
Cytotoxic Indole Alkaloid 3α-Acetonyltabersonine Induces Glioblastoma Apoptosis via Inhibition of DNA Damage Repair

Abstract
:1. Introduction
2. Results
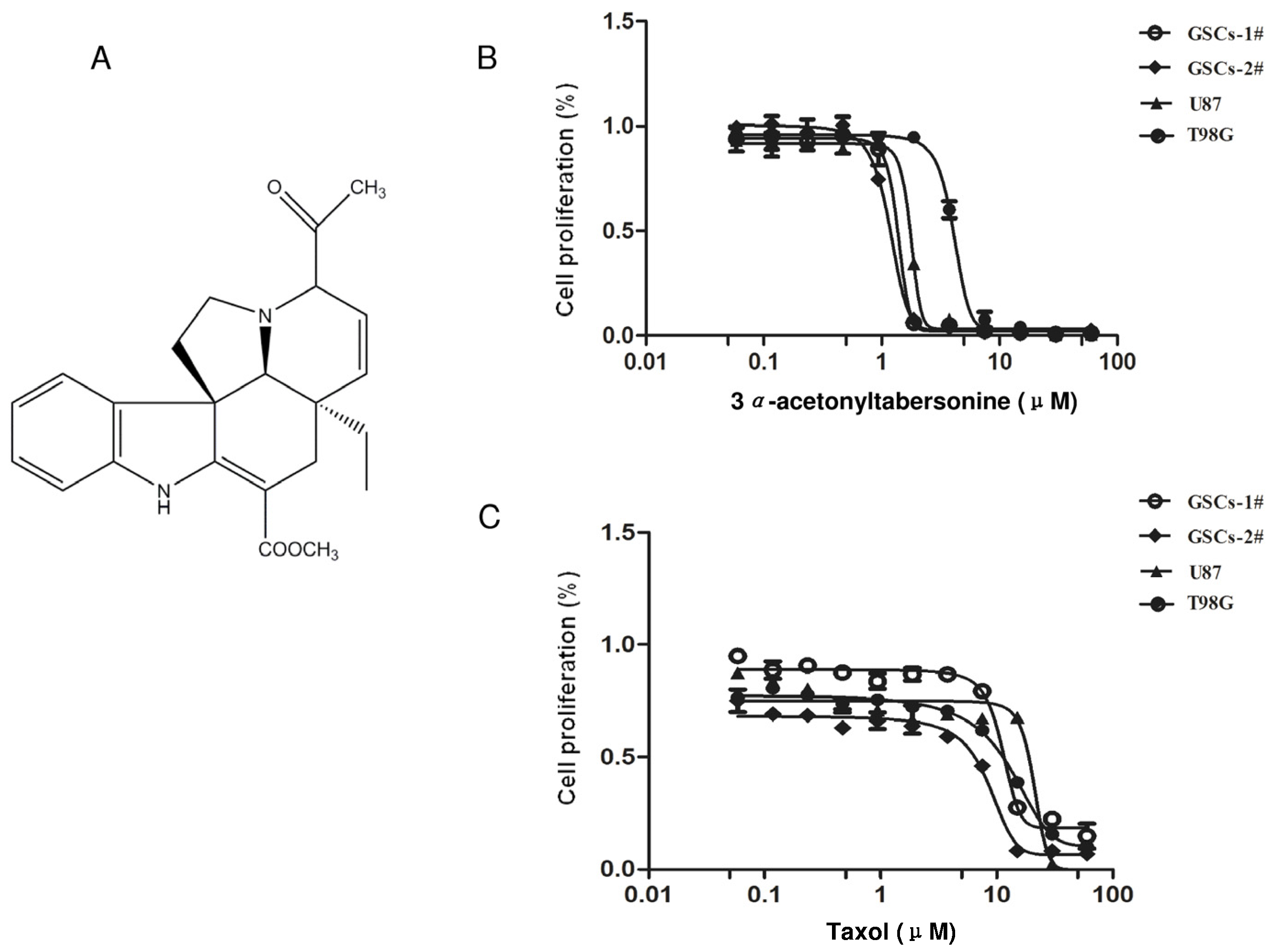
2.1. 3α-Acetonyltabersonine Showed Antitumor Cytotoxicity in Glioblastoma
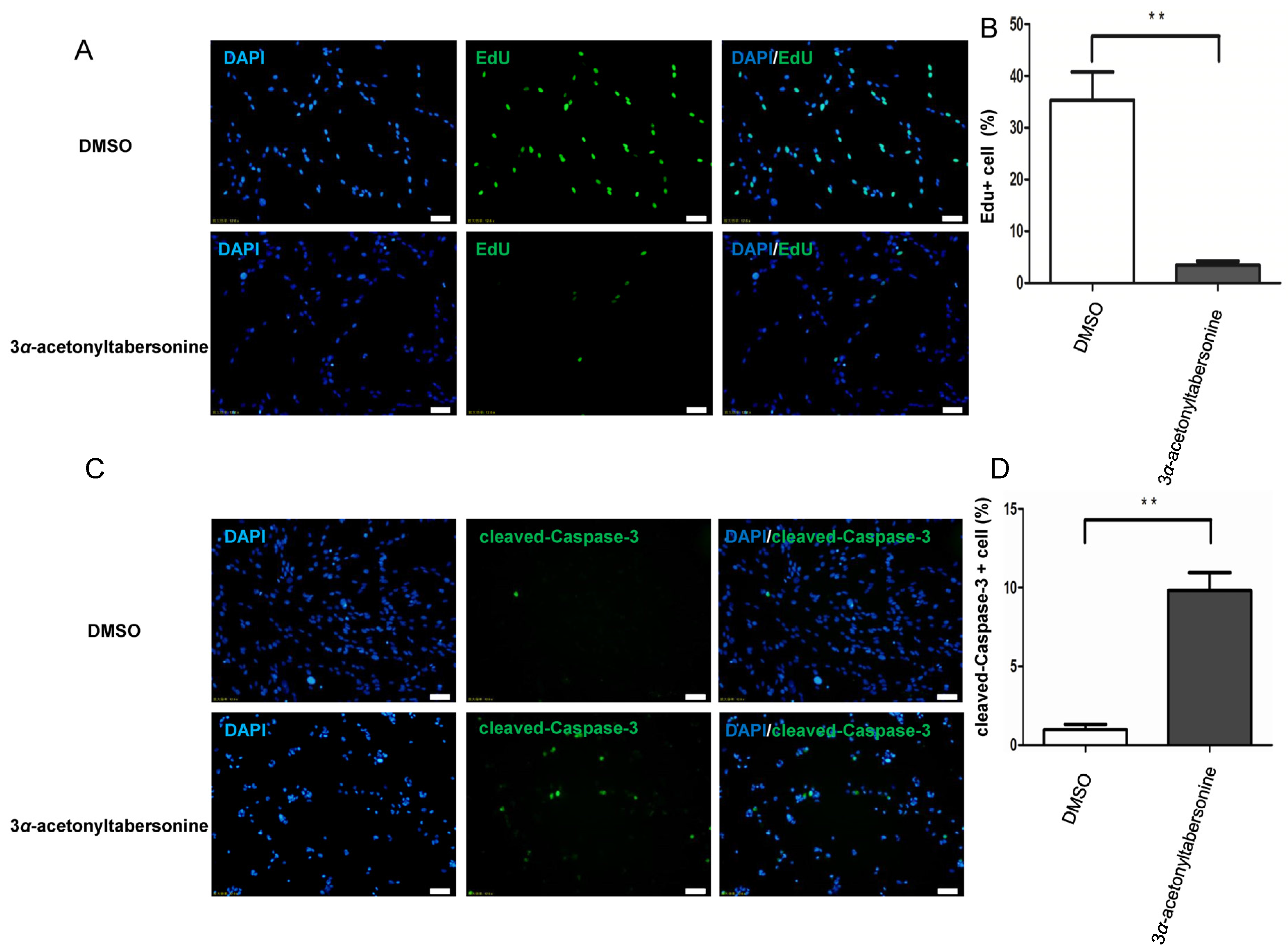
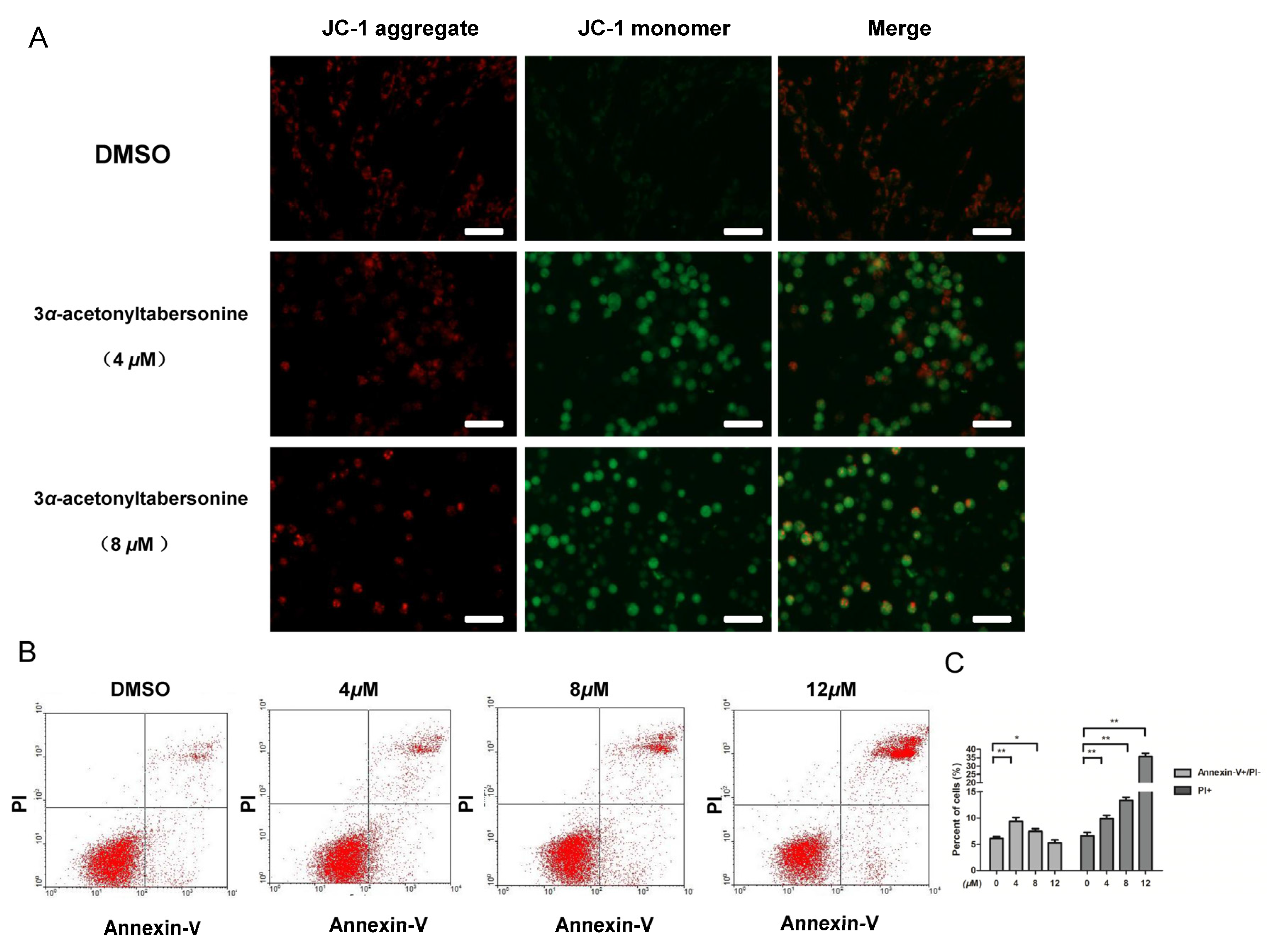
2.2. 3α-Acetonyltabersonine Suppressed Proliferation and Induced Apoptosis in GSCs
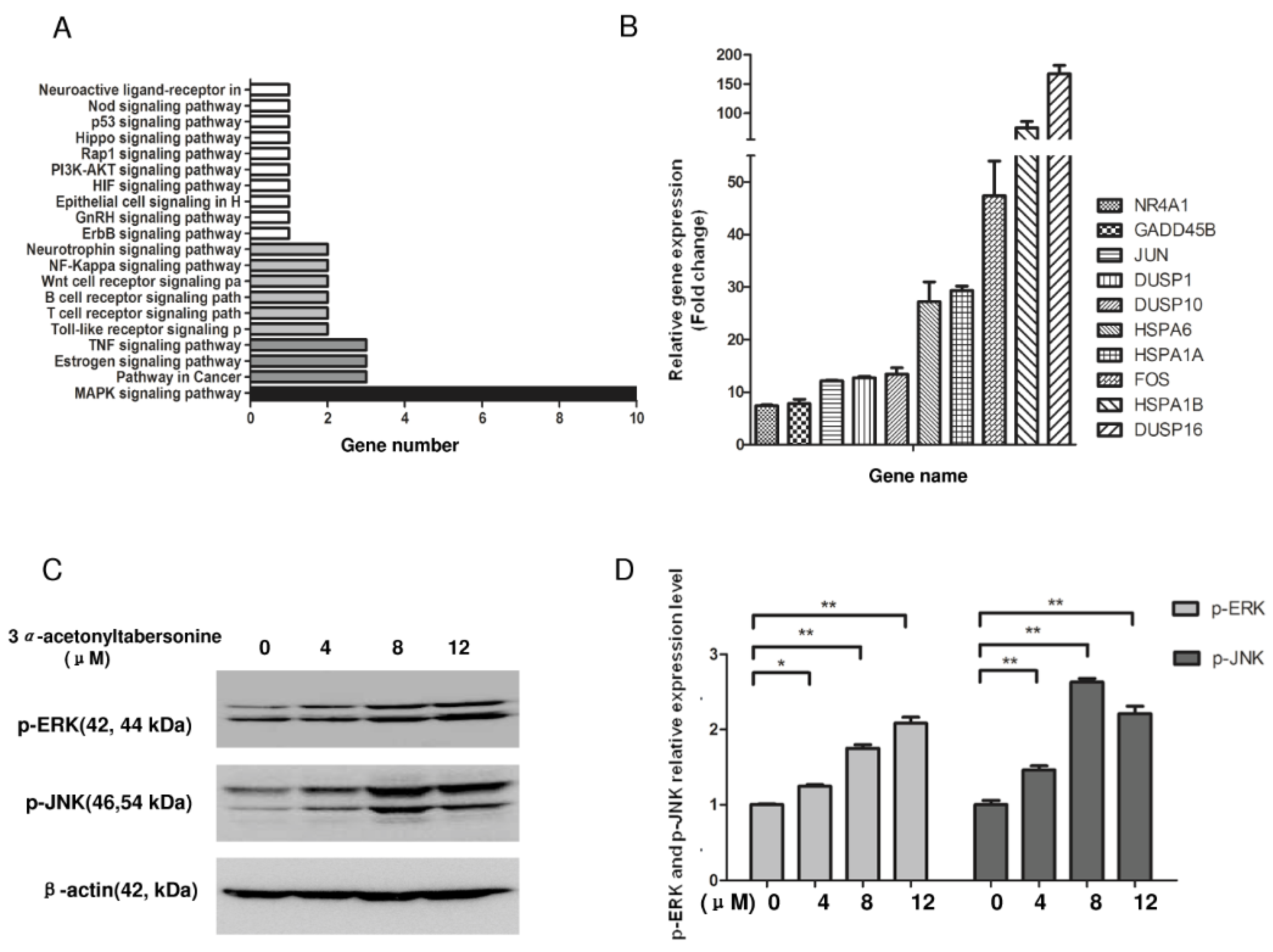
2.3. Phosphorylation of the MAPK Signaling Pathway Was Modulated by the Activity of 3α-Acetonyltabersonine
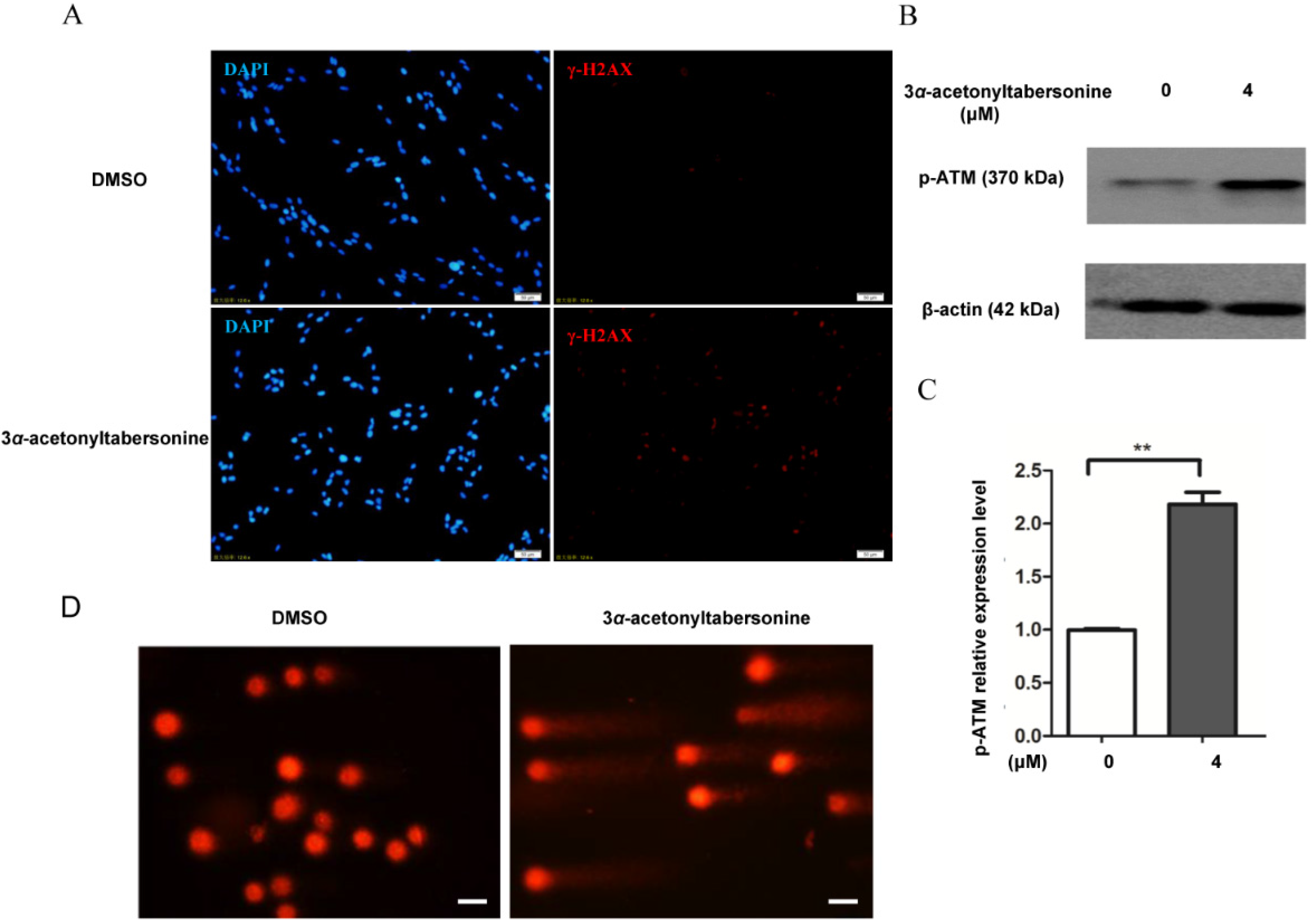
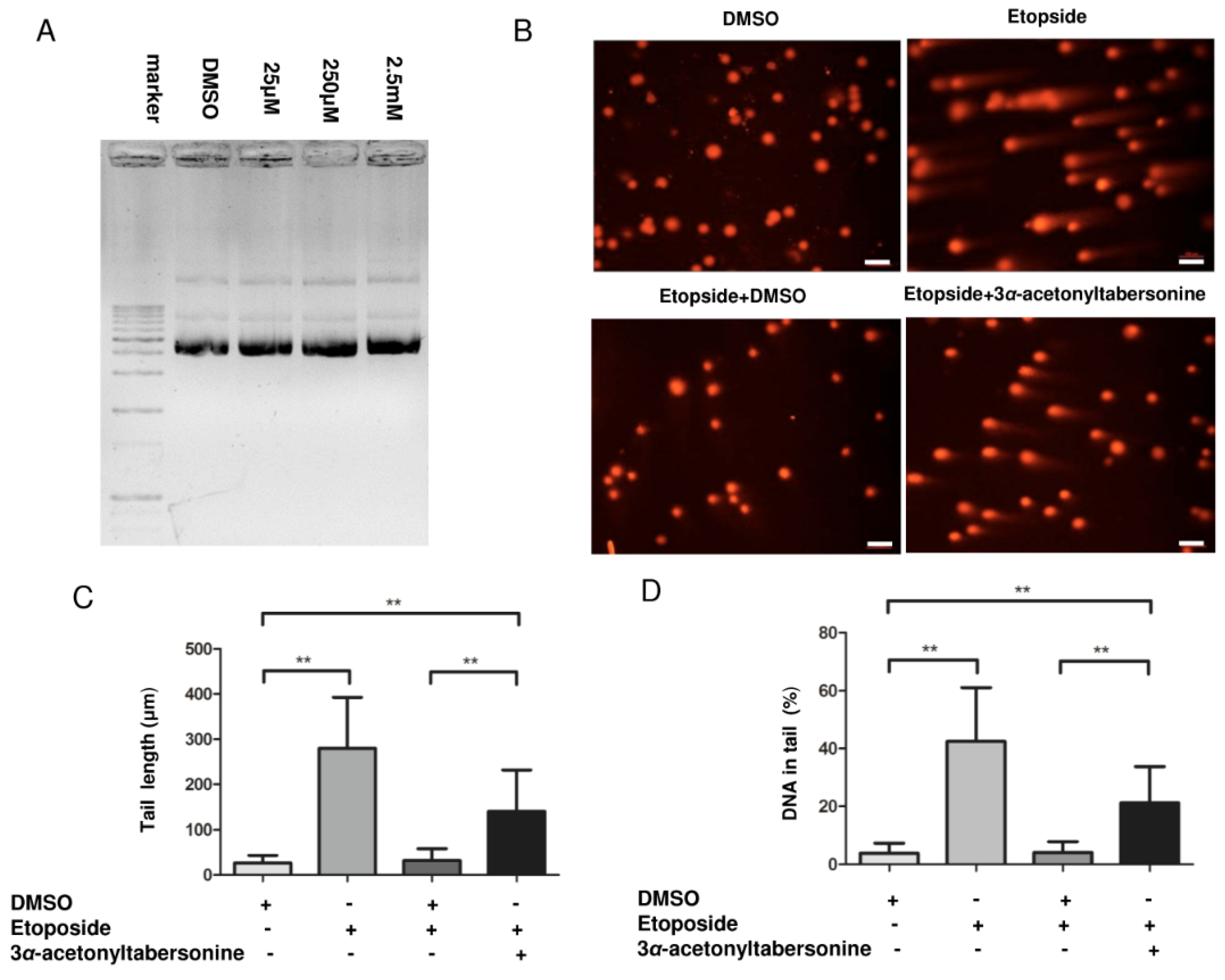
2.4. 3α-Acetonyltabersonine Inhibited DNA Damage Repair
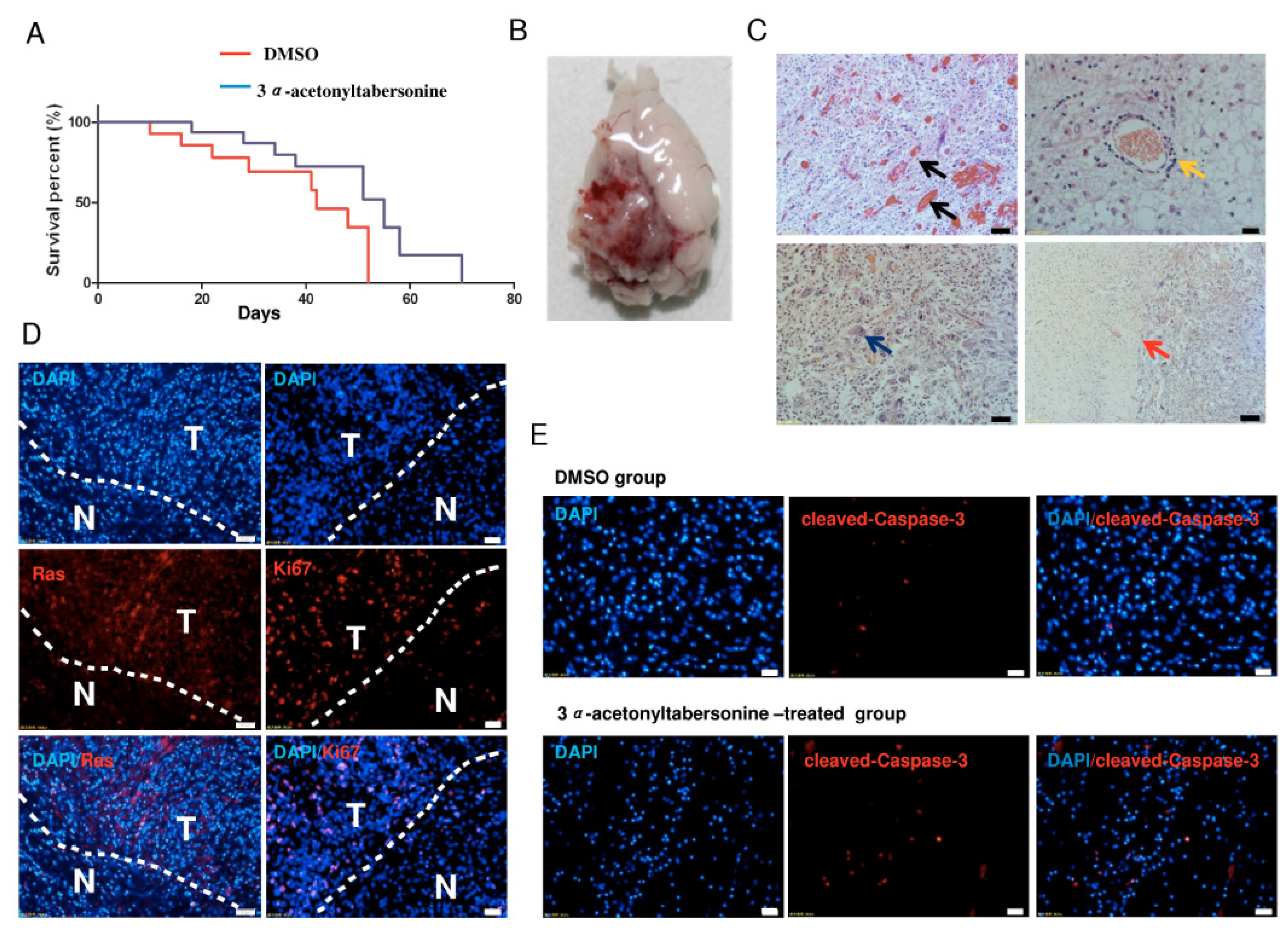
2.5. 3α-Acetonyltabersonine Increased the Lifespan in Glioblastoma Mouse Model
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Cell Culture
5.3. Cytotoxicity Activity
5.4. DNA Proliferation Assay
5.5. Immunofluorescence
5.6. MMP Measurement
5.7. Apoptosis Measurement
5.8. RNA Preparation and RNA-seq
5.9. Real-Time PCR
5.10. Western Blotting
5.11. Comet Assay
5.12. Plasmid Cleavage Assay
5.13. DNA Damage Recovery Assay
5.14. Lentiviral Production
5.15. Intracranial Injections and 3α-Acetonyltabersonine Treatment
5.16. Histology and Immunohistochemistry
5.17. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Spano, V.; Attanzio, A.; Cascioferro, S.; Carbone, A.; Montalbano, A.; Barraja, P.; Tesoriere, L.; Cirrincione, G.; Diana, P.; Parrino, B. Synthesis and antitumor activity of new thiazole nortopsentin analogs. Mar. Drugs 2016, 14, 226. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Parrino, B.; Di Vita, G.; Attanzio, A.; Spano, V.; Montalbano, A.; Barraja, P.; Tesoriere, L.; Livrea, M.A.; Diana, P.; et al. Synthesis and antiproliferative activity of thiazolyl-bis-pyrrolo[2,3-b]pyridines and indolyl-thiazolyl-pyrrolo[2,3-c]pyridines, nortopsentin analogues. Mar. Drugs 2015, 13, 460–492. [Google Scholar] [CrossRef] [PubMed]
