
Molecular Modeling and Simulation Tools in the Development of Peptide-Based Biosensors for Mycotoxin Detection: Example of Ochratoxin



Abstract
:1. Introduction
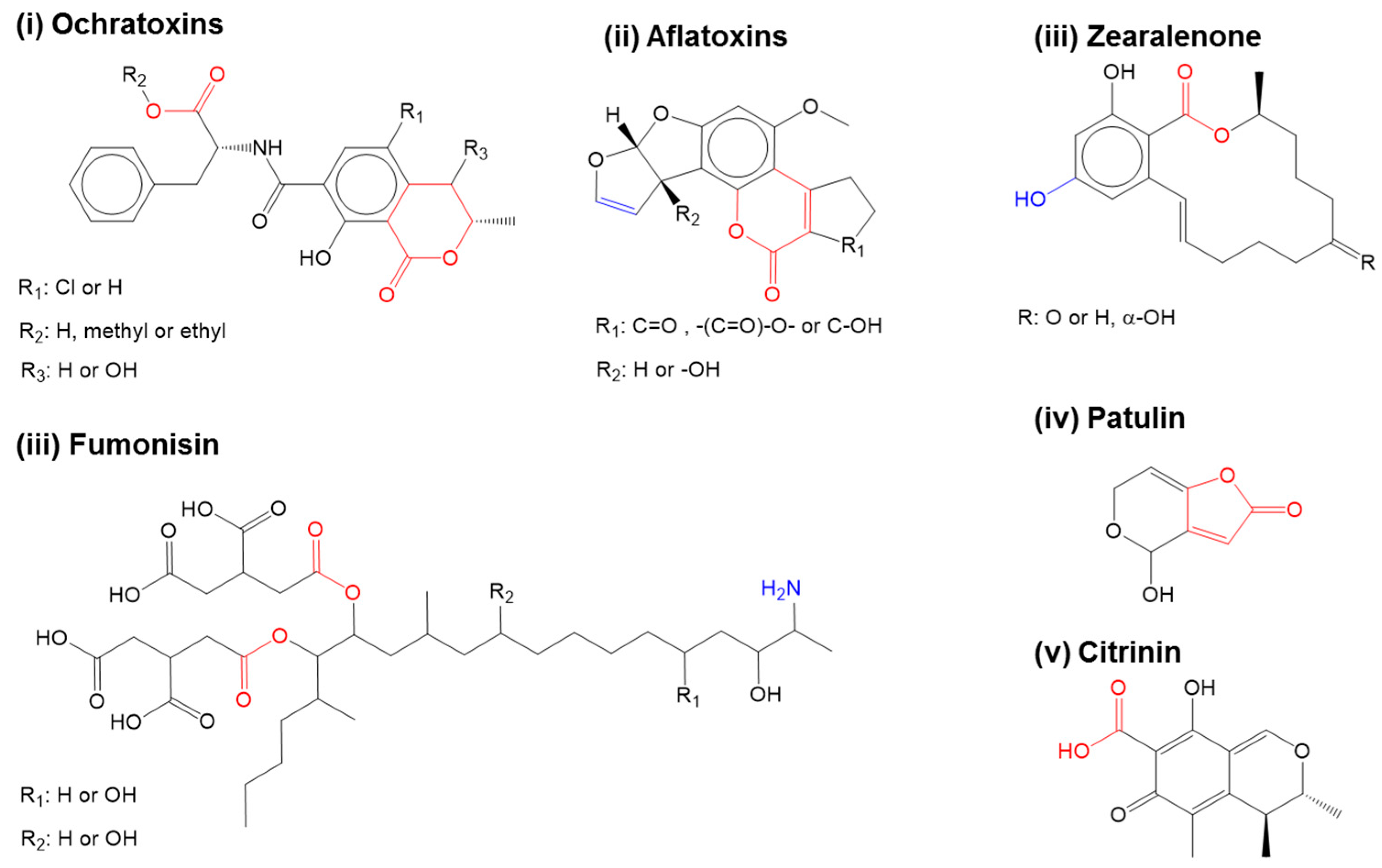
2. Challenges in the Fabrication of Sensing Element for Mycotoxin Monitoring
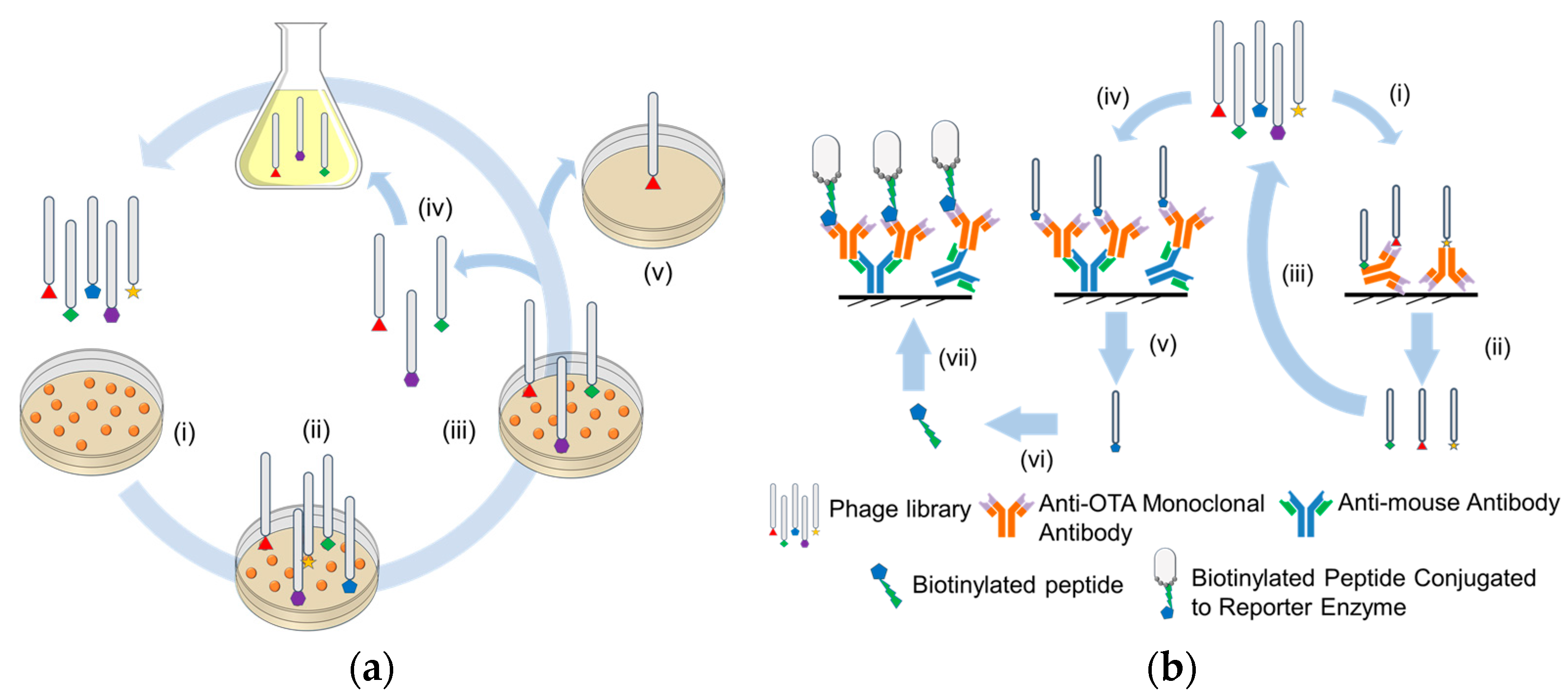
2.1. Screening of Peptide Receptors for Mycotoxin Specificity in Monitoring
2.2. Sub-Molecular Processes Influencing the Performance of Peptide Bioceceptors
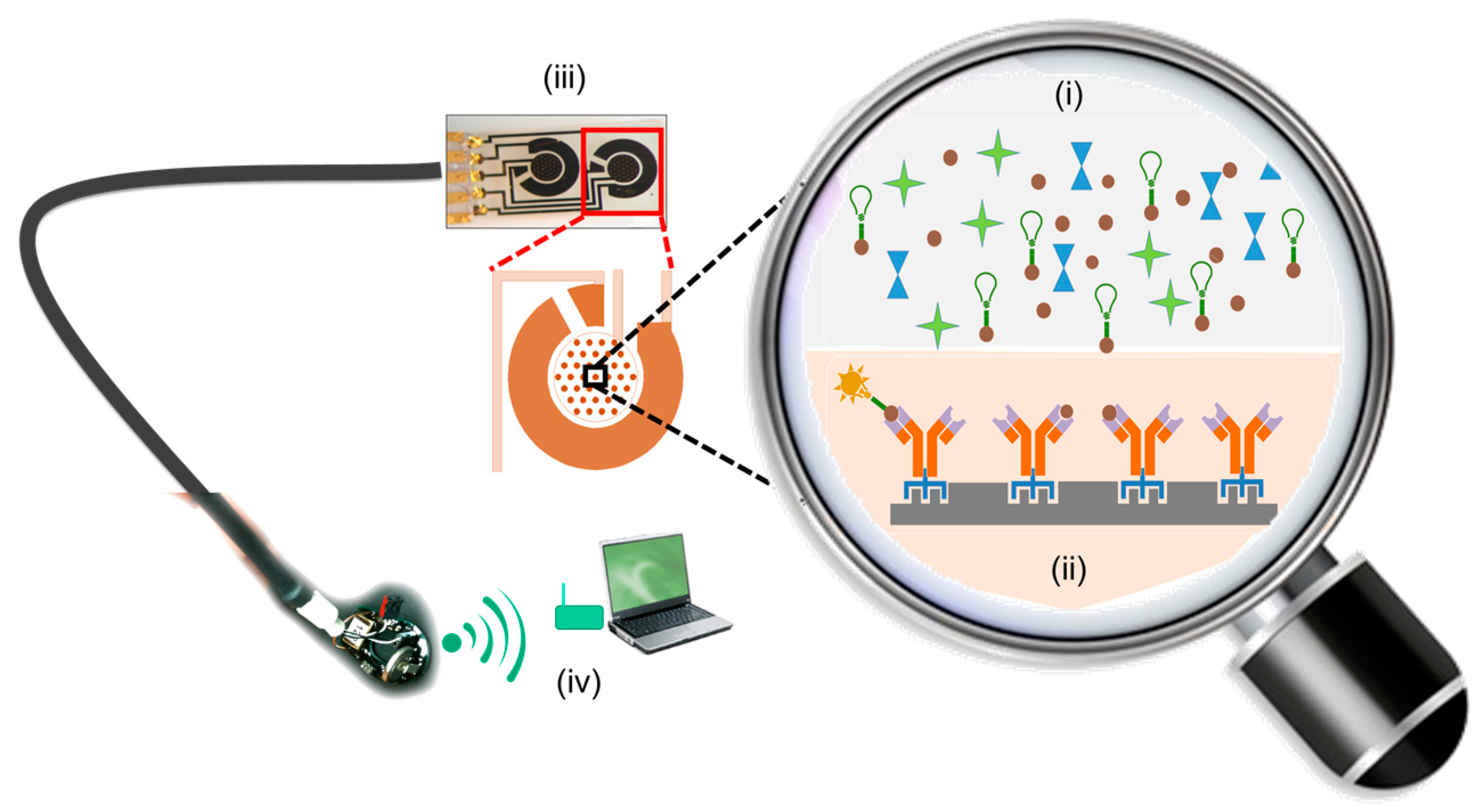
2.2.1. Non-Specific Peptide-Surface Interactions Influence Bioreceptor Performance
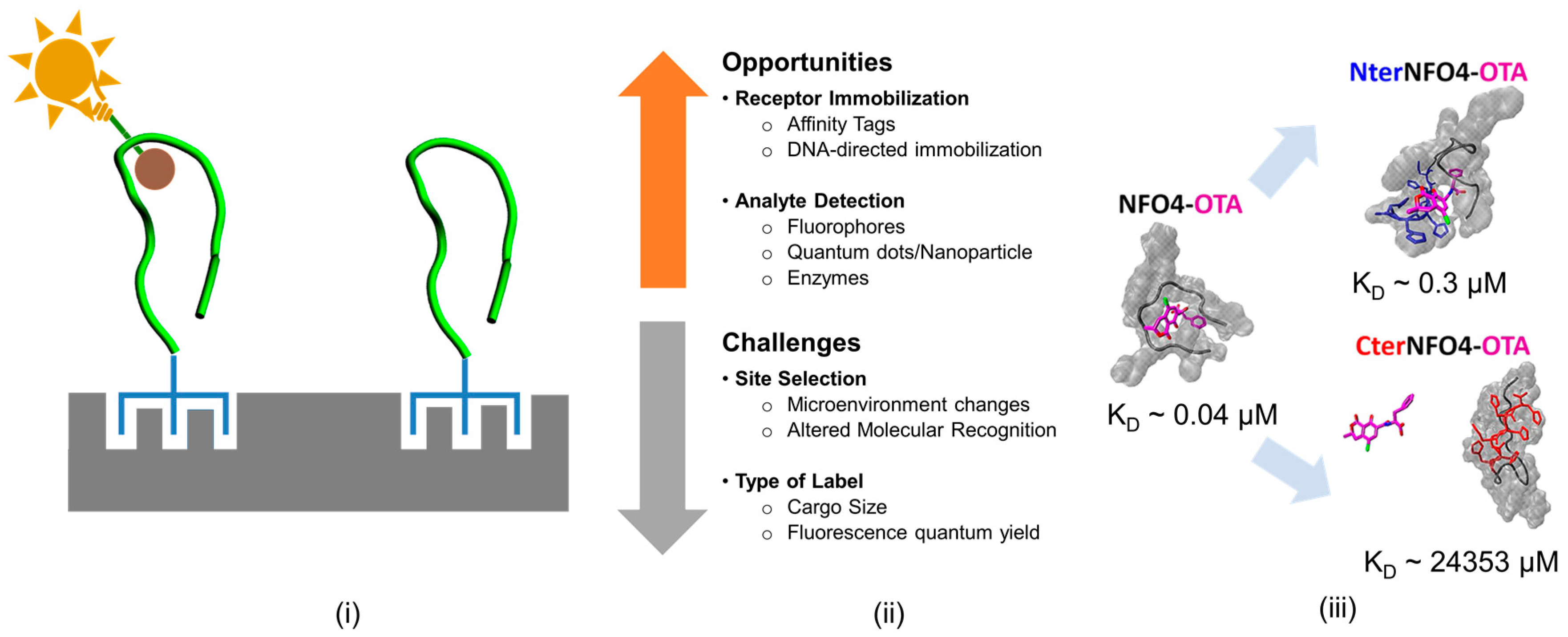
2.2.2. Site-Specificity and Type of Chemical/Biological Modification Influence Sensing Element Performance
3. Molecular Modeling Approaches towards Design and Fabrication of Peptide Bioreceptors with Optimal Hapten Binding
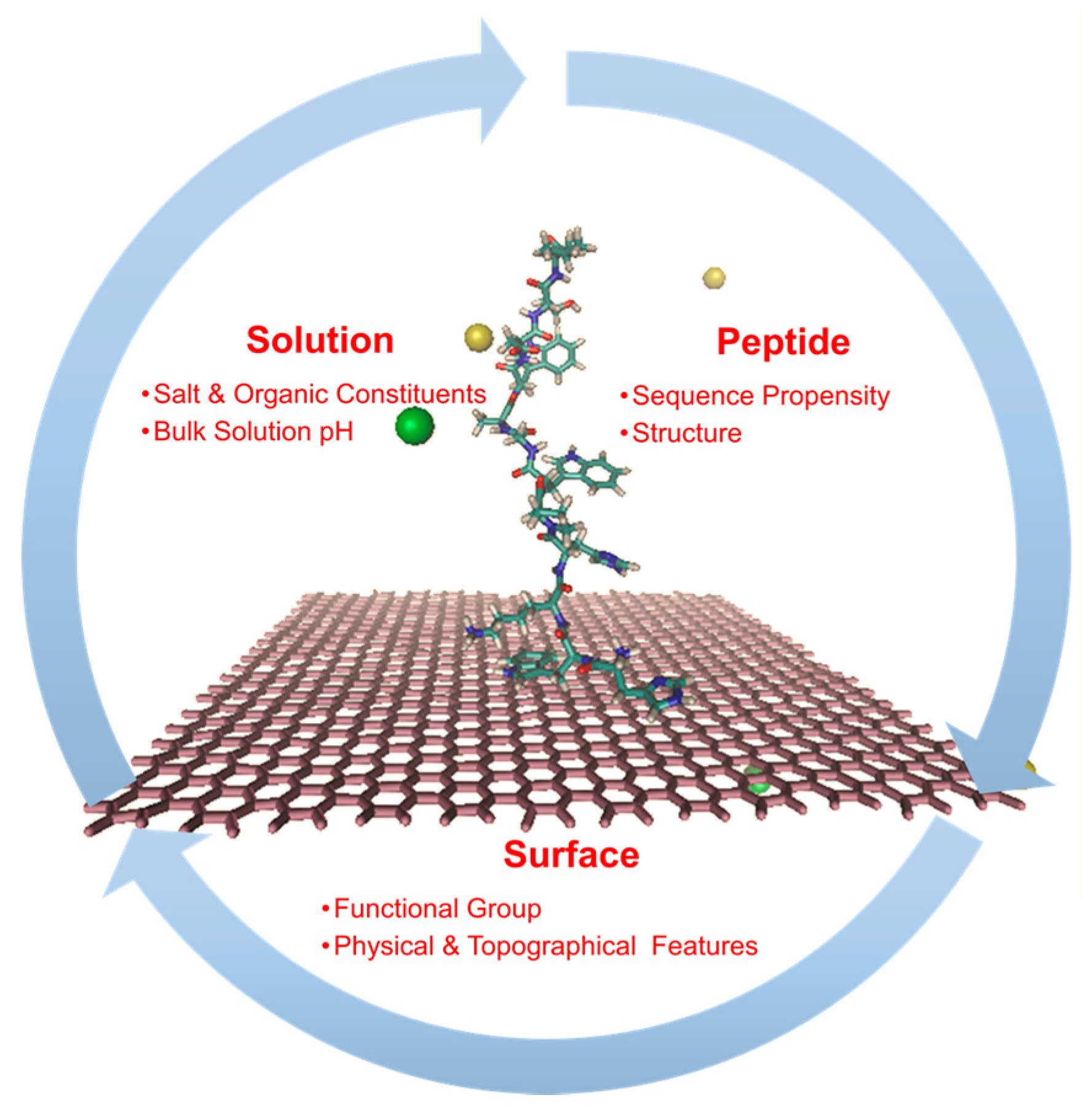
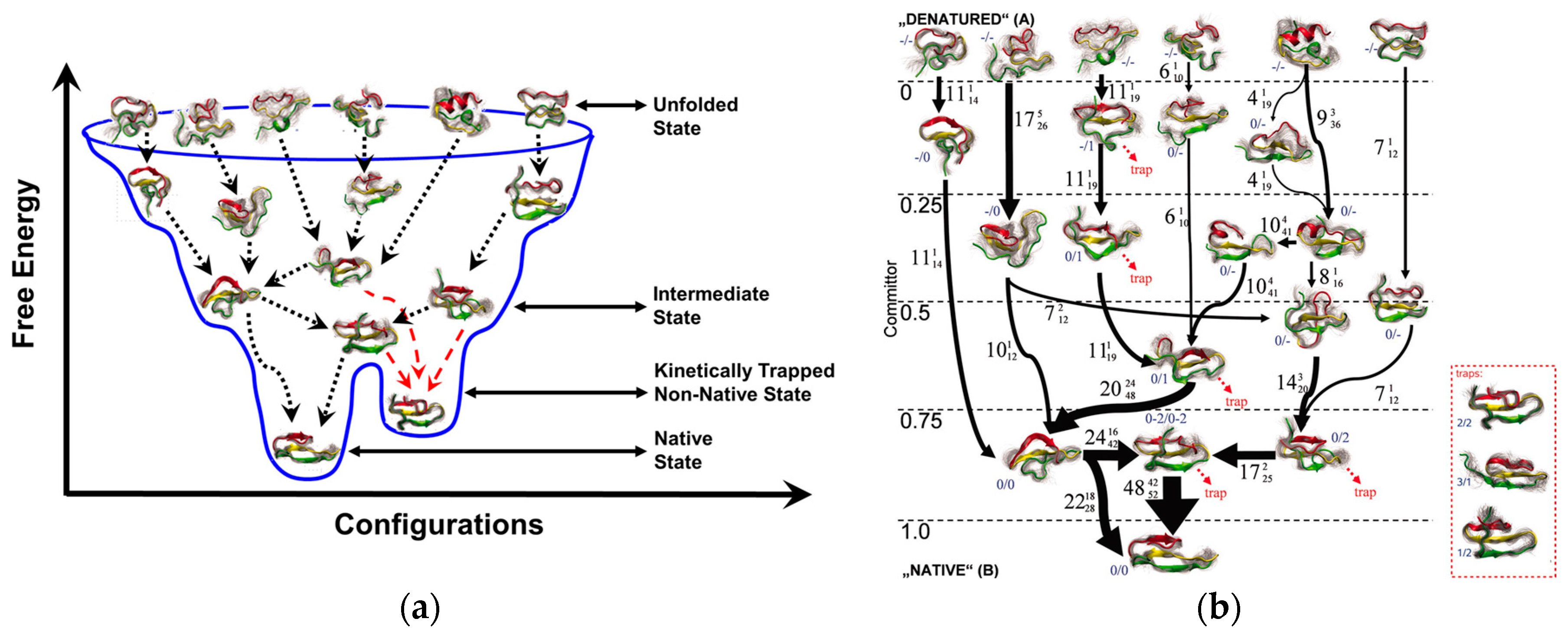
3.1. Basic Considerations in the Application of Molecular Dynamics Simulations to Peptide Bioreceptors
3.2. Evaluating the In-Solution Mycotoxin Recognition Properties of Peptide Bioreceptors
3.3. Evaluating the Effect of Site-Specific Modifications on Recognition Properties of Peptide Bioreceptors
3.4. Evaluating the Effect of Interfaces with Solid Substrates on the Recognition Properties of Peptide Bioreceptors
4. Conclusions and Outlook
Acknowledgments
Conflicts of Interest
References
- Jacxsens, L.; Uyttendaele, M.; Luning, P.; Allende, A. Food safety management and risk assessment in the fresh produce supply chain. IOP Conf. Ser. Mater. Sci. Eng. 2017, 193, 012020. [Google Scholar] [CrossRef]
- Misihairabgwi, J.M.; Ezekiel, C.N.; Sulyok, M.; Shephard, G.S.; Krska, R. Mycotoxin contamination of foods in southern africa: A 10-year review (2007–2016). Crit. Rev. Food Sci. Nutr. 2017. [Google Scholar] [CrossRef] [PubMed]
- Sheahan, M.; Barrett, C.B. Review: Food loss and waste in sub-saharan africa. Food Policy 2017, 70, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pinotti, L.; Ottoboni, M.; Giromini, C.; Dell’Orto, V.; Cheli, F. Mycotoxin contamination in the eu feed supply chain: A focus on cereal byproducts. Toxins 2016, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, I.; Audenaert, K.; De Gelder, L. Biodegradation of mycotoxins: Tales from known and unexplored worlds. Front. Microbiol. 2016, 7, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A: 50 Years of Research. Toxins 2016, 8, 191. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; Bingle, L.E. Comparative ochratoxin toxicity: A review of the available data. Toxins 2015, 7, 4253–4282. [Google Scholar] [CrossRef] [PubMed]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Novotna, E. Ochratoxin A: Developmental and reproductive toxicity-an overview. Birth Defects Res. Part B 2013, 98, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Koszegi, T.; Poor, M. Ochratoxin A: Molecular interactions, mechanisms of toxicity and prevention at the molecular level. Toxins 2016, 8, 111. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, B.D.; Srivastava, S.; Ali, M.A.; Singh, C. Nanomaterial-based biosensors for food toxin detection. Appl. Biochem. Biotechnol. 2014, 174, 880–896. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Sun, Y.M.; Beier, R.C.; Lei, H.T.; Gee, S.; Hammock, B.D.; Wang, H.; Wang, Z.H.; Sun, X.L.; Shen, Y.D.; et al. Immunochemical techniques for multianalyte analysis of chemical residues in food and the environment: A review. Trac-Trend Anal. Chem. 2017, 88, 25–40. [Google Scholar] [CrossRef]
- Wang, Z.H.; Beier, R.C.; Shen, J.Z. Immunoassays for the detection of macrocyclic lactones in food matrices—A review. Trac-Trend Anal. Chem. 2017, 92, 42–61. [Google Scholar] [CrossRef]
