
Targeting Metastasis with Snake Toxins: Molecular Mechanisms


Abstract
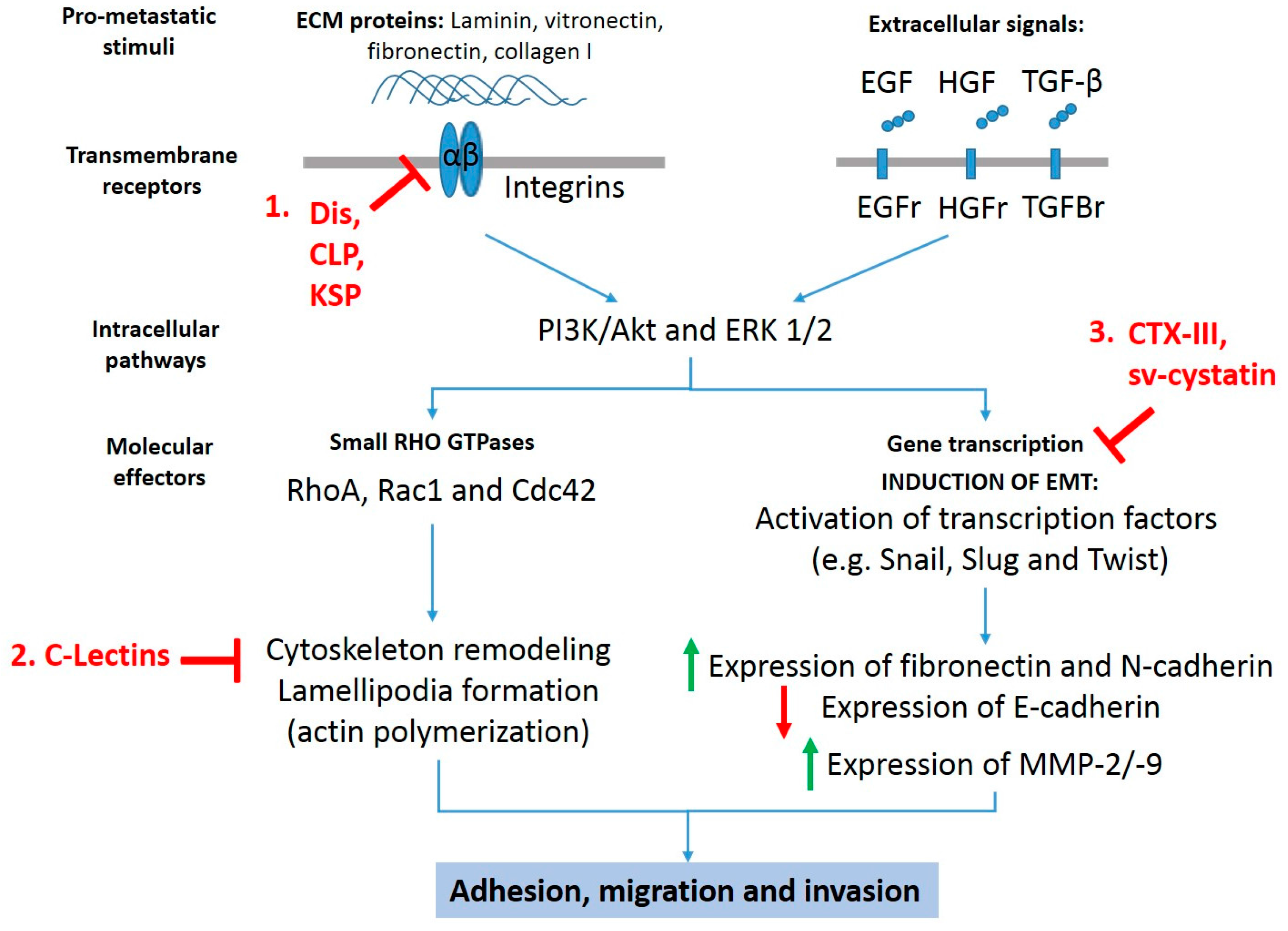
:1. Introduction
2. Snake Toxins as Inhibitors of Cancer Metastasis
3. Inhibition of Extracellular Matrix Component-Dependent Adhesion and Migration
4. Inhibition of Epithelial–Mesenchymal Transition
5. Alterations in the Actin/Cytoskeleton Network
6. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gandalovičová, A.; Rosel, D.; Fernandes, M.; Veselý, P.; Heneberg, P.; Čermák, V.; Petruželka, L.; Kumar, S.; Sanz-Moreno, V.; Brábek, J. Migrastatics-Anti-metastatic and anti-invasion drugs: Promises and challenges. Trends Cancer 2017, 3, 391–406. [Google Scholar] [CrossRef] [PubMed]
- Riggi, N.; Aguet, M.; Stamenkovic, I. Cancer metastasis: A reappraisal of its underlying mechanisms and their relevance to treatment. Annu. Rev. Pathol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Celià-Terrassa, T.; Kang, Y. Distinctive properties of metastasis-initiating cells. Genes Dev. 2016, 30, 892–908. [Google Scholar] [CrossRef] [PubMed]
- Tomaskovic-Crook, E.; Thompson, E.; Thiery, J. Epithelial to mesenchymal transition and breast cancer. Breast Cancer Res. 2009, 11, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M.; Gonzalez, S.; Welin, J.; Fuxe, J. Epithelial-mesenchymal transition in cancer metastasis through the lymphatic system. Mol. Oncol. 2017, 11, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Lambert, A.; Pattabiraman, D.; Weinberg, R. Emerging biological principles of metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [PubMed]
- Sleeman, J.; Steeg, P. Cancer metastasis as a therapeutic target. Eur. J. Cancer 2010, 46, 1177–1180. [Google Scholar] [CrossRef] [PubMed]
- Heath, A.S.; Yibin, K. Determinants of organotropic metastasis. Annu. Rev. Cancer Biol. 2017, 1, 403–423. [Google Scholar]
- Aranda-Souza, M.; Rossato, F.; Costa, R.; Figueira, T.; Castilho, R.; Guarniere, M.; Nunes, E.; Coelho, L.; Correia, M.; Vercesi, A. A lectin from Bothrops leucurus snake venom raises cytosolic calcium levels and promotes B16-F10 melanoma necrotic cell death via mitochondrial permeability transition. Toxicon 2014, 82, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, K.; Shirazi, F.; Mirakabadi, A.; Vatanpour, H. Cobra venom cytotoxins; apoptotic or necrotic agents? Toxicon 2015, 108, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Prinholato da Silva, C.; Costa, T.; Paiva, R.; Cintra, A.; Menaldo, D.; Antunes, L.; Sampaio, S. Antitumor potential of the myotoxin BthTX-I from Bothrops jararacussu snake venom: Evaluation of cell cycle alterations and death mechanisms induced in tumor cell lines. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, D.; Lopes, D.; Azevedo, F.; Gimenes, S.; Silva, M.; Achê, D.; Gomes, M.; Vecchi, L.; Goulart, L.; Yoneyama, K.; et al. In vitro antitumor and antiangiogenic effects of Bothropoidin, a metalloproteinase from Bothrops pauloensis snake venom. Int. J. Biol. Macromol. 2017, 97, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Dhananjaya, B.; Sivashankari, P. Snake venom derived molecules in tumor angiogenesis and its application in cancer therapy; an overview. Curr. Top. Med. Chem. 2015, 15, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, F.; Lopes, D.; Cirilo-Gimenes, S.; Achê, D.; Vecchi, L.; Alves, P.; de Oliveira Guimarães, D.; Rodrigues, R.; Goulart, L.; de Melo Rodrigues, V.; et al. Human breast cancer cell death induced by BnSP-6, a Lys-49 PLA homologue from Bothrops pauloensis venom. Int. J. Biol. Macromol. 