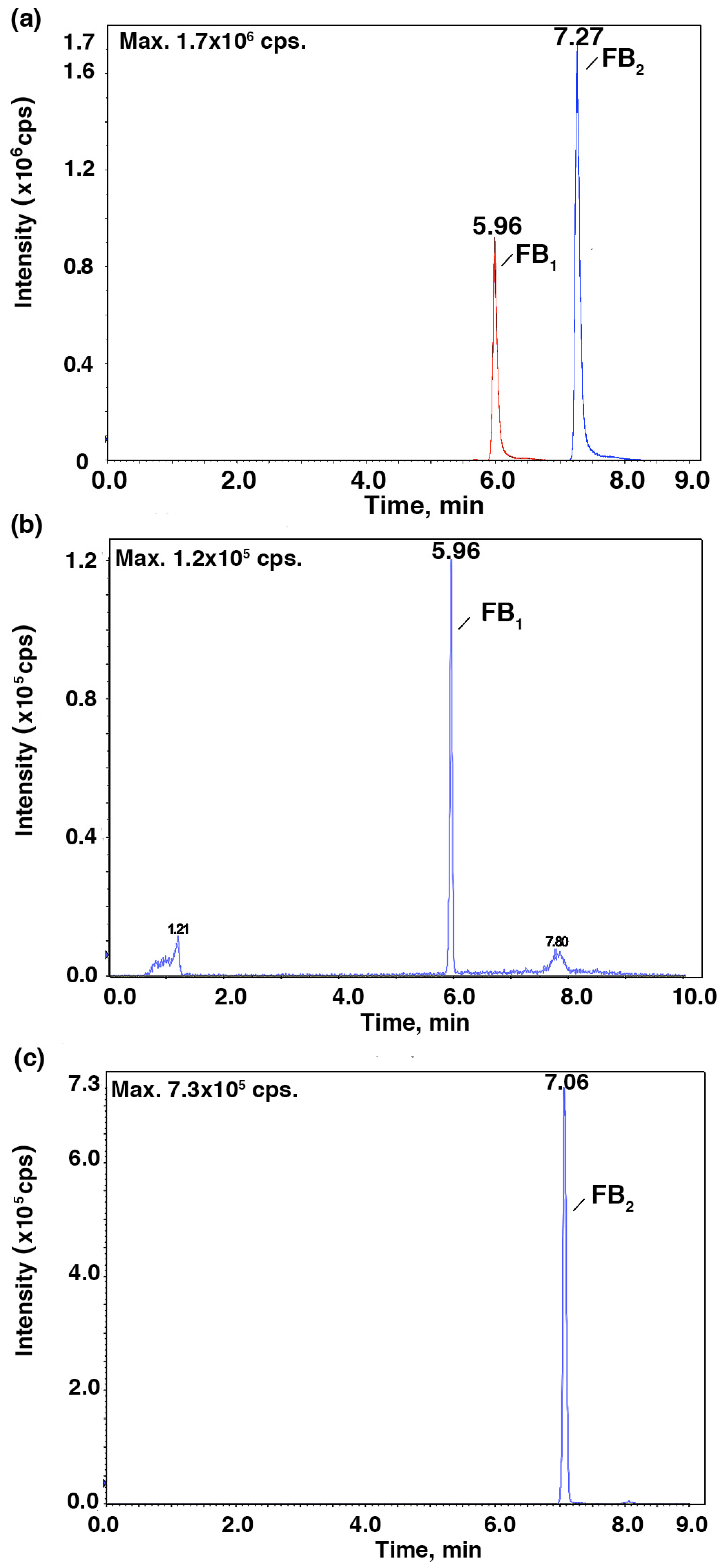
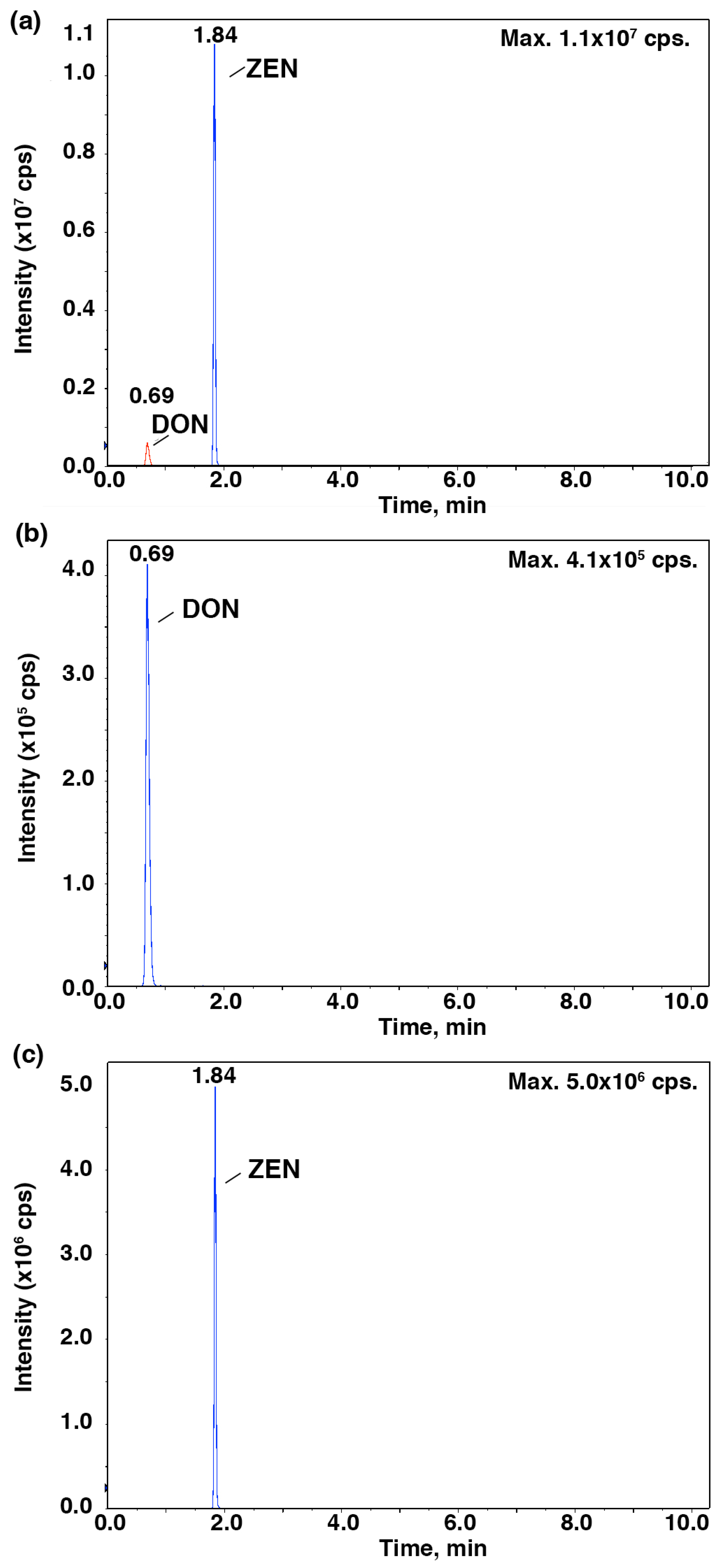
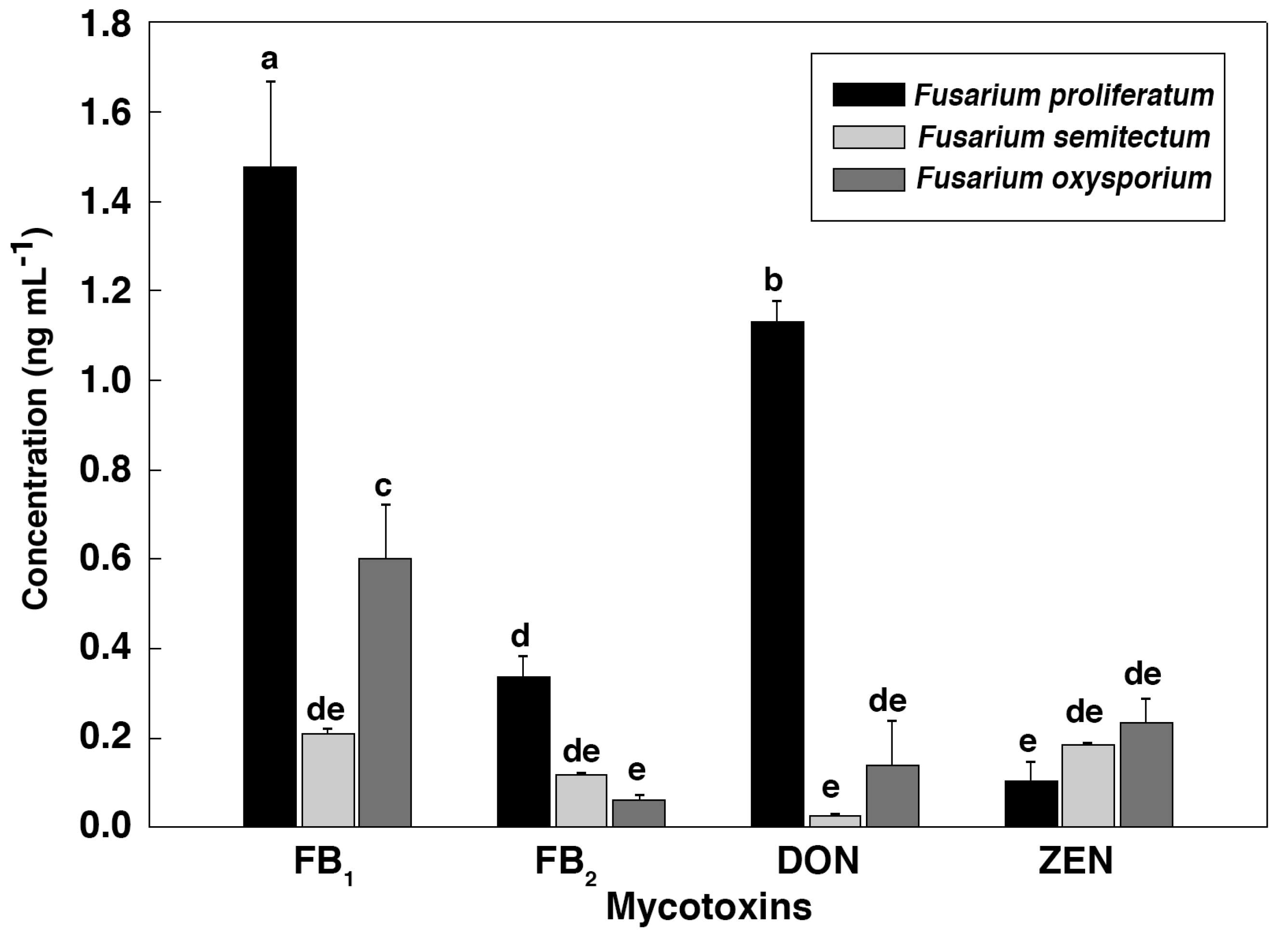
2. Results and Discussion
3. Materials and Methods
3.1. Chemicals and Solutions
3.2. Fungal Fermentation
3.3. Luminescence Inhibition Assays
3.4. Mycotoxin Extraction
3.5. HPLC-MS/MS
3.6. Calculation of a Relationship between Concentration and Effect
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Drusch, S.; Ragab, W. Mycotoxins in fruits, fruit juices, and dried fruits. J. Food Prot. 2003, 66, 1514–1527. [Google Scholar] [CrossRef] [PubMed]
- Wambacq, E.; Vanhoutte, I.; Audenaert, K.; De Gelder, L.; Haesaert, G. Occurrence, prevention and remediation of toxigenic fungi and mycotoxins in silage: A review. J. Sci. Food Agric. 2016, 96, 2284–2302. [Google Scholar] [CrossRef] [PubMed]
- Drusch, S.; Aumann, J. Mycotoxins in fruits: Microbiology, occurrence, and changes during fruit processing. Adv. Food Nutr. Res. 2005, 50, 33–78. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Tussell, R.; Quijano-Ramayo, A.; Cortes-Velazquez, A.; Lappe, P.; Larque-Saavedra, A.; Perez-Brito, D. PCR-based detection and characterization of the fungal pathogens Colletotrichum gloeosporioides and Colletotrichum capsici causing anthracnose in papaya (Carica papaya L.) in the Yucatan peninsula. Mol. Biotechnol. 2008, 40, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Drenth, A.; Guest, D.I. Fungal and oomycete diseases of tropical tree fruit crops. Annu. Rev. Phytopathol. 2016, 54, 373–395. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Li, J.; Jiang, Y.M.; Duan, X.W.; Qu, H.X.; Yang, B.; Chen, F.; Sivakumar, D. Natural occurrence, analysis and prevention of mycotoxins in fruits and their processed products. Crit. Rev. Food Sci. Nutr. 2014, 54, 64–83. [Google Scholar] [CrossRef] [PubMed]