- Parrino, B.; Carbone, A.; Di Vita, G.; Ciancimino, C.; Attanzio, A.; Spano, V.; Montalbano, A.; Barraja, P.; Tesoriere, L.; Livrea, M.A.; et al. 3-[4-(1H-indol-3-yl)-1,3-thiazol-2-yl]-1H-pyrrolo[2,3-b]pyridines, nortopsentin analogues with antiproliferative activity. Mar. Drugs 2015, 13, 1901–1924. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Pennati, M.; Barraja, P.; Montalbano, A.; Parrino, B.; Spano, V.; Lopergolo, A.; Sbarra, S.; Doldi, V.; Zaffaroni, N.; et al. Synthesis and antiproliferative activity of substituted 3[2-(1H-indol-3-yl)-1,3-thiazol-4-yl]-1H-pyrrolo[3,2-b]pyridines, marine alkaloid nortopsentin analogues. Curr. Med. Chem. 2014, 21, 1654–1666. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L. Biogenetically patterned synthesis of monoterpenoid indole alkaloids from secologanin and its derivatives. Chin. J. Org. Chem. 2003, 23, 784–793. [Google Scholar]
- Sersa, G.; Krzic, M.; Sentjurc, M.; Ivanusa, T.; Beravs, K.; Cemazar, M.; Auersperg, M.; Swartz, H.M. Reduced tumor oxygenation by treatment with vinblastine. Cancer Res. 2001, 61, 4266–4271. [Google Scholar] [PubMed]
- Jordan, M.A.; Kamath, K. How do microtubule-targeted drugs work? An overview. Curr. Cancer Drug Targets 2007, 7, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.; Marshall, E.; Green, J.A. Ten-year survival in advanced malignant melanoma following treatment with interferon and vindesine. Ann. Oncol. 2000, 11, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Béni, Z.; Háda, V.; Dubrovay, Z.; Szantay, C., Jr. Structure elucidation of indole-indoline type alkaloids: A retrospective account from the point of view of current NMRand MS technology. J. Pharm. Biomed. Anal. 2012, 69, 106–124. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.M.; Feng, T.; Wang, Y.Y.; Li, X.Y.; Ye, Z.N.; An, T.; Qing, C.; Luo, X.D.; Li, Y. Bisleuconothine A, a bisindole alkaloid, inhibits colorectal cancer cell in vitro and in vivo targeting Wnt signaling. Oncotarget 2016, 7, 10203–10214. [Google Scholar] [PubMed]
- Liu, Y.P.; Li, Y.; Cai, X.H.; Li, X.Y.; Kong, L.M.; Cheng, G.G.; Luo, X.D. Melodinines M–U, cytotoxic alkaloids from Melodinus suaveolens. J. Nat. Prod. 2012, 75, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Yang, K.X.; Yang, X.W.; Khan, A.; Liu, L.; Wang, B.; Zhao, Y.L.; Liu, Y.P.; Li, Y.; Luo, X.D. New cytotoxic tigliane diterpenoids from Croton caudatus. Planta Med. 2016, 82, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.G.; Zhao, Y.L.; Zhang, Y.; Lunga, P.K.; Hu, D.B.; Li, Y.; Gu, J.; Song, C.W.; Sun, W.B.; Liu, Y.P.; et al. Indole alkaloids from cultivated Vinca major. Tetrahedron 2014, 70, 8723–8729. [Google Scholar] [CrossRef]