2016, 82, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Costa, T.; Menaldo, D.; Zoccal, K.; Burin, S.; Aissa, A.; Castro, F.; Faccioli, L.; Greggi-Antunes, L.; Sampaio, S. CR-LAAO, an l-amino acid oxidase from Calloselasma rhodostoma venom, as a potential tool for developing novel immunotherapeutic strategies against cancer. Sci. Rep. 2017, 7, 42673. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, J.; Staren, E.; Appert, H. Adhesion and migration of extracellular matrix-stimulated breast cancer. J. Surg. Res. 2003, 110, 287–294. [Google Scholar] [CrossRef]
- Anderson, L.; Owens, T.; Naylor, M. Structural and mechanical functions of integrins. Biophys. Rev. 2014, 6, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Longmate, W.; DiPersio, C. Beyond adhesion: Emerging roles for integrins in control of the tumor microenvironment. F1000Research 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Kwakwa, K.; Sterling, J. Integrin αvβ3 signaling in tumor-induced bone disease. Cancers (Basel) 2017, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Li, Z. The roles of integrin αvβ6 in cancer. Cancer Lett. 2017, 403, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, R.; Alahari, S. Important role of integrins in the cancer biology. Cancer Metastasis Rev. 2010, 29, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Marelli, U.; Rechenmacher, F.; Sobahi, T.; Mas-Moruno, C.; Kessler, H. Tumor Targeting via Integrin Ligands. Front. Oncol. 2013, 3, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapp, T.; Rechenmacher, F.; Neubauer, S.; Maltsev, O.; Cavalcanti-Adam, E.; Zarka, R.; Reuning, U.; Notni, J.; Wester, H.; Mas-Moruno, C.; et al. A Comprehensive Evaluation of the Activity and Selectivity Profile of Ligands for RGD-binding Integrins. Sci. Rep. 2017, 7, 39805. [Google Scholar] [CrossRef] [PubMed]
- Moura-da-Silva, A.; Theakston, R.; Crampton, J. Evolution of disintegrin cysteine-rich and mammalian matrix-degrading metalloproteinases: Gene duplication and divergence of a common ancestor rather than convergent evolution. J. Mol. Evol. 1996, 43, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.; Marcinkiewicz, C.; Monleón, D.; Esteve, V.; Celda, B.; Juárez, P.; Sanz, L. Snake venom disintegrins: Evolution of structure and function. Toxicon 2005, 45, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Takeya, H.; Iwanaga, S. Snake venom metalloproteinases: Structure, function and relevance to the mammalian ADAM/ADAMTS family proteins. Biochim. Biophys. Acta 2012, 1824, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Weldon, C.; Mackessy, S. Alsophinase, a new P-III metalloproteinase with α-fibrinogenolytic and hemorrhagic activity from the venom of the rear-fanged Puerto Rican Racer Alsophis portoricensis (Serpentes: Dipsadidae). Biochimie 2012, 94, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Peichoto, M.E.; Paes Leme, A.F.; Pauletti, B.A.; Batista, I.C.; Mackessy, S.P.; Acosta, O.; Santoro, M.L. Autolysis at the disintegrin domain of patagonfibrase, a metalloproteinase from Philodryas patagoniensis (Patagonia Green Racer; Dipsadidae) venom. BBA Proteins Proteom. 2010, 1804, 1937–1942. [Google Scholar] [CrossRef] [PubMed]
