
Content of the Saponin Protodioscin in Brachiaria spp. from the Eastern Plains of Colombia


Abstract
:1. Introduction
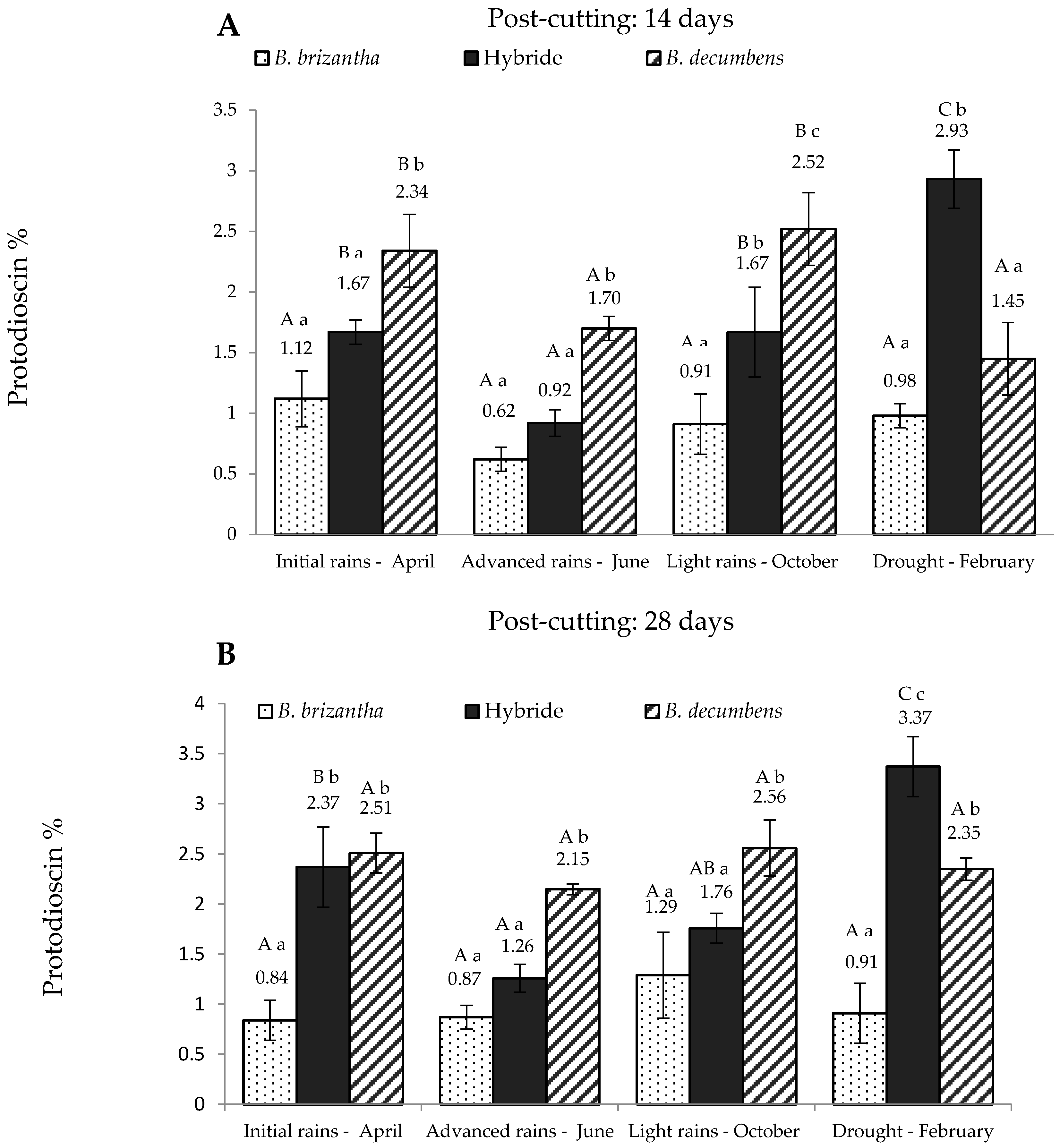