- Gu, J.; Cheng, G.G.; Qian, S.Y.; Li, Y.; Liu, Y.P.; Luo, X.D. Dysoxydensins A-G, seven new clerodane diterpenoids from Dysoxylum densiflorum. Planta Med. 2014, 80, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.G.; Cai, X.H.; Zhang, B.H.; Li, Y.; Gu, J.; Bao, M.F.; Liu, Y.P.; Luo, X.D. Cinchona alkaloids from Cinchona succirubra and Cinchona ledgeriana. Planta Med. 2014, 80, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.F.; Yan, J.M.; Cheng, G.G.; Li, X.Y.; Liu, Y.P.; Li, Y.; Cai, X.H.; Luo, X.D. Cytotoxic indole alkaloids from Tabernaemontana divaricata. J. Nat. Prod. 2013, 76, 1406–1412. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.P.; Zhao, Y.L.; Feng, T.; Cheng, G.G.; Zhang, B.H.; Li, Y.; Cai, X.H.; Luo, X.D. Melosuavines A–H, cytotoxic bisindole alkaloid derivatives from Melodinus suaveolens. J. Nat. Prod. 2013, 76, 2322–2329. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Li, X.N.; Zhang, B.H.; Li, Y.; Cai, X.H.; Liu, Y.P.; Luo, X.D. Gardovatine, a novel strychnos-strychnos bisindole alkaloid with cytotoxicity from Gardneria oveta. Bioorg. Med. Chem. Lett. 2013, 23, 5563–5565. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.H.; Li, Y.; Liu, Y.P.; Li, X.N.; Bao, M.F.; Luo, X.D. Alkaloids from Melodinus yunnanensis. Phytochemistry 2012, 83, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.P.; Cai, X.H.; Feng, T.; Li, Y.; Li, X.N.; Luo, X.D. Triterpene and sterol derivatives from the roots of Breynia fruticosa. J. Nat. Prod. 2011, 74, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Wang, Y.Y.; Su, J.; Li, Y.; Cai, X.H.; Luo, X.D. Amaryllidaceae alkaloids from Lycoris radiata. Helv. Chim. Acta 2011, 94, 178–183. [Google Scholar] [CrossRef]
- Cai, X.H.; Li, Y.; Su, J.; Liu, Y.P.; Li, X.N.; Luo, X.D. Novel indole and quinoline alkaloids from Melodinus yunnanensis. Nat. Prod. Bioprospect. 2011, 1, 25–28. [Google Scholar] [CrossRef]
- Cai, X.H.; Jiang, H.; Li, Y.; Cheng, G.G.; Liu, Y.P.; Feng, T.; Luo, X.D. Cytotoxic indole alkaloids from Melodinus fusiformis and M. morsei. Chin. J. Nat. Med. 2011, 9, 259–263. [Google Scholar] [CrossRef]