- Ching, A.T.; Paes Leme, A.F.; Zelanis, A.; Rocha, M.M.; Furtado, M.D.F.; Silva, D.A.; Trugilho, M.R.; da Rocha, S.L.; Perales, J.; Ho, P.L.; et al. Venomics profiling of Thamnodynastes strigatus unveils matrix metalloproteinases and other novel proteins recruited to the toxin arsenal of rear-fanged snakes. J. Proteome Res. 2012, 11, 1152–1162. [Google Scholar] [CrossRef] [PubMed]
- Ching, A.T.; Rocha, M.M.; Paes Leme, A.F.; Pimenta, D.C.; de Fatima, D.F.M.; Serrano, S.M.; Ho, P.L.; Junqueira-de-Azevedo, I.L. Some aspects of the venom proteome of the Colubridae snake Philodryas olfersii revealed from a Duvernoy’s (venom) gland transcriptome. FEBS Lett. 2006, 580, 4417–4422. [Google Scholar] [CrossRef] [PubMed]
- Urra, F.; Pulgar, R.; Gutiérrez, R.; Hodar, C.; Cambiazo, V.; Labra, A. Identification and molecular characterization of five putative toxins from the venom gland of the snake Philodryas chamissonis (Serpentes: Dipsadidae). Toxicon 2015, 108, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Kamiguti, A.; Theakston, R.; Sherman, N.; Fox, J. Mass spectrophotometric evidence for P-III/P-IV metalloproteinases in the venom of the Boomslang (Dispholidus typus). Toxicon 2000, 38, 1613–1620. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, X.; Hu, T.; Zhou, W.; Cui, Q.; Tian, J.; Zheng, Y.; Fan, Q. Discovery of toxin-encoding genes from the false viper Macropisthodon rudis, a rear-fanged snake, by transcriptome analysis of venom gland. Toxicon 2015, 106, 72–78. [Google Scholar] [CrossRef] [PubMed]
- McGivern, J.; Wray, K.; Margres, M.; Couch, M.; Mackessy, S.; Rokyta, D. RNA-seq and high-definition mass spectrometry reveal the complex and divergent venoms of two rear-fanged colubrid snakes. BMC Genom. 2014, 15, 1061. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.; Marcinkiewicz, C. Non-RGD-containing snake venom disintegrins, functional and structural relations. Toxicon 2011, 58, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J. The continuing saga of snake venom disintegrins. Toxicon 2013, 62, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S. ADAM and ADAMTS family proteins and snake venom metalloproteinases: A structural overview. Toxins (Basel) 2016, 8, E155. [Google Scholar] [CrossRef] [PubMed]
- Saviola, A.; Modahl, C.; Mackessy, S. Disintegrins of Crotalus simus tzabcan venom: Isolation, characterization and evaluation of the cytotoxic and anti-adhesion activities of tzabcanin, a new RGD disintegrin. Biochimie 2015, 116, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Saviola, A.; Burns, P.; Mukherjee, A.; Mackessy, S. The disintegrin tzabcanin inhibits adhesion and migration in melanoma and lung cancer cells. Int. J. Biol. Macromol. 2016, 88, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Suntravat, M.; Barret, H.; Jurica, C.; Lucena, S.; Perez, J.; Sánchez, E. Recombinant disintegrin (r-Cam-dis) from Crotalus adamanteus inhibits adhesion of human pancreatic cancer cell lines to laminin-1 and vitronectin. J. Venom Res. 2015, 6, 1–10. [Google Scholar] [PubMed]