- Bazin, I.; Tria, S.A.; Hayat, A.; Marty, J.L. New biorecognition molecules in biosensors for the detection of toxins. Biosens. Bioelectron. 2017, 87, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martínez, L.M.; Marquez-Ipiña, A.R.; López-Pacheco, F.; Pérez-Chavarría, R.; González-Vázquez, J.C.; González-González, E.; Trujillo-de Santiago, G.; Ponce-Ponce de León, C.A.; Zhang, Y.S.; Dokmeci, M.R.; et al. Antibody derived peptides for detection of ebola virus glycoprotein. PLoS ONE 2015, 10, e0135859. [Google Scholar] [CrossRef] [PubMed]
- Pai, N.P.; Vadnais, C.; Denkinger, C.; Engel, N.; Pai, M. Point-of-care testing for infectious diseases: Diversity, complexity, and barriers in low- and middle-income countries. PLoS Med. 2012, 9, e1001306. [Google Scholar] [CrossRef] [PubMed]
- Yue, N.; Kuang, H.; Sun, L.; Wu, L.H.; Xu, C.L. An empirical analysis of the impact of eu’s new food safety standards on china’s tea export. Int. J. Food Sci. Technol. 2010, 45, 745–750. [Google Scholar] [CrossRef]
- Novak, M.T.; Kotanen, C.N.; Carrara, S.; Guiseppi-Elie, A.; Moussy, F.G. Diagnostic tools and technologies for infectious and non-communicable diseases in low-and-middle-income countries. Health Technol. 2013, 3, 271–281. [Google Scholar] [CrossRef]
- Kotanen, C.N.; Guiseppi-Elie, A. Monitoring systems and quantitative measurement of biomolecules for the management of trauma. Biomed. Microdevices 2013, 15, 561–577. [Google Scholar] [CrossRef] [PubMed]
- Karunwi, O.; Wilson, A.N.; Kotanen, C.; Guiseppi-Elie, A. Engineering the abio-bio interface to enable more than moore in functional bioelectronics. J. Electrochem. Soc. 2013, 160, B60–B65. [Google Scholar] [CrossRef]
- Kotanen, C.N.; Moussy, F.G.; Carrara, S.; Guiseppi-Elie, A. Implantable enzyme amperometric biosensors. Biosens. Bioelectron. 2012, 35, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Strycharz-Glaven, S.M.; Snider, R.M.; Guiseppi-Elie, A.; Tender, L.M. On the electrical conductivity of microbial nanowires and biofilms. Energy Environ. Sci. 2011, 4, 4366–4379. [Google Scholar] [CrossRef]
- Yang, L.; Guiseppi-Wilson, A.; Guiseppi-Elie, A. Design considerations in the use of interdigitated microsensor electrode arrays (imes) for impedimetric characterization of biomimetic hydrogels. Biomed. Microdevices 2011, 13, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.; Brahim, S.; Ishihara, K.; Guiseppi-Elie, A. Molecularly engineered p(hema)-based hydrogels for implant biochip biocompatibility. Biomaterials 2005, 26, 4767–4778. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.; Smith, S.; Windle, B.; Guiseppi-Elie, A. Impact of surface chemistry and blocking strategies on DNA microarrays. Nucleic Acids Res. 2003, 31, e87. [Google Scholar] [CrossRef] [PubMed]
- Anthony, G.-E.; Chenghong, L.; Ray, H.B. Direct electron transfer of glucose oxidase on carbon nanotubes. Nanotechnology 2002, 13, 559–564. [Google Scholar]
- Brahim, S.; Narinesingh, D.; Guiseppi-Elie, A. Bio-smart hydrogels: Co-joined molecular recognition and signal transduction in biosensor fabrication and drug delivery. Biosens. Bioelectron. 2002, 17, 973–981. [Google Scholar] [CrossRef]
- Thyparambil, A.A.; Bazin, I.; Guiseppi-Elie, A. Evaluation of ochratoxin recognition by peptides using explicit solvent molecular dynamics. Toxins 2017, 9, 164. [Google Scholar] [CrossRef] [PubMed]
- Thyparambil, A.A.; Abramyan, T.M.; Bazin, I.; Guiseppi-Elie, A. Site of tagging influences the ochratoxin recognition by peptide nfo4: A molecular dynamics study. J. Chem. Inf. Model. 2017, 57, 2035–2044. [Google Scholar] [CrossRef] [PubMed]
- Demey, H.; Tria, S.A.; Soleri, R.; Guiseppi-Elie, A.; Bazin, I. Sorption of his-tagged Protein G and Protein G onto chitosan/divalent metal ion sorbent used for detection of microcystin-LR. Environ. Sci. Pollut. Res. 2017, 24, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Tria, S.A.; Lopez-Ferber, D.; Gonzalez, C.; Bazin, I.; Guiseppi-Elie, A. Microfabricated biosensor for the simultaneous amperometric and luminescence detection and monitoring of ochratoxin A. Biosens. Bioelectron. 2016, 79, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Soleri, R.; Demey, H.; Tria, S.A.; Guiseppi-Elie, A.; Hassine, A.I.; Gonzalez, C.; Bazin, I. Peptide conjugated chitosan foam as a novel approach for capture-purification and rapid detection of hapten—Example of ochratoxin A. Biosens. Bioelectron. 2015, 67, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, Z.; Zhang, Q.; Li, P. Mycotoxin determination in foods using advanced sensors based on antibodies or aptamers. Toxins 2016, 8, 239. [Google Scholar] [CrossRef] [PubMed]