- Feng, T.; Li, Y.; Liu, Y.P.; Cai, X.H.; Wang, Y.Y.; Luo, X.D. Melotenine A, a cytotoxic monoterpenoid indole alkaloid from Melodinus tenuicaudatus. Org. Lett. 2010, 12, 968–971. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Cai, X.H.; Liu, Y.P.; Li, Y.; Wang, Y.Y.; Luo, X.D. Melodinines A-G, monoterpenoid indole alkaloids from Melodinus henryi. J. Nat. Prod. 2010, 73, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Li, Y.; Wang, Y.Y.; Cai, X.H.; Liu, Y.P.; Luo, X.D. Cytotoxic indole alkaloids from Melodinus tenuicaudatus. J. Nat. Prod. 2010, 73, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Li, Y.; Wang, Y.Y.; Cai, X.H.; Feng, T.; Luo, X.D. Tirucallane-type alkaloids from the bark of Dysoxylum laxiracemosum. J. Nat. Prod. 2010, 73, 1385–1388. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.H.; Wang, Y.Y.; Zhao, P.J.; Li, Y.; Luo, X.D. Dolabellane diterpenoids from Aglaia odorata. Phytochemistry 2010, 71, 1020–1024. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Li, Y.; Cai, X.H.; Gong, X.; Liu, Y.P.; Zhang, R.T.; Zhang, X.Y.; Tan, Q.G.; Luo, X.D. Monoterpenoid indole alkaloids from Alstonia yunnanensis. J. Nat. Prod. 2009, 72, 1836–1841. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Clarke, I.D.; Terasaki, M.; Bonn, V.E.; Hawkins, C.; Squire, J.; Dirks, P.B. Identification of a cancer stem cell in human brain tumors. Cancer Res. 2003, 63, 5821–5828. [Google Scholar] [PubMed]
- Frank, N.Y.; Schatton, T.; Frank, M.H. The therapeutic promise of the cancer stem cell concept. J. Clin. Investig. 2010, 120, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Croker, A.K.; Allan, A.L. Cancer stem cells: Implications for the progression and treatment of metastatic disease. J. Cell. Mol. Med. 2008, 12, 374–390. [Google Scholar] [CrossRef] [PubMed]
- Lamy, E.; Herz, C.; Lutz-Bonengel, S.; Hertrampf, A.; Marton, M.R.; Mersch-Sundermann, V. The MAPK pathway signals telomerase modulation in response to isothiocyanate-induced DNA damage of human liver cancer cells. PLoS ONE 2013, 8, e53240. [Google Scholar] [CrossRef] [PubMed]