- Suntravat, M.; Helmke, T.; Atphaisit, C.; Cuevas, E.; Lucena, S.; Uzcátegui, N.; Sánchez, E.; Rodriguez-Acosta, A. Expression, purification, and analysis of three recombinant ECD disintegrins (r-colombistatins) from P-III class snake venom metalloproteinases affecting platelet aggregation and SK-MEL-28 cell adhesion. Toxicon 2016, 122, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, C.; Casali, B.; Lino, R.; Pachane, B.; Santos, P.; Horwitz, A.; Selistre-de-Araujo, H.; Lamers, M. Inhibition of αvβ3 integrin induces loss of cell directionality of oral squamous carcinoma cells (OSCC). PLoS ONE 2017, 12, e0176226. [Google Scholar] [CrossRef] [PubMed]
- Lucena, S.; Castro, R.; Lundin, C.; Hofstetter, A.; Alaniz, A.; Suntravat, M.; Sánchez, E. Inhibition of pancreatic tumoral cells by snake venom disintegrins. Toxicon 2015, 93, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Lucena, S.; Sanchez, E.; Perez, J. Anti-metastatic activity of the recombinant disintegrin, r-mojastin 1, from the Mohave rattlesnake. Toxicon 2011, 57, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Lucena, S.; Jia, Y.; Soto, J.; Parral, J.; Cantu, E.; Brannon, J.; Lardner, K.; Ramos, C.; Seoane, A.; Sánchez, E. Anti-invasive and anti-adhesive activities of a recombinant disintegrin, r-viridistatin 2, derived from the Prairie rattlesnake (Crotalus viridis viridis). Toxicon 2012, 60, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Jebali, J.; Fakhfekh, E.; Morgen, M.; Srairi-Abid, N.; Majdoub, H.; Gargouri, A.; El Ayeb, M.; Luis, J.; Marrakchi, N.; Sarray, S. Lebecin, a new C-type lectin like protein from Macrovipera lebetina venom with anti-tumor activity against the breast cancer cell line MDA-MB231. Toxicon 2014, 86, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Morjen, M.; Kallech-Ziri, O.; Bazaa, A.; Othman, H.; Mabrouk, K.; Zouari-Kessentini, R.; Sanz, L.; Calvete, J.; Srairi-Abid, N.; El Ayeb, M.; et al. PIVL, a new serine protease inhibitor from Macrovipera lebetina transmediterranea venom, impairs motility of human glioblastoma cells. Matrix Biol. 2013, 32, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Ramos, O.; Kauskot, A.; Cominetti, M.; Bechyne, I.; Salla-Pontes, C.; Chareyre, F.; Manent, J.; Vassy, R.; Giovannini, M.; Legrand, C.; et al. A novel alpha(v)beta (3)-blocking disintegrin containing the RGD motive, DisBa-01, inhibits bFGF-induced angiogenesis and melanoma metastasis. Clin. Exp. Metastasis 2008, 25, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Selistre-de-Araujo, H.; Pontes, C.; Montenegro, C.; Martin, A. Snake venom disintegrins and cell migration. Toxins (Basel) 2010, 2, 2606–2621. [Google Scholar] [CrossRef] [PubMed]
- Sarray, S.; Berthet, V.; Calvete, J.; Secchi, J.; Marvaldi, J.; El-Ayeb, M.; Marrakchi, N.; Luis, J. Lebectin, a novel C-type lectin from Macrovipera lebetina venom, inhibits integrin-mediated adhesion, migration and invasion of human tumour cells. Lab. Investig. 