- McKeague, M.; Velu, R.; De Girolamo, A.; Valenzano, S.; Pascale, M.; Smith, M.; DeRosa, M.C. Comparison of in-solution biorecognition properties of aptamers against ochratoxin A. Toxins 2016, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.H. Recent advances for the detection of ochratoxin A. Toxins 2015, 7, 5276–5300. [Google Scholar] [CrossRef] [PubMed]
- Baggiani, C.; Giovannoli, C.; Anfossi, L. Man-made synthetic receptors for capture and analysis of ochratoxin A. Toxins 2015, 7, 4083–4098. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, A.; Arya, S.K.; Vasudev, A.; Bhansali, S. Recent advances in detection of ochratoxin-A. Open J. Appl. Biosens. 2013, 2, 1–11. [Google Scholar] [CrossRef]
- Piro, B.; Shi, S.; Reisberg, S.; Noel, V.; Anquetin, G. Comparison of electrochemical immunosensors and aptasensors for detection of small organic molecules in environment, food safety, clinical and public security. Biosensors 2016, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Heurich, M.; Altintas, Z.; Tothill, I.E. Computational design of peptide ligands for ochratoxin A. Toxins 2013, 5, 1202–1218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Sun, M.; Kang, Y.; Xie, H.; Wang, X.; Song, H.; Li, X.; Fang, W. Identification of a high-affinity monoclonal antibody against ochratoxin A and its application in enzyme-linked immunosorbent assay. Toxicon 2015, 106, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Siantar, D.P.; Halverson, C.A.; Kirmiz, C.; Peterson, G.F.; Hill, N.R.; Dugar, S.M. Ochratoxin A in wine: Survey by antibody- and polymeric-based spe columns using hplc/fluorescence detection. Am. J. Enol. Vitic. 2003, 54, 170–177. [Google Scholar]
- Rhouati, A.; Yang, C.; Hayat, A.; Marty, J.-L. Aptamers: A promising tool for ochratoxin A detection in food analysis. Toxins 2013, 5, 1988. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Aguado, J.A.; Penner, G. Determination of ochratoxin A with a DNA aptamer. J. Agric. Food Chem. 2008, 56, 10456–10461. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Chen, C.; Huang, X.; Chen, X.; Wang, L.; Xiong, Y. Phage-free peptide elisa for ochratoxin A detection based on biotinylated mimotope as a competing antigen. Talanta 2016, 146, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Giovannoli, C.; Passini, C.; Volpi, G.; Di Nardo, F.; Anfossi, L.; Baggiani, C. Peptide-based affinity media for solid-phase extraction of ochratoxin A from wine samples: Effect of the solid support on binding properties. Talanta 2015, 144, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Giraudi, G.; Ferrero, V.E.; Anfossi, L.; Baggiani, C.; Giovannoli, C.; Tozzi, C. Solid-phase extraction of ochratoxin A from wine based on a binding hexapeptide prepared by combinatorial synthesis. J. Chromatogr. A 2007, 1175, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Bazin, I.; Andreotti, N.; Hassine, A.I.; De Waard, M.; Sabatier, J.M.; Gonzalez, C. Peptide binding to ochratoxin A mycotoxin: A new approach in conception of biosensors. Biosens. Bioelectron. 2013, 40, 240–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazin, I.; Faucet-Marquis, V.; Monje, M.C.; El Khoury, M.; Marty, J.L.; Pfohl-Leszkowicz, A. Impact of ph on the stability and the cross-reactivity of ochratoxin A and citrinin. Toxins 2013, 5, 2324–2340. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Yu, Z.; He, Q.; Xu, Y. An immunoassay for ochratoxin A without the mycotoxin. Food Control 2007, 18, 872–877. [Google Scholar] [CrossRef]