- Nandi, S.; Reinert, L.S.; Hachem, A.; Mazan-Mamczarz, K.; Hagner, P.; He, H.; Gartenhaus, R.B. Phosphorylation of MCT-1 by p44/42 mapk is required for its stabilization in response to DNA damage. Oncogene 2007, 26, 2283–2289. [Google Scholar] [CrossRef] [PubMed]
- Colis, L.C.; Woo, C.M.; Hegan, D.C.; Li, Z.; Glazer, P.M.; Herzon, S.B. The cytotoxicity of (-)-lomaiviticin A arises from induction of double-strand breaks in DNA. Nat. Chem. 2014, 6, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Niola, F.; Zhao, X.; Singh, D.; Sullivan, R.; Castano, A.; Verrico, A.; Zoppoli, P.; Friedmann-Morvinski, D.; Sulman, E.; Barrett, L.; et al. Mesenchymal high-grade glioma is maintained by the ID-RAP1 axis. J. Clin. Investig. 2013, 123, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Adiseshaiah, P.P.; Clogston, J.D.; McLeland, C.B.; Rodriguez, J.; Potter, T.M.; Neun, B.W.; Skoczen, S.L.; Shanmugavelandy, S.S.; Kester, M.; Stern, S.T.; et al. Synergistic combination therapy with nanoliposomal C6-ceramide and vinblastine is associated with autophagy dysfunction in hepatocarcinoma and colorectal cancer models. Cancer Lett. 2013, 337, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Passer, B.J.; Cheema, T.; Wu, S.; Wu, C.L.; Rabkin, S.D.; Martuza, R.L. Combination of vinblastine and oncolytic herpes simplex virus vector expressing IL-12 therapy increases antitumor and antiangiogenic effects in prostate cancer models. Cancer Gene Ther. 2013, 20, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.; Dingle, B.; Rodrigues, G.; Vincent, M.; Ash, R.; Dar, R.; Inculet, R.; Kocha, W.; Malthaner, R.; Sanatani, M.; et al. Analysis of a novel protocol of combined induction chemotherapy and concurrent chemoradiation in unresected non-small-cell lung cancer: a ten-year experience with vinblastine, Cisplatin, and radiation therapy. Clin. Lung Cancer 2010, 11, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Ospovat, I.; Siegelmann-Danieli, N.; Grenader, T.; Hubert, A.; Hamburger, T.; Peretz, T. Mitomycin C and vinblastine: An active regimen in previously treated breast cancer patients. Tumori 2009, 95, 683–686. [Google Scholar] [PubMed]