2004, 84, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Sarray, S.; Delamarre, E.; Marvaldi, J.; El Ayeb, M.; Marrakchi, N.; Luis, J. Lebectin and lebecetin, two C-type lectins from snake venom, inhibit alpha5beta1 and alphaV-containing integrins. Matrix Biol. 2007, 26, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Morjen, M.; Honoré, S.; Bazaa, A.; Abdelkafi-Koubaa, Z.; Ellafi, A.; Mabrouk, K.; Kovacic, H.; El Ayeb, M.; Marrakchi, N.; Luis, J. PIVL, a snake venom Kunitz-type serine protease inhibitor, inhibits in vitro and in vivo angiogenesis. Microvasc. Res. 2014, 95, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Ansieau, S.; Bastid, J.; Doreau, A.; Morel, A.; Bouchet, B.; Thomas, C.; Fauvet, F.; Puisieux, I.; Doglioni, C.; Piccinin, S.; et al. Induction of EMT by twist proteins as a collateral effect of tumor-promoting inactivation of premature senescence. Cancer Cell 2008, 14, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.; Durrans, A.; Lee, S.; Sheng, J.; Li, F.; Wong, S.; Choi, H.; El Rayes, T.; Ryu, S.; Troeger, J.; et al. Epithelial-to-mesenchymal transition is not required for lung metastasis but contributes to chemoresistance. Nature 2015, 527, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Kajiyama, H.; Shibata, K.; Terauchi, M.; Yamashita, M.; Ino, K.; Nawa, A.; Kikkawa, F. Chemoresistance to paclitaxel induces epithelial-mesenchymal transition and enhances metastatic potential for epithelial ovarian carcinoma cells. Int. J. Oncol. 2007, 31, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Shibue, T.; Weinberg, R. EMT, CSCs, and drug resistance: The mechanistic link and clinical implications. Nat. Rev. Clin. Oncol. 2017, 14, 611–629. [Google Scholar] [CrossRef] [PubMed]
- Mulholland, D.; Kobayashi, N.; Ruscetti, M.; Zhi, A.; Tran, L.; Huang, J.; Gleave, M.; Wu, H. Pten loss and RAS/MAPK activation cooperate to promote EMT and metastasis initiated from prostate cancer stem/progenitor cells. Cancer Res. 2012, 72, 1878–1889. [Google Scholar] [CrossRef] [PubMed]
- Bo, H.; Zhang, S.; Gao, L.; Chen, Y.; Zhang, J.; Chang, X.; Zhu, M. Upregulation of Wnt5a promotes epithelial-to-mesenchymal transition and metastasis of pancreatic cancer cells. BMC Cancer 2013, 13, 496. [Google Scholar] [CrossRef] [PubMed]
- Foroni, C.; Broggini, M.; Generali, D.; Damia, G. Epithelial-mesenchymal transition and breast cancer: Role, molecular mechanisms and clinical impact. Cancer Treat. Rev. 2012, 38, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Nieto, M.; Cano, A. The epithelial-mesenchymal transition under control: Global programs to regulate epithelial plasticity. Semin. Cancer Biol. 2012, 22, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, R.; Huang, C.; Chang, D.; Yu, C. Cardiotoxin III from the Taiwan cobra (Naja naja atra). Determination of structure in solution and comparison with short neurotoxins. J. Mol. Biol. 1994, 235, 1291–1301. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.; Chang, S.; Lin, K.; Chiu, C.; Chang, L.; Lin, S. Taiwan cobra cardiotoxin III inhibits Src kinase leading to apoptosis and cell cycle arrest of oral squamous cell carcinoma Ca9-22 cells. Toxicon 2010, 56, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Su, J.; Chien, C.; Chuang, P.; Chang, L.; Lin, S. Down-regulation of the JAK2/PI3K-mediated signaling activation is involved in Taiwan cobra cardiotoxin III-induced apoptosis of human breast MDA-MB-231 cancer cells. Toxicon 2010, 55, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Lin, S.; Chang, L. Involvement of mitochondrial alteration and reactive oxygen species generation in Taiwan cobra cardiotoxin-induced apoptotic death of human neuroblastoma SK-N-SH cells. Toxicon 2008, 52, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.; Liang, S.; Han, L.; Chou, H.; Chou, C.; Lin, S.; Chiu, C. Cardiotoxin III inhibits proliferation and migration of oral cancer cells through MAPK and MMP signaling. Sci. World J. 2013, 2013, 650946. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Chien, C.; Hsieh, C.; Tsai, P.; Chang, L.; Lin, S. Antimetastatic potential of cardiotoxin III involves inactivation of PI3K/Akt and p38 MAPK signaling pathways in human breast cancer MDA-MB-231 cells. Life Sci. 2012, 90, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Chiang, S.; Cabrera, R.; Segall, J. Tumor cell intravasation. Am. J. Physiol. Cell Physiol. 2016, 311, C1–C14. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.; Hsu, S.; Xia, W.; Cao, X.; Shih, J.; Wei, Y.; Abbruzzese, J.; Hortobagyi, G.; Hung, M. Epidermal growth factor receptor cooperates with signal transducer and activator of transcription 3 to induce epithelial-mesenchymal transition in cancer cells via up-regulation of TWIST gene expression. Cancer Res. 2007, 67, 9066–9076. [Google Scholar] [CrossRef] [PubMed]
- Balanis, N.; Carlin, C. Stress-induced EGF receptor signaling through STAT3 and tumor progression in triple-negative breast cancer. Mol. Cell. Endocrinol. 2017, 451, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kong, J.; Chang, H.; Kim, H.; Kim, A. EGF induces epithelial-mesenchymal transition through phospho-Smad2/3-Snail signaling pathway in breast cancer cells. Oncotarget 2016, 7, 85021–85032. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.; Hsieh, C.; Chiu, C.; Wang, C.; Chang, L.; Lin, S. Cardiotoxin III suppresses MDA-MB-231 cell metastasis through the inhibition of EGF/EGFR-mediated signaling pathway. Toxicon 2012, 60, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.; Fu, Y.; Chang, L.; Lin, S. Taiwan cobra cardiotoxin III suppresses EGF/EGFR-mediated epithelial-to-mesenchymal transition and invasion of human breast cancer MDA-MB-231 cells. Toxicon 2016, 111, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.; Chu, C.; Chiu, C.; Chang, L.; Lin, S. Cardiotoxin III suppresses hepatocyte growth factor-stimulated migration and invasion of MDA-MB-231 cells. Cell Biochem. Funct. 2014, 32, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.; Fu, Y.; Chang, L.; Lin, S. Cardiotoxin III Inhibits Hepatocyte Growth Factor-Induced Epithelial-Mesenchymal Transition and Suppresses Invasion of MDA-MB-231 Cells. J. Biochem. Mol. Toxicol. 2016, 30, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.; Sloane, B. Cysteine cathepsins: Multifunctional enzymes in cancer. Nat. Rev. Cancer 2006, 6, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Fonović, M.; Turk, B. Cysteine cathepsins and extracellular matrix degradation. Biochim. Biophys. Acta 2014, 1840, 2560–2570. [Google Scholar] [CrossRef] [PubMed]
- Löser, R.; Pietzsch, J. Cysteine cathepsins: Their role in tumor progression and recent trends in the development of imaging probes. Front. Chem. 2015, 3, 37. [Google Scholar] [CrossRef] [PubMed]
- Sobotič, B.; Vizovišek, M.; Vidmar, R.; Van Damme, P.; Gocheva, V.; Joyce, J.; Gevaert, K.; Turk, V.; Turk, B.; Fonović, M. Proteomic Identification of Cysteine Cathepsin Substrates Shed from the Surface of Cancer Cells. Mol. Cell. Proteom. 2015, 14, 2213–2228. [Google Scholar] [CrossRef] [PubMed]