- He, Z.Y.; He, Q.H.; Xu, Y.; Li, Y.P.; Liu, X.; Chen, B.; Lei, D.; Sun, C.H. Ochratoxin a mimotope from second-generation peptide library and its application in immunoassay. Anal. Chem. 2013, 85, 10304–10311. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Li, X.; Lam, K.S. Combinatorial chemistry in drug discovery. Curr. Opin. Chem. Biol. 2017, 38, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Macarron, R.; Banks, M.N.; Bojanic, D.; Burns, D.J.; Cirovic, D.A.; Garyantes, T.; Green, D.V.S.; Hertzberg, R.P.; Janzen, W.P.; Paslay, J.W.; et al. Impact of high-throughput screening in biomedical research. Nat. Rev. Drug Discov. 2011, 10, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Scior, T.; Bender, A.; Tresadern, G.; Medina-Franco, J.L.; Martinez-Mayorga, K.; Langer, T.; Cuanalo-Contreras, K.; Agrafiotis, D.K. Recognizing pitfalls in virtual screening: A critical review. J. Chem. Inf. Model. 2012, 52, 867–881. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.R. Pathways to structure–property relationships of peptide–materials interfaces: Challenges in predicting molecular structures. Acc. Chem. Res. 2017, 50, 1617–1624. [Google Scholar] [CrossRef] [PubMed]
- Schwaminger, S.; Blank-Shim, S.A.; Borkowska-Panek, M.; Anand, P.; Fraga-García, P.; Fink, K.; Wenzel, W.; Berensmeier, S. Experimental characterization and simulation of amino acid and peptide interactions with inorganic materials. Eng. Life Sci. 2017. [Google Scholar] [CrossRef]
- Schroeder, M.M.; Wang, Q.; Badieyan, S.; Chen, Z.; Marsh, E.N.G. Effect of surface crowding and surface hydrophilicity on the activity, stability and molecular orientation of a covalently tethered enzyme. Langmuir 2017, 33, 7152–7159. [Google Scholar] [CrossRef] [PubMed]
- Jonkheijm, P.; Weinrich, D.; Schröder, H.; Niemeyer, C.M.; Waldmann, H. Chemical strategies for generating protein biochips. Angew. Chem. Int. Ed. 2008, 47, 9618–9647. [Google Scholar] [CrossRef] [PubMed]
- Campuzano, S.; Yanez-Sedeno, P.; Pingarron, J.M. Electrochemical affinity biosensors in food safety. Chemosensors 2017, 5, 8. [Google Scholar] [CrossRef]
- Samanta, D.; Sarkar, A. Immobilization of bio-macromolecules on self-assembled monolayers: Methods and sensor applications. Chem. Soc. Rev. 2011, 40, 2567–2592. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wei, S.; Wu, J.; Jasensky, J.; Xi, C.; Li, H.; Xu, Y.; Wang, Q.; Marsh, E.N.G.; Brooks, C.L.; et al. Effects of peptide immobilization sites on the structure and activity of surface-tethered antimicrobial peptides. J. Phys. Chem. C 2015, 119, 7146–7155. [Google Scholar] [CrossRef]
- Onaizi, S.A.; Leong, S.S.J. Tethering antimicrobial peptides: Current status and potential challenges. Biotechnol. Adv. 2011, 29, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Frauenfelder, H.; Chen, G.; Berendzen, J.; Fenimore, P.W.; Jansson, H.; McMahon, B.H.; Stroe, I.R.; Swenson, J.; Young, R.D. A unified model of protein dynamics. Proc. Natl. Acad. Sci. USA 2009, 106, 5129–5134. [Google Scholar] [CrossRef] [PubMed]
- Daniel, R.M.; Dunn, R.V.; Finney, J.L.; Smith, J.C. The role of dynamics in enzyme activity. Annu. Rev. Biophys. Biomol. Struct. 2003, 32, 69–92. [Google Scholar] [CrossRef] [PubMed]
- Swenson, J.; Jansson, H.; Hedström, J.; Bergman, R. Properties of hydration water and its role in protein dynamics. J. Phys. Condens. Matter 2007, 19, 205109. [Google Scholar] [CrossRef]
- Chen, S.H.; Liu, L.; Fratini, E.; Baglioni, P.; Faraone, A.; Mamontov, E. Observation of fragile-to-strong dynamic crossover in protein hydration water. Proc. Natl. Acad. Sci. USA 2006, 103, 9012–9016. [Google Scholar] [CrossRef] [PubMed]
- Gavrilov, Y.; Leuchter, J.D.; Levy, Y. On the coupling between the dynamics of protein and water. Phys. Chem. Chem. Phys. 2017, 19, 8243–8257. [Google Scholar] [CrossRef] [PubMed]
- Scoppola, E.; Sodo, A.; McLain, S.E.; Ricci, M.A.; Bruni, F. Water-peptide site-specific interactions: A structural study on the hydration of glutathione. Biophys. J. 2014, 106, 1701–1709. [Google Scholar] [CrossRef] [PubMed]