- Lipton, J.H.; Gospodarowicz, M.; Reingold, S. Acute myeloid leukemia following therapy of Hodgkin’s disease with radiotherapy and ABVD (doxorubicin, bleomycin, vinblastine, and dacarbazine). Hematol. Oncol. 1996, 14, 29–31. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, Z.; Wang, Y.; He, X. Cytotoxic indole alkaloids against human leukemia cell lines from the toxic plant Peganum harmala. Toxins 2015, 7, 4507–4518. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Debatin, K.M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef] [PubMed]
- Hothi, P.; Martins, T.J.; Chen, L.; Deleyrolle, L.; Yoon, J.G.; Reynolds, B.; Foltz, G. High-throughput chemical screens identify disulfiram as an inhibitor of human glioblastoma stem cells. Oncotarget 2012, 3, 1124–1136. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.; Bhaskara, V.K.; Babu, P.P. Implications of mitogen-activated protein kinase signaling in glioma. J. Neurosci. Res. 2016, 94, 114–127. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.K.; Choi, C.H.; Oh, H.L.; Kim, Y.K. Role of ERK activation in cisplatin-induced apoptosis in A172 human glioma cells. Neurotoxicology 2004, 25, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Pommier, Y. Topoisomerase I inhibitors: Camptothecins and beyond. Nat. Rev. Cancer 2006, 6, 789–802. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, I.B.; Lim, K.H.; Kam, T.S.; Loh, H.S. Synergistic cytotoxic effects of combined δ-tocotrienol and jerantinine B on human brain and colon cancers. J. Ethnopharmacol. 2016, 184, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Azari, H.; Rahman, M.; Sharififar, S.; Reynolds, B.A. Isolation and expansion of the adult mouse neural stem cells using the neurosphere assay. J. Vis. Exp. 2010, 45, e2393. [Google Scholar] [CrossRef] [PubMed]
- Eng, C.H.; Wang, Z.C.; Tkach, D.; Toral-Barza, L.; Ugwonali, S.; Liu, S.M.; Fitzgerald, S.L.; George, E.; Frias, E.; Cochran, N.; et al. Macroautophagy is dispensable for growth of KRAS mutant tumors and chloroquine efficacy. Proc. Natl. Acad. Sci. USA 2016, 113, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ding, Y.; Chen, C.H.; Zhou, Z.M.; Ding, C.Y.; Chen, H.Y.; Zhou, J.; Chen, C.S. A new oridonin analog suppresses triple-negative breast cancer cells and tumor growth via the induction of death receptor 5. Cancer Lett. 2016, 380, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Buommino, E.; Schiraldi, C.; Baroni, A.; Paoletti, I.; Lamberti, M.; De