- Chilaka, C.A.; De Boevre, M.; Atanda, O.O.; De Saeger, S. The status of Fusarium mycotoxins in sub-Saharan africa: A review of emerging trends and post-harvest mitigation strategies towards food control. Toxins 2017, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Stoev, S.D. Foodborne mycotoxicoses, risk assessment and underestimated hazard of masked mycotoxins and joint mycotoxin effects or interaction. Environ. Toxicol. Pharmacol. 2015, 39, 794–809. [Google Scholar] [CrossRef] [PubMed]
- Flajs, D.; Domijan, A.M.; Ivić, D.; Cvjetković, B.; Peraica, M. ELISA and HPLC analysis of ochratoxin A in red wines of Croatia. Food Control 2009, 20, 590–592. [Google Scholar] [CrossRef]
- Gong, L.; Jiang, Y.M.; Chen, F. Molecular strategies for detection and quantification of mycotoxin-producing Fusarium species: A review. J. Sci. Food Agric. 2015, 95, 1767–1776. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.D.; Wong, J.W.; Zhang, K.; Hayward, D.G.; Lee, N.S.; Trucksess, M.W. Multi-mycotoxin analysis of finished grain and nut products using high-performance liquid chromatography–triple-quadrupole mass spectrometry. J. Agric. Food Chem. 2013, 61, 4771–4782. [Google Scholar] [CrossRef] [PubMed]
- Escrivá, L.; Font, G.; Manyes, L. In vivo toxicity studies of Fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, H.; Nakagawa, H. Differences in the toxicities of trichothecene mycotoxins, deoxynivalenol and nivalenol, in cultured cells. Jpn. Agric. Res. Q. 2014, 48, 393–397. [Google Scholar] [CrossRef]
- Kouadio, J.H.; Moukha, S.; Brou, K.; Gnakri, D. Lipid metabolism disorders, lymphocytes cells death, and renal toxicity induced by very low levels of deoxynivalenol and fumonisin B1 alone or in combination following 7 days oral administration to mice. Toxicol. Int. 2013, 20, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Jiang, Z.; Zhao, Q.; Zhang, Z.; Su, H.; Gao, X.; Ye, Z. Acute toxicity assessment of explosive-contaminated soil extracting solution by luminescent bacteria assays. Environ. Sci. Pollut. Res. Int. 2016, 23, 22803–22809. [Google Scholar] [CrossRef] [PubMed]