- Ebbinghaus, S.; Kim, S.J.; Heyden, M.; Yu, X.; Heugen, U.; Gruebele, M.; Leitner, D.M.; Havenith, M. An extended dynamical hydration shell around proteins. Proc. Natl. Acad. Sci. USA 2007, 104, 20749–20752. [Google Scholar] [CrossRef] [PubMed]
- Levy, Y.; Onuchic, J.N. Water mediation in protein folding and molecular recognition. Annu. Rev. Biophys. Biomol. Struct. 2006, 35, 389–415. [Google Scholar] [CrossRef] [PubMed]
- Flores, S.C.; Kherb, J.; Konelick, N.; Chen, X.; Cremer, P.S. The effects of hofmeister cations at negatively charged hydrophilic surfaces. J. Phys. Chem. C 2012, 116, 5730–5734. [Google Scholar] [CrossRef]
- Gurau, M.C.; Kim, G.; Lim, S.M.; Albertorio, F.; Fleisher, H.C.; Cremer, P.S. Organization of water layers at hydrophilic interfaces. ChemPhysChem 2003, 4, 1231–1233. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.Y.; Lim, S.M.; Albertorio, F.; Kim, G.; Gurau, M.C.; Yang, R.D.; Holden, M.A.; Cremer, P.S. The vroman effect: A molecular level description of fibrinogen displacement. J. Am. Chem. Soc. 2003, 125, 12782–12786. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Gurau, M.; Kim, J.; Cremer, P.S. Investigations of lysozyme adsorption at the air/water and quartz/water interfaces by vibrational sum frequency spectroscopy. Langmuir 2002, 18, 2807–2811. [Google Scholar] [CrossRef]
- Mezger, M.; Reichert, H.; Schöder, S.; Okasinski, J.; Schröder, H.; Dosch, H.; Palms, D.; Ralston, J.; Honkimäki, V. High-resolution in situ X-ray study of the hydrophobic gap at the water-octadecyl-trichlorosilane interface. Proc. Natl. Acad. Sci. USA 2006, 103, 18401–18404. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.J.; Varilly, P.; Chandler, D. Fluctuations of water near extended hydrophobic and hydrophilic surfaces. J. Phys. Chem. B 2010, 114, 1632–1637. [Google Scholar] [CrossRef] [PubMed]
- Nihonyanagi, S.; Yamaguchi, S.; Tahara, T. Water hydrogen bond structure near highly charged interfaces is not like ice. J. Am. Chem. Soc. 2010, 132, 6867–6869. [Google Scholar] [CrossRef] [PubMed]
- Fears, K.P.; Creager, S.E.; Latour, R.A. Determination of the surface pk of carboxylic- and amine-terminated alkanethiols using surface plasmon resonance spectroscopy. Langmuir 2008, 24, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.M.; Hughes, Z.E.; Walsh, T.R. Binding affinities of amino acid analogues at the charged aqueous titania interface: Implications for titania-binding peptides. Langmuir 2014, 30, 13321–13329. [Google Scholar] [CrossRef] [PubMed]
- Corrales Ureña, Y.R.; Wittig, L.; Vieira Nascimento, M.; Faccioni, J.L.; Lisboa Filho, P.N.; Rischka, K. Influences of the ph on the adsorption properties of an antimicrobial peptide on titanium surfaces. Appl. Adhes. Sci. 2015, 3, 7. [Google Scholar] [CrossRef]
- Puddu, V.; Perry, C.C. Peptide adsorption on silica nanoparticles: Evidence of hydrophobic interactions. ACS Nano 2012, 6, 6356–6363. [Google Scholar] [CrossRef] [PubMed]
- Schwierz, N.; Horinek, D.; Netz, R.R. Reversed anionic hofmeister series: The interplay of surface charge and surface polarity. Langmuir 2010, 26, 7370–7379. [Google Scholar] [CrossRef] [PubMed]
- Lund, M.; Jungwirth, P. Patchy proteins, anions and the hofmeister series. J. Phys. Condens. Matter 2008, 20, 494218. [Google Scholar] [CrossRef]
- Boström, M.; Williams, D.R.M.; Ninham, B.W. Specific ion effects: Why the properties of lysozyme in salt solutions follow a hofmeister series. Biophys. J. 2003, 85, 686–694. [Google Scholar] [CrossRef]
- Zhang, Y.; Cremer, P.S. Chemistry of hofmeister anions and osmolytes. Annu. Rev. Phys. Chem. 2010, 61, 63–83. [Google Scholar] [CrossRef] [PubMed]
- Heyda, J.; Lund, M.; Oncak, M.; Slavicek, P.; Jungwirth, P. Reversal of hofmeister ordering for pairing of nh4(+) vs alkylated ammonium cations with halide anions in water. J. Phys. Chem. B 2010, 114, 10843–10852. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cremer, P.S. The inverse and direct hofmeister series for lysozyme. Proc. Natl. Acad. Sci. USA 2009, 106, 15249–15253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cremer, P.S. Interactions between macromolecules and ions: The hofmeister series. Curr. Opin. Chem. Biol. 2006, 10, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Furyk, S.; Bergbreiter, D.E.; Cremer, P.S. Specific ion effects on the water solubility of macromolecules: Pnipam and the hofmeister series. J. Am. Chem. Soc. 2005, 127, 14505–14510. [Google Scholar] [CrossRef] [PubMed]