Rosa, M.; Tufano, M.A. Ectoine from halophilic microorganisms induces the expression of hsp70 and hsp70b’ in human keratinocytes modulating the proinflammatory response. Cell Stress Chaperones 2005, 10, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Parsian, A.J.; Sheren, J.E.; Tao, T.Y.; Goswami, P.C.; Malyapa, R.; van Rheeden, R.; Watson, M.S.; Hunt, C.R. The human hsp70b gene at the hspa7 locus of chromosome 1 is transcribed but non-functional. Biochim. Acta BBA-Gene Struct. Expr. 2000, 1494, 201–205. [Google Scholar] [CrossRef]
- Gavin, D.P.; Sharma, R.P.; Chase, K.A.; Matrisciano, F.; Dong, E.; Guidotti, A. Growth arrest and DNA-damage-inducible, beta (gadd45b)-mediated DNA demethylation in major psychosis. Neuropsychopharmacology 2012, 37, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhang, S.; Feng, K.; Wu, F.; Wan, Y.; Wang, Z.; Zhang, J.; Wang, Y.; Yan, W.; Fu, Z.; et al. MicroRNA-125b-2 confers human glioblastoma stem cells resistance to temozolomide through the mitochondrial pathway of apoptosis. Int. J. Oncol. 2012, 40, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Zhang, W.D.; Duan, Y.L.; Lu, Y.Q.; Cun, Y.X.; Li, C.H.; Guo, K.; Nie, W.H.; Li, L.; Zhang, R.G.; et al. Filia is an ESC-specific regulator of DNA damage response and safeguards genomic stability. Cell Stem. Cell 2015, 16, 684–698. [Google Scholar] [CrossRef] [PubMed]
- Marumoto, T.; Tashiro, A.; Friedmann-Morvinski, D.; Scadeng, M.; Soda, Y.; Gage, F.H.; Verma, I.M. Development of a novel mouse glioma model using lentiviral vectors. Nat. Med. 2009, 15, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Carro, M.S.; Lim, W.K.; Alvarez, M.J.; Bollo, R.J.; Zhao, X.; Snyder, E.Y.; Sulman, E.P.; Anne, S.L.; Doetsch, F.; Colman, H.; et al. The transcriptional network for mesenchymal transformation of brain tumours. Nature 2010, 463, 318–325. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| HSPA6 | CTCCAGCATCCGACAAGAAGC | CTCCAGCATCCGACAAGAAGC [53] |
| HSPA1B | CCCCATCATCAGCGGACTG | AACACCCTTACAGTATCAAC [54] |
| HSPA1A | CCCCATCATCAGCGGACTG | GGCAAGTTCAGTACTTCACC [54] |
| FOS | CAACTTCAT TCCCACGGTCA | TGGCAATCTCGGTCTGCAAA a |
| JUN | GCGTTAGCATGAGTTGGCAC | CGCATGAGGAACCGCATCGC b |
| NR4A1 | CCCTGAAGTTGTTCCCCTCAC | GCCCTCAAGGTGTGGAGAAG c |