- Kratasyuk, V.A.; Esimbekova, E.N. Applications of luminous bacteria enzymes in toxicology. Comb. Chem. High Throughput Screen. 2015, 18, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Zabar, R.; Sarakha, M.; Chung, P.W.; Trebše, P. Stability of pesticides’ residues under ultraviolet germicidal irradiation. Acta Chim. Slov. 2011, 58, 326–332. [Google Scholar] [PubMed]
- Sarter, S.; Zakhia, N. Chemiluminescent and bioluminescent assays as innovative prospects for mycotoxin determination in food and feed. Luminescence 2004, 19, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, E.; Deluca, M. Studies of the control of luminescence in Beneckeaharveyi: Properties of the NADH and NADPH:FMN oxidoreductases. Biochemistry 1978, 17, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Dams, R.I.; Biswas, A.; Olesiejuk, A.; Fernandes, T.; Christofi, N. Silver nanotoxicity using a light-emitting biosensor Pseudomonas putida isolated from a wastewater treatment plant. J. Hazard. Mater. 2011, 195, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jiang, G.X.; Yang, B.; Dong, X.H.; Feng, L.Y.; Lin, S.; Chen, F.; Ashraf, M.; Jiang, Y.M. A luminescent bacterium assay of fusaric acid produced by Fusarium proliferatum from banana. Anal. Bioanal. Chem. 2012, 402, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, S.S.; Dou, R.N.; Liu, H.L.; Li, X.L. Toxicities of 30 ionic liquids to Vibrio qinghaiensis sp. Q67. Environ. Sci. 2011, 32, 1108–1113. [Google Scholar]
- Mo, L.Y.; Liu, S.S.; Zhu, Y.N.; Liu, H.L.; Liu, H.Y.; Yi, Z.S. Combined toxicity of the mixtures of phenol and aniline derivatives to Vibrio qinghaiensis sp.-Q67. Bull. Environ. Contam. Toxicol. 2011, 87, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Ding, K.K.; Lu, L.P.; Wang, J.Y.; Wang, J.P.; Zhou, M.Q.; Zheng, C.W.; Liu, J.S.; Zhang, C.L.; Zhuang, S.L. In vitro and in silico investigations of the binary-mixture toxicity of phthalate esters and cadmium (II) to Vibrio qinghaiensis sp. Q67. Sci. Total Environ. 