- Gurau, M.C.; Lim, S.M.; Castellana, E.T.; Albertorio, F.; Kataoka, S.; Cremer, P.S. On the mechanism of the hofmeister effect. J. Am. Chem. Soc. 2004, 126, 10522–10523. [Google Scholar] [CrossRef] [PubMed]
- Kunz, W.; Henle, J.; Ninham, B.W. ‘Zur lehre von der wirkung der salze’ (about the science of the effect of salts): Franz hofmeister’s historical papers. Curr. Opin. Colloid Interface Sci. 2004, 9, 19–37. [Google Scholar] [CrossRef]
- Walsh, T.R.; Knecht, M.R. Biointerface structural effects on the properties and applications of bioinspired peptide-based nanomaterials. Chem. Rev. 2017, 117, 12641–12704. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Flores, S.C.; Lim, S.M.; Zhang, Y.; Yang, T.; Kherb, J.; Cremer, P.S. Specific anion effects on water structure adjacent to protein monolayers. Langmuir 2010, 26, 16447–16454. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yang, T.; Kataoka, S.; Cremer, P.S. Specific ion effects on interfacial water structure near macromolecules. J. Am. Chem. Soc. 2007, 129, 12272–12279. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.R.; Claridge, S.A. Peptide interfaces with graphene: An emerging intersection of analytical chemistry, theory, and materials. Anal. Bioanal. Chem. 2016, 408, 2649–2658. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Jasensky, J.; Li, Y.; Chen, Z. Engineering and characterization of peptides and proteins at surfaces and interfaces: A case study in surface-sensitive vibrational spectroscopy. Acc. Chem. Res. 2016, 49, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Rimola, A.; Costa, D.; Sodupe, M.; Lambert, J.-F.; Ugliengo, P. Silica surface features and their role in the adsorption of biomolecules: Computational modeling and experiments. Chem. Rev. 2013, 113, 4216–4313. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Latour, R.A. Correlation between desorption force measured by atomic force microscopy and adsorption free energy measured by surface plasmon resonance spectroscopy for peptide−surface interactions. Langmuir 2010, 26, 18852–18861. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Yang, Z.; Chong, Y.; Ge, C.; Weber, J.K.; Bell, D.R.; Zhou, R. Surface curvature relation to protein adsorption for carbon-based nanomaterials. Sci. Rep. 2015, 5, 10886. [Google Scholar] [CrossRef] [PubMed]
- Hughes, Z.E.; Tomasio, S.M.; Walsh, T.R. Efficient simulations of the aqueous bio-interface of graphitic nanostructures with a polarisable model. Nanoscale 2014, 6, 5438–5448. [Google Scholar] [CrossRef] [PubMed]
- Paula, A.J.; Silveira, C.P.; Martinez, D.S.T.; Souza Filho, A.G.; Romero, F.V.; Fonseca, L.C.; Tasic, L.; Alves, O.L.; Durán, N. Topography-driven bionano-interactions on colloidal silica nanoparticles. ACS Appl. Mater. Interfaces 2014, 6, 3437–3447. [Google Scholar] [CrossRef] [PubMed]
- Hahm, J.-I. Fundamentals of nanoscale polymer—Protein interactions and potential contributions to solid-state nanobioarrays. Langmuir 2014, 30, 9891–9904. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Kawaguchi, J.; Lee, S.; Yamaguchi, T. On the solvent role in alcohol-induced α-helix formation of chymotrypsin inhibitor 2. Pure Appl. Chem. 2008, 80, 1337. [Google Scholar] [CrossRef]
- Thomas, P.D.; Dill, K.A. Local and nonlocal interactions in globular proteins and mechanisms of alcohol denaturation. Protein Sci. 1993, 2, 2050–2065. [Google Scholar] [CrossRef] [PubMed]