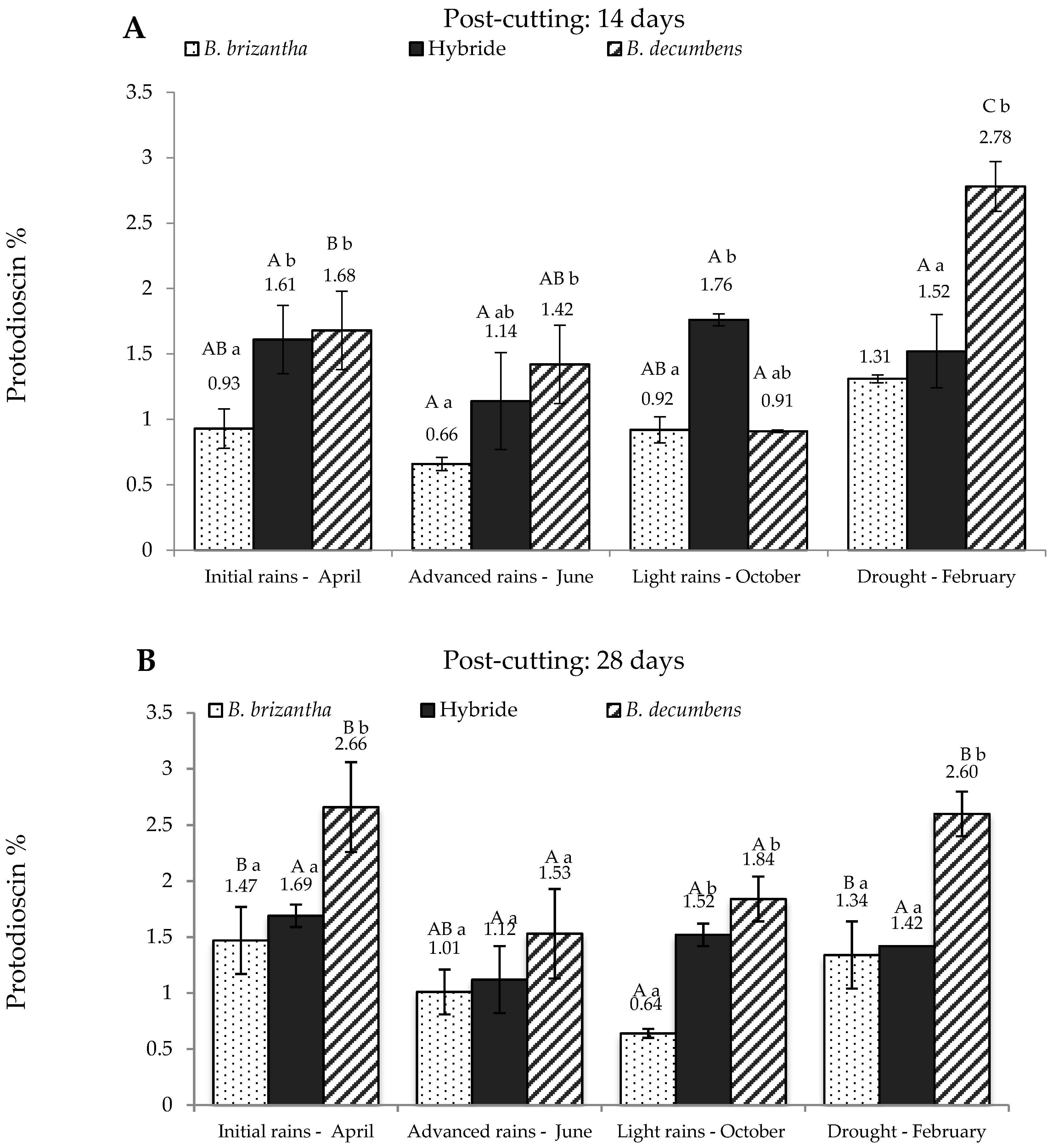
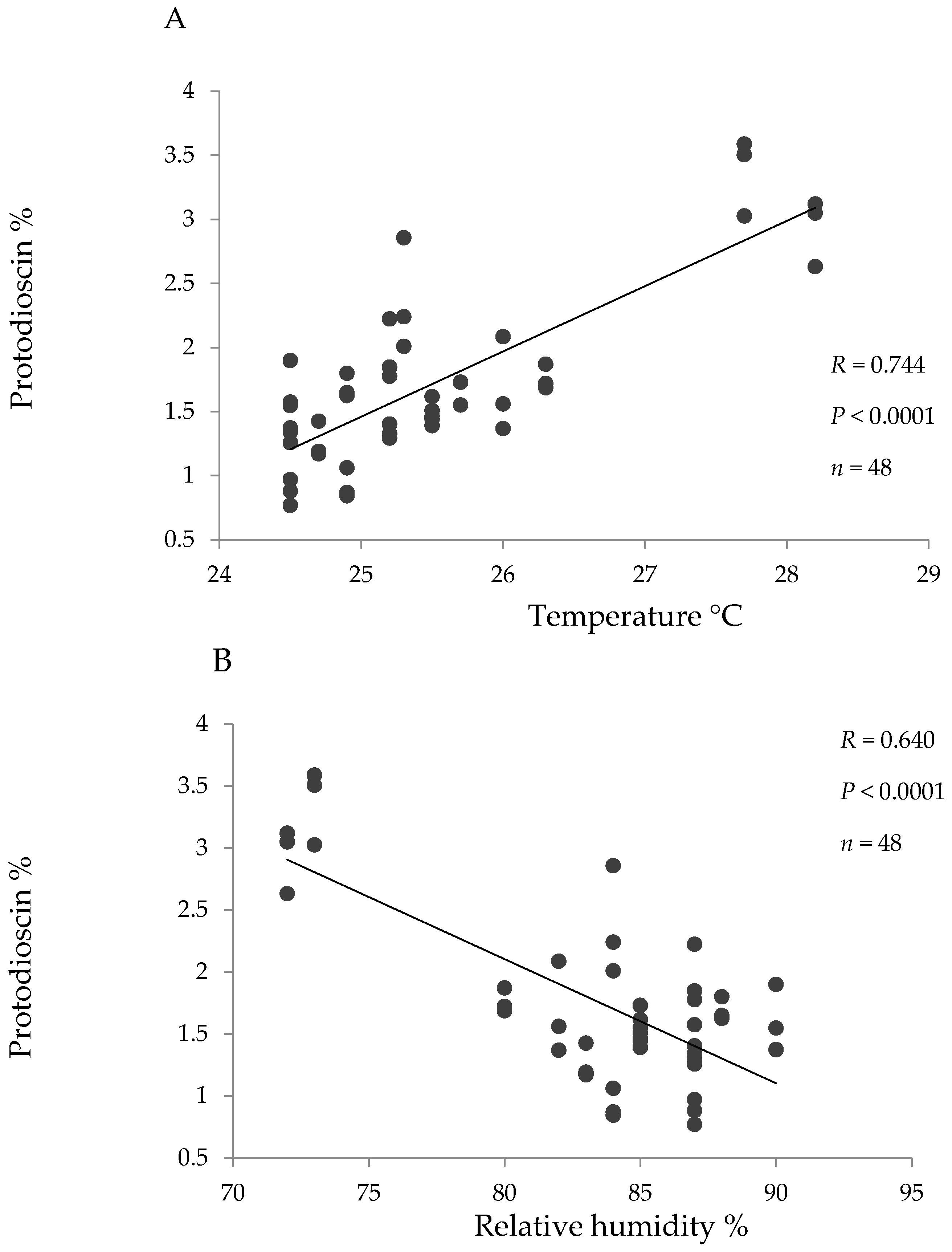
2. Results
3. Discussion
4. Materials and Methods
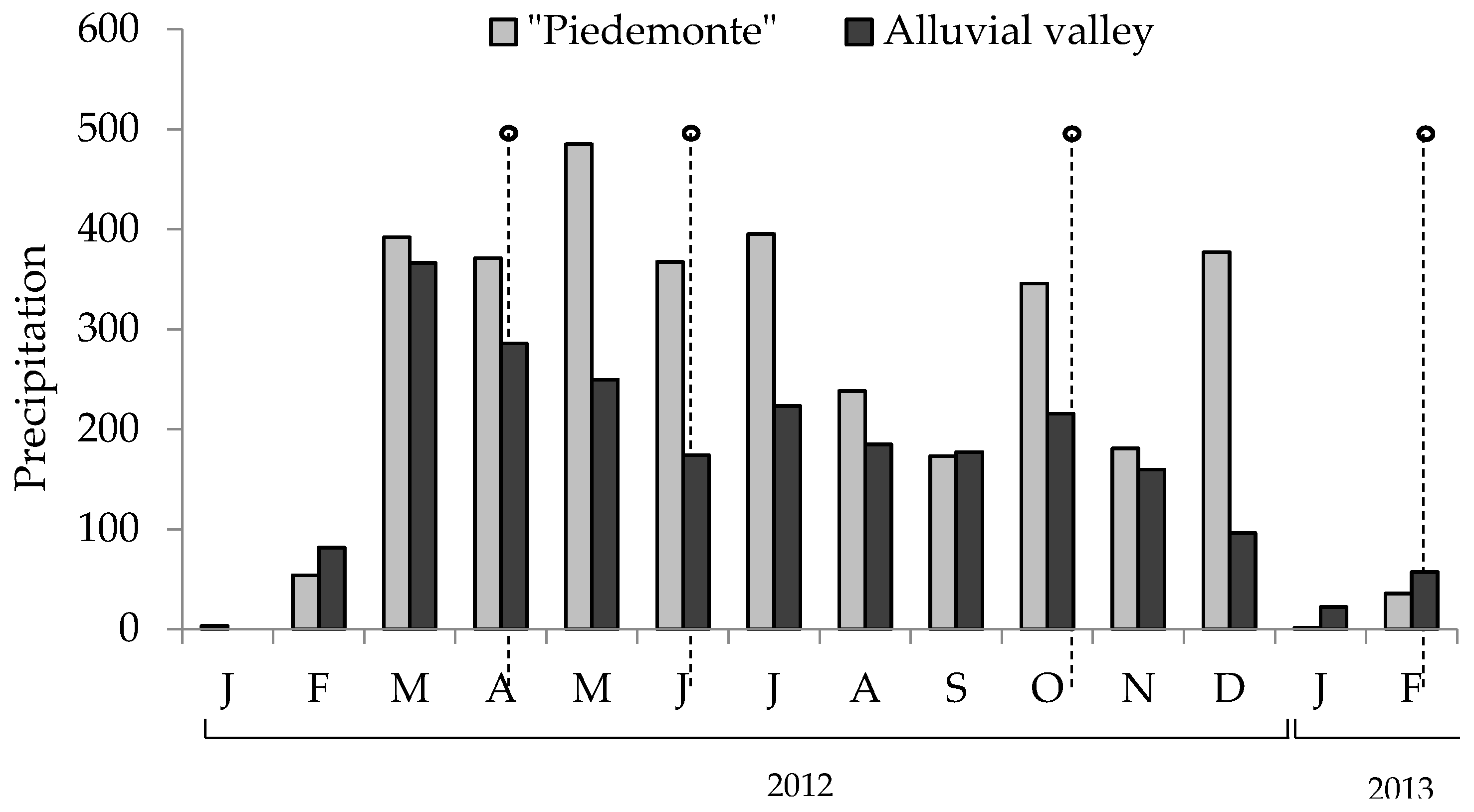
4.1. Region of Study