| GADD45B | ATTGCAACATGACGCTGGAAGAGC | GATGAGCGTGAAGTGGATT [55] |
| DUSP16 | CACACCACCATTACATCATCG | AACAGTCTGAAGAGAGAGAGGC [55] |
| DUSP10 | GCGGCAGTACTTTGAAGAGGCTTT | AGTCATGGTCATCCGAGTGTGCTT [56] |
| DUSP1 | CGAAGCGTTTTCGGCTTCC | CACCCTGATCGTAGAGTGG [56] |
| Name | Product Number | Postscript |
|---|---|---|
| cleaved-Caspase-3 | CST, 9661 | Primary antibody |
| γ-H2AX | Abcam, ab2893 | Primary antibody |
| p-ERK | CST, 4370 | Primary antibody |
| p-JNK | CST, 4671 | Primary antibody |
| p-ATM | Abcam, 36810 | Primary antibody |
| β-actin | Sigma, A1978 | Primary antibody |
| Flag | Sigma, F2555 | Primary antibody |
| Ki67 | Vector, vp-K452 | Primary antibody |
| Goat anti-Rabbit IgG Fc Dylight 488 | Abcam, ab98462 | Secondary antibody |
| Goat anti -Rabbit IgG (Cy3) | Abcam, ab6939 | Secondary antibody |
| Goat anti-Mouse IgG (Cy3) | Abcam, ab97035 | Secondary antibody |
| Goat anti-Rabbit IgG peroxidase conjugate | Sigma, A6154 | Secondary antibody |
| Goat anti-Mouse IgG peroxidase conjugate | Sigma, A4416 | Secondary antibody |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhao, Y.; Zhou, X.; Ni, W.; Dai, Z.; Yang, D.; Hao, J.; Luo, L.; Liu, Y.; Luo, X.; et al. Cytotoxic Indole Alkaloid 3α-Acetonyltabersonine Induces Glioblastoma Apoptosis via Inhibition of DNA Damage Repair. Toxins 2017, 9, 150. https://doi.org/10.3390/toxins9050150
Li Y, Zhao Y, Zhou X, Ni W, Dai Z, Yang D, Hao J, Luo L, Liu Y, Luo X, et al. Cytotoxic Indole Alkaloid 3α-Acetonyltabersonine Induces Glioblastoma Apoptosis via Inhibition of DNA Damage Repair. Toxins. 2017; 9(5):150. https://doi.org/10.3390/toxins9050150
Chicago/Turabian StyleLi, Yuan, Yunli Zhao, Xia Zhou, Wei Ni, Zhi Dai, Dong Yang, Junjun Hao, Lin Luo, Yaping Liu, Xiaodong Luo, and et al. 2017. "Cytotoxic Indole Alkaloid 3α-Acetonyltabersonine Induces Glioblastoma Apoptosis via Inhibition of DNA Damage Repair" Toxins 9, no. 5: 150. https://doi.org/10.3390/toxins9050150
APA StyleLi, Y., Zhao, Y., Zhou, X., Ni, W., Dai, Z., Yang, D., Hao, J., Luo, L., Liu, Y., Luo, X., & Zhao, X. (2017). Cytotoxic Indole Alkaloid 3α-Acetonyltabersonine Induces Glioblastoma Apoptosis via Inhibition of DNA Damage Repair. Toxins, 9(5), 150. https://doi.org/10.3390/toxins9050150