2017, 580, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Menz, J.; Schneider, M.; Kümmerer, K. Toxicity testing with luminescent bacteria—Characterization of an automated method for the combined assessment of acute and chronic effects. Chemosphere 2013, 93, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.L.; Liu, S.S.; Su, B.X.; Qin, L.T. Predicting synergistic toxicity of heavy metals and ionic liquids on photobacterium Q67. J. Hazard. Mater. 2014, 268, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, S.S.; Yu, Z.Y.; Liu, H.L.; Zhang, J. The time-dependent hormetic effects of 1-alkyl-3-methylimidazolium chloride and their mixtures on Vibrio qinghaiensis sp. Q67. J. Hazard. Mater. 2013, 258–259, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Yates, I.E.; Porter, J.K. Bacterial bioluminescence as a bioassay for mycotoxins. Appl. Environ. Microbiol. 1982, 44, 1072–1075. [Google Scholar] [PubMed]
- Bouslimi, A.; Bouaziz, C.; Ayed-Boussema, I.; Hassen, W.; Bacha, H. Individual and combined effects of ochratoxin A and citrinin on viability and DNA fragmentation in cultured vero cells and on chromosome aberrations in mice bone marrow cells. Toxicology 2008, 251, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Krifaton, C.; Kriszt, B.; Szoboszlay, S.; Cserháti, M.; Szucs, A.; Kukolya, J. Analysis of aflatoxin-B1-degrading microbes by use of a combined toxicity-profiling method. Mutat. Res. 2011, 726, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kratasyuk, V.; L’Vova, L.S.; Egorova, O.I.; Kudryasheva, N.S.; Orlova, N.Yu.; Bytev, V.O. Effect of Fusarium mycotoxins on a bacterial bioluminescence system in vitro. Appl. Biochem. Microbiol. 1998, 34, 207–209. [Google Scholar]
- Gastélum-Martínez, E.; Compant, S.; Taillandier, P.; Mathieu, F. Control of T-2 toxin in Fusarium langsethiae and Geotrichum candidum co-culture. Arhiv za Higijenu Rada i Toksikologiju 2013, 63, 447–456. [Google Scholar] [CrossRef] [Green Version]
- Logrieco, A.; Moretti, A.; Ritieni, A.; Bottalico, A.; Corda, P. Occurrence and toxigenicity of Fusarium proliferatum from preharvest maize ear rot, and associated mycotoxins, in Italy. Plant Dis. 1995, 79, 727–731. [Google Scholar] [CrossRef]