4.2. Sampling
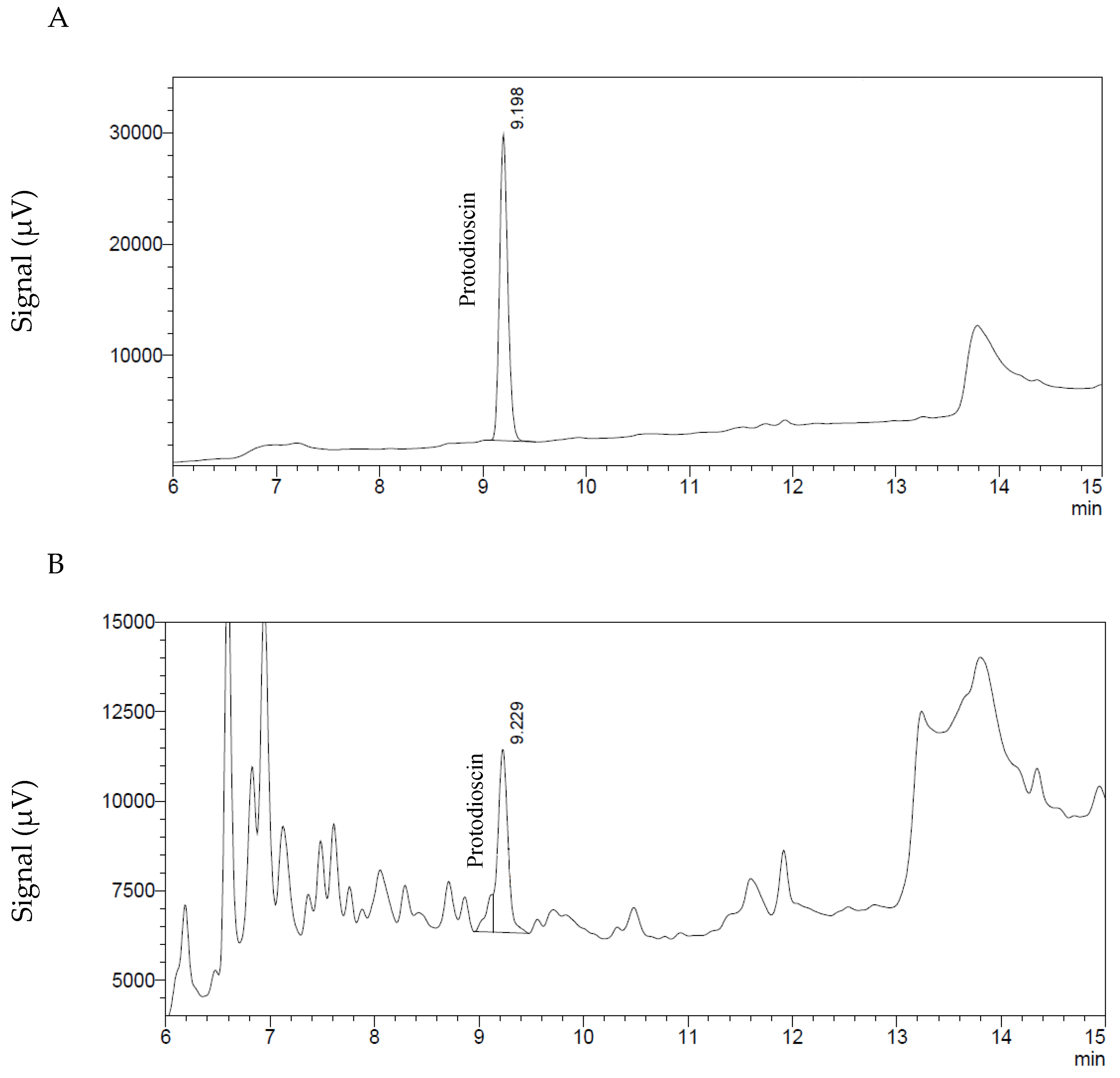
4.3. Determination and Quantification of Protodioscin
Acknowledgments
Author Contributions
Conflicts of Interest
References
- ICA (Instituto Colombiano Agropecuario). Censo Pecuario Nacional 2016. Consolidado por Especies. Available online: http://www.ica.gov.co/getdoc/8232c0e5-be97-42bd-b07b-9cdbfb07fcac/Censos-2008.aspx Censo Pecuario Nacional-2016 (accessed on 24 September 2016).
- WWF (World Wild Foundation). Tropical and Subtropical Grasslands, Savannas and Shrublands