- Gocheva, V.; Zeng, W.; Ke, D.; Klimstra, D.; Reinheckel, T.; Peters, C.; Hanahan, D.; Joyce, J. Distinct roles for cysteine cathepsin genes in multistage tumorigenesis. Genes Dev. 2006, 20, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Friedl, P.; Alexander, S. Cancer invasion and the microenvironment: Plasticity and reciprocity. Cell Biochem. Funct. 2011, 147, 992–1009. [Google Scholar] [CrossRef] [PubMed]
- Jedeszko, C.; Sloane, B. Cysteine cathepsins in human cancer. Biol. Chem. 2004, 385, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, A.; Bano, B. Journey of cystatins from being mere thiol protease inhibitors to at heart of many pathological conditions. Int. J. Biol. Macromol. 2017, 102, 674–693. [Google Scholar] [CrossRef] [PubMed]
- Brillard-Bourdet, M.; Nguyên, V.; Ferrer-di Martino, M.; Gauthier, F.; Moreau, T. Purification and characterization of a new cystatin inhibitor from Taiwan cobra (Naja naja atra) venom. Biochem. J. 1998, 331, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Xie, Q.; Wang, X.; Li, X.; Chen, Y.; Lin, X.; Lin, J. Inhibition of invasion and metastasis of MHCC97H cells by expression of snake venom cystatin through reduction of proteinases activity and epithelial-mesenchymal transition. Arch. Pharm. Res. 2011, 34, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Mayor, R.; Etienne-Manneville, S. The front and rear of collective cell migration. Nat. Rev. Mol. Cell Biol. 2016, 17, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Zegers, M.; Friedl, P. Rho GTPases in collective cell migration. Small GTPases 2014, 5, e28997. [Google Scholar] [CrossRef] [PubMed]
- Sartim, M.; Sampaio, S. Snake venom galactoside-binding lectins: A structural and functional overview. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathan, J.; Mondal, S.; Sarkar, A.; Chakrabarty, D. Daboialectin, a C-type lectin from Russell’s viper venom induces cytoskeletal damage and apoptosis in human lung cancer cells in vitro. Toxicon 2017, 127, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.; Marangoni, S.; Oliveira, B.; Novello, J. Isolation and characterization of a new lectin from the venom of the snake Bothrops jararacussu. IUBMB Life 1998, 44, 933–938. [Google Scholar] [CrossRef]
- Nolte, S.; de Castro Damasio, D.; Baréa, A.; Gomes, J.; Magalhães, A.; Mello Zischler, L.; Stuelp-Campelo, P.; Elífio-Esposito, S.; Roque-Barreira, M.; Reis, C.; et al. BJcuL, a lectin purified from Bothrops jararacussu venom, induces apoptosis in human gastric carcinoma cells accompanied by inhibition of cell adhesion and actin cytoskeleton disassembly. Toxicon 2012, 59, 81–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Carvalho, D.; Schmitmeier, S.; Novello, J.; Markland, F. Effect of BJcuL (a lectin from the venom of the snake Bothrops jararacussu) on adhesion and growth of tumor and endothelial cells. Toxicon 2001, 39, 1471–1476. [Google Scholar] [CrossRef]