- Mikušová, P.; Šrobárová, A.; Sulyok, M.; Santini, A. Fusarium fungi and associated metabolites presence on grapes from Slovakia. Mycotoxin Res. 2013, 29, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, M.; Huerta, T.; Mateo, R. Mycotoxin production by Fusarium species isolated from bananas. Appl. Environ. Microbiol. 1997, 63, 364–369. [Google Scholar] [PubMed]
- Xie, H.H.; Liang, Y.G.; Xue, J.H.; Xu, Q.L.; Jiang, Y.M.; Wei, X.Y. Secondary metabolites of the phytopathogen Peronophythora litchii. Nat. Prod. Commun. 2010, 5, 245–248. [Google Scholar] [PubMed]
- Frisvad, J.C.; Smedsgaard, J.; Larsen, T.O.; Samson, R.A. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud. Mycol. 2004, 49, 201–241. [Google Scholar]
- García-Pajón, C.M.; Collado, I.G. Secondary metabolites isolated from Colletotrichum species. Nat. Prod. Rep. 2003, 20, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.J.; An, C.H.; Han, J.R. Wet-plate culture studies of Penicillium sp. PT95 and Q1 for mass production of sclerotia. J. Basic Microbiol. 2014, 54, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.T.; Lee, Y.R.; Jin, J.M.; Han, K.H.; Kim, H.; Kim, J.C.; Lee, T.; Yun, S.H.; Lee, Y.W. Two different polyketide synthase genes are required for synthesis of zearalenone in Gibberella zeae. Mol. Microbiol. 2005, 58, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Waśkiewicz, A.; Beszterda, M.; Bocianowski, J.; Goliński, P. Natural occurrence of fumonisins and ochratoxin A in some herbs and spices commercialized in Poland analyzed by UPLC-MS/MS method. Food Microbiol. 2013, 36, 426–431. [Google Scholar] [CrossRef] [PubMed]
- International Organization for Standardization. Water Quality—Determination of the Inhibitory Effect of Water Samples on the Light Emission of Vibrio fischeri (Luminescent Bacteria Test)—Part 3: Method Using Freeze-Dried Bacteria; ISO 11348-3:2007; International Organization for Standardization: Geneva, Switzerland, 2007. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycotoxins | Formula | R2 | IC50 (μg mL−1) |
|---|---|---|---|
| Fumonisin B1 | 0.98 | 14.88 ± 0.14 | |