- Wasserberg, D.; Cabanas-Danés, J.; Prangsma, J.; O’Mahony, S.; Cazade, P.-A.; Tromp, E.; Blum, C.; Thompson, D.; Huskens, J.; Subramaniam, V.; et al. Controlling protein surface orientation by strategic placement of oligo-histidine tags. ACS Nano 2017, 11, 9068–9083. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Rusling, J.F.; Dixit, C.K. Site-selective orientated immobilization of antibodies and conjugates for immunodiagnostics development. Methods 2017, 116, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Strop, P.; Liu, S.H.; Dorywalska, M.; Delaria, K.; Dushin, R.G.; Tran, T.T.; Ho, W.H.; Farias, S.; Casas, M.G.; Abdiche, Y.; et al. Location matters: Site of conjugation modulates stability and pharmacokinetics of antibody drug conjugates. Chem. Biol. 2013, 20, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Toseland, C.P. Fluorescent labeling and modification of proteins. J. Chem. Biol. 2013, 6, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Roda, A.; Mirasoli, M.; Michelini, E.; Di Fusco, M.; Zangheri, M.; Cevenini, L.; Roda, B.; Simoni, P. Progress in chemical luminescence-based biosensors: A critical review. Biosens. Bioelectron. 2016, 76, 164–179. [Google Scholar] [CrossRef] [PubMed]
- Mobley, D.L.; Dill, K.A. Binding of small-molecule ligands to proteins: “What you see” is not always “what you get”. Structure 2009, 17, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Boehr, D.D.; Nussinov, R.; Wright, P.E. The role of dynamic conformational ensembles in biomolecular recognition. Nat. Chem. Biol. 2009, 5, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Plattner, N.; Noe, F. Protein conformational plasticity and complex ligand-binding kinetics explored by atomistic simulations and markov models. Nat. Commun. 2015, 6, 7653. [Google Scholar] [CrossRef] [PubMed]
- Karunwi, O.; Baldwin, C.; Griesheimer, G.; Sarupria, S.; Guiseppi-Elie, A. Molecular dynamics simulations of peptide–swcnt interactions related to enzyme conjugates for biosensors and biofuel cells. Nano LIFE 2013, 3, 1343007. [Google Scholar] [CrossRef]
- Abramyan, T.M.; Snyder, J.A.; Thyparambil, A.A.; Stuart, S.J.; Latour, R.A. Cluster analysis of molecular simulation trajectories for systems where both conformation and orientation of the sampled states are important. J. Comput. Chem. 2016, 37, 1973–1982. [Google Scholar] [CrossRef] [PubMed]
- Abramyan, T.M.; Snyder, J.A.; Yancey, J.A.; Thyparambil, A.A.; Wei, Y.; Stuart, S.J.; Latour, R.A. Parameterization of an interfacial force field for accurate representation of peptide adsorption free energy on high-density polyethylene. Biointerphases 2015, 10, 021002. [Google Scholar] [CrossRef] [PubMed]
- Snyder, J.A.; Abramyan, T.; Yancey, J.A.; Thyparambil, A.A.; Wei, Y.; Stuart, S.J.; Latour, R.A. Development of a tuned interfacial force field parameter set for the simulation of protein adsorption to silica glass. Biointerphases 2012, 7, 56. [Google Scholar] [CrossRef] [PubMed]
- Noé, F.; Schütte, C.; Vanden-Eijnden, E.; Reich, L.; Weikl, T.R. Constructing the equilibrium ensemble of folding pathways from short off-equilibrium simulations. Proc. Natl. Acad. Sci. USA 2009, 106, 19011–19016. [Google Scholar] [CrossRef] [PubMed]
- Latour, R.A. Perspectives on the simulation of protein-surface interactions using empirical force field methods. Colloids Surf. B 2014, 124, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Latour, R.A. Molecular simulation of protein-surface interactions: Benefits, problems, solutions, and future directions (review). Biointerphases 2008, 3, FC2–FC12. [Google Scholar] [CrossRef] [PubMed]
- Maximova, T.; Moffatt, R.; Ma, B.; Nussinov, R.; Shehu, A. Principles and overview of sampling methods for modeling macromolecular structure and dynamics. PLoS Comput. Biol. 2016, 12, e1004619. [Google Scholar] [CrossRef] [PubMed]
- Lane, T.J.; Shukla, D.; Beauchamp, K.A.; Pande, V.S. To milliseconds and beyond: Challenges in the simulation of protein folding. Curr. Opin. Struct. Biol. 2013, 23, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Abrams, C.; Bussi, G. Enhanced sampling in molecular dynamics using metadynamics, replica-exchange, and temperature-acceleration. Entropy 2013, 16, 163–199. [Google Scholar] [CrossRef]