- Peichoto, M.E.; Teibler, P.; Mackessy, S.P.; Leiva, L.; Acosta, O.; Goncalves, L.R.; Tanaka-Azevedo, A.M.; Santoro, M.L. Purification and characterization of patagonfibrase, a metalloproteinase showing alpha-fibrinogenolytic and hemorrhagic activities, from Philodryas patagoniensis snake venom. BBA Gen. Subj. 2007, 1770, 810–819. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.N.; Timoniuk, A.; Maruñak, S.; Teibler, P.; Acosta, O.; Peichoto, M.E. Biochemical and biological analysis of Philodryas baroni (Baron’s Green Racer; Dipsadidae) venom: Relevance to the findings of human risk assessment. Hum. Exp. Toxicol. 2014, 33, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Heyborne, W.H.; Mackessy, S.P. Identification and characterization of a taxon-specific three-finger toxin from the venom of the Green Vinesnake (Oxybelis fulgidus; family Colubridae). Biochimie 2013, 95, 1923–1932. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, T.; Saha, P.; Sarkar, A.; Gomes, A. Evaluation of cytotoxicity of a purified venom protein from Naja kaouthia (NKCT1) using gold nanoparticles for targeted delivery to cancer cell. Chem. Biol. Interact. 2017, 261, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Badr, G.; Sayed, D.; Maximous, D.; Mohamed, A.; Gul, M. Increased susceptibility to apoptosis and growth arrest of human breast cancer cells treated by a snake venom-loaded silica nanoparticles. Cell. Physiol. Biochem. 2014, 34, 1640–1651. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Toxin Name | Snake Species | Adhesive Motif | Integrin Target | ECM Ligand | Effect | Ref. |
|---|---|---|---|---|---|---|
| r-Cam-dis recombinant disintegrin | Crotalus adamanteus | RGD | αvβ3 | laminin-1 | Inhibition of adhesion in pancreatic cancer cells | [41] |
| r-Colombistatins recombinant disintegrin-like domains from Class-III SVMP | Bothrops colombiensis | ECD | n.d. | collagen I | Inhibition of adhesion in SK-Mel-28 melanoma cells | [42] |
| DisBa-01, recombinant disintegrin | Bothrops alternatus | RGD | αvβ3 | fibronectin | Loss of cell directionality of migrating oral squamous carcinoma cells | [43] |
| r-mojastn-1, recombinant disintegrin | Crotalus scutulatus scutulatus | RGD | αvβ3, α3, and β1, | fibronectin and vitronectin | Inhibition of adhesion and migration of BXPC-3 pancreatic cancer cell line | [44,45] |
| r-viridistatin-2, recombinant disintigrin | Crotalus viridis viridis | RGD | αvβ3 | fibronectin and vitronectin | Inhibition of adhesion, migration and invasion of several cancer cell lines | [44,46] |
| Lebecin, C-type lectin-like protein | Macrovipera lebetina | - | αvβ3 | fibronectin and fibrinogen | Inhibition of adhesion and migration of MDA-MB-231 breast cancer cells | [47] |
| PIVL, Kunitz-type serin protease inhibitor | Macrovipera lebetina transmediterranea | RGN | αvβ3 | fibronectin and fibrinogen | Inhibition of adhesion, migration and invasion of human glioblastoma U87 cells | [48] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urra, F.A.; Araya-Maturana, R. Targeting Metastasis with Snake Toxins: Molecular Mechanisms. Toxins 2017, 9, 390. https://doi.org/10.3390/toxins9120390
Urra FA, Araya-Maturana R. Targeting Metastasis with Snake Toxins: Molecular Mechanisms. Toxins. 2017; 9(12):390. https://doi.org/10.3390/toxins9120390
Chicago/Turabian StyleUrra, Félix A., and Ramiro Araya-Maturana. 2017. "Targeting Metastasis with Snake Toxins: Molecular Mechanisms" Toxins 9, no. 12: 390. https://doi.org/10.3390/toxins9120390
APA StyleUrra, F. A., & Araya-Maturana, R. (2017). Targeting Metastasis with Snake Toxins: Molecular Mechanisms. Toxins, 9(12), 390. https://doi.org/10.3390/toxins9120390