| Deoxynivalenol | 0.99 | 30.74 ± 0.33 | |
| Zearalenone | 0.98 | 9.71 ± 0.17 | |
| Ochratoxin A | 0.99 | 11.02 ± 0.10 | |
| Patulin | 0.99 | 13.31 ± 0.14 | |
| Citrinin | 0.99 | 12.23 ± 0.10 |
| Fungus | Original | Formula | R2 | IC50/% |
|---|---|---|---|---|
| Fusarium proliferatum | Averrhoa carambola L. | 0.98 | 17.49 ± 2.15 | |
| Fusarium semitectum | Musa nana Lour. | 0.98 | 92.56 ± 11.20 | |
| Fusarium oxysporium | Musa nana Lour. | 0.98 | 33.33 ± 5.67 | |
| Fusarium verticillioides | Litchi chinensis Sonn. | 0.99 | 28.61 ± 5.40 | |
| Colletotrichum gloeosporioides | Mangifera indica L. | 0.97 | 115.90 ± 19.13 | |
| Peronophythora litchii | Litchi chinensis Sonn. | 0.99 | 69.45 ± 9.24 | |
| Penicillium italicum | Citrus reticulata Blanco | N/A | ||
| Penicillium digitatum | Citrus reticulata Blanco | 0.97 | 213.50 ± 46.08 | |
| Geotrichum candidum | Citrus reticulata Blanco | N/A | ||
| Phytophthora infestans | Musa nana Lour. | 0.99 | 119.51 ± 12.22 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jian, Q.; Gong, L.; Li, T.; Wang, Y.; Wu, Y.; Chen, F.; Qu, H.; Duan, X.; Jiang, Y. Rapid Assessment of the Toxicity of Fungal Compounds Using Luminescent Vibrio qinghaiensis sp. Q67. Toxins 2017, 9, 335. https://doi.org/10.3390/toxins9100335
Jian Q, Gong L, Li T, Wang Y, Wu Y, Chen F, Qu H, Duan X, Jiang Y. Rapid Assessment of the Toxicity of Fungal Compounds Using Luminescent Vibrio qinghaiensis sp. Q67. Toxins. 2017; 9(10):335. https://doi.org/10.3390/toxins9100335
Chicago/Turabian StyleJian, Qijie, Liang Gong, Taotao Li, Yong Wang, Yu Wu, Feng Chen, Hongxia Qu, Xuewu Duan, and Yueming Jiang. 2017. "Rapid Assessment of the Toxicity of Fungal Compounds Using Luminescent Vibrio qinghaiensis sp. Q67" Toxins 9, no. 10: 335. https://doi.org/10.3390/toxins9100335
APA StyleJian, Q., Gong, L., Li, T., Wang, Y., Wu, Y., Chen, F., Qu, H., Duan, X., & Jiang, Y. (2017). Rapid Assessment of the Toxicity of Fungal Compounds Using Luminescent Vibrio qinghaiensis sp. Q67. Toxins, 9(10), 335. https://doi.org/10.3390/toxins9100335