- Fiorin, G.; Klein, M.L.; Henin, J. Using collective variables to drive molecular dynamics simulations. Mol. Phys. 2013, 111, 3345–3362. [Google Scholar] [CrossRef]
- Cossio, P.; Marinelli, F.; Laio, A.; Pietrucci, F. Optimizing the performance of bias-exchange metadynamics: Folding a 48-residue lysm domain using a coarse-grained model. J. Phys. Chem. B 2010, 114, 3259–3265. [Google Scholar] [CrossRef] [PubMed]
- Pietrucci, F.; Marinelli, F.; Carloni, P.; Laio, A. Substrate binding mechanism of HIV-1 protease from explicit-solvent atomistic simulations. J. Am. Chem. Soc. 2009, 131, 11811–11818. [Google Scholar] [CrossRef] [PubMed]
- Piana, S.; Laio, A. A bias-exchange approach to protein folding. J. Phys. Chem. B 2007, 111, 4553–4559. [Google Scholar] [CrossRef] [PubMed]
- Hughes, Z.E.; Walsh, T.R. What makes a good graphene-binding peptide? Adsorption of amino acids and peptides at aqueous graphene interfaces. J. Mater. Chem. B 2015, 3, 3211–3221. [Google Scholar] [CrossRef]
- Comer, J.; Chen, R.; Poblete, H.; Vergara-Jaque, A.; Riviere, J.E. Predicting adsorption affinities of small molecules on carbon nanotubes using molecular dynamics simulation. ACS Nano 2015, 9, 11761–11774. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Kang, Z.; Ye, X.; Wu, T.; Zhu, Q. Molecular simulation of flavin adenine dinucleotide immobilized on charged single-walled carbon nanotubes for biosensor applications. Biomaterials 2012, 33, 8757–8770. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ye, X.-S.; Wu, T.; Wang, C.-T.; Shen, J.-W.; Kang, Y. Conformational mobility of gox coenzyme complex on single-wall carbon nanotubes. Sensors 2008, 8, 8453. [Google Scholar] [CrossRef] [PubMed]
- Mijajlovic, M.; Penna, M.J.; Biggs, M.J. Free energy of adsorption for a peptide at a liquid/solid interface via nonequilibrium molecular dynamics. Langmuir 2013, 29, 2919–2926. [Google Scholar] [CrossRef] [PubMed]
- Az’hari, S.; Ghayeb, Y. Effect of chirality, length and diameter of carbon nanotubes on the adsorption of 20 amino acids: A molecular dynamics simulation study. Mol. Simul. 2014, 40, 392–398. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recognition Molecule | Affinity (μM) | Selectivity |
|---|---|---|
| Human serum albumin 1 | 0.019–1 | Not selective |
| Antibody 2 | 0.00001–0.083 | 20 |
| DNA Aptamer 3 | 0.096–0.370 | 6–100 |
| Peptide 4 | 0.00007–29.4 | 3 |
| Petpide Sequence | Screening Approach | Affinity (μM)-Expt 1 | Affinity (μM)-Predicted 2 | Reference |
|---|---|---|---|---|
| SNLHPK | Phage display library | 29.4 | 1991 | [46] |
| CSIVEDGK | Molecular modeling | 12.0 | 1861 | [39] |
| GPAGIDGPAGIRC | Molecular modeling | 16.0 | 2563 | [39] |
| VYMNRKYYKCCK | Mimotype mapping—NADH-FMN oxidoreductase | 0.079 | 1.47 | [47] |
| IRPMVDP | Mimotype mapping—mAb | 0.0017 | NA | [49] |
| AETYGFQLHAMK | Mimotype mapping—2nd gen peptide library | 0.00013 | NA | [50] |
| GMVQTIF with pentamer spacer (GGGSK) and biotin tag | Mimotype mapping—mAb 2A11 | 0.00007 | NA | [44] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thyparambil, A.A.; Bazin, I.; Guiseppi-Elie, A. Molecular Modeling and Simulation Tools in the Development of Peptide-Based Biosensors for Mycotoxin Detection: Example of Ochratoxin. Toxins 2017, 9, 395. https://doi.org/10.3390/toxins9120395
Thyparambil AA, Bazin I, Guiseppi-Elie A. Molecular Modeling and Simulation Tools in the Development of Peptide-Based Biosensors for Mycotoxin Detection: Example of Ochratoxin. Toxins. 2017; 9(12):395. https://doi.org/10.3390/toxins9120395
Chicago/Turabian StyleThyparambil, Aby A., Ingrid Bazin, and Anthony Guiseppi-Elie. 2017. "Molecular Modeling and Simulation Tools in the Development of Peptide-Based Biosensors for Mycotoxin Detection: Example of Ochratoxin" Toxins 9, no. 12: 395. https://doi.org/10.3390/toxins9120395
APA StyleThyparambil, A. A., Bazin, I., & Guiseppi-Elie, A. (2017). Molecular Modeling and Simulation Tools in the Development of Peptide-Based Biosensors for Mycotoxin Detection: Example of Ochratoxin. Toxins, 9(12), 395. https://doi.org/10.3390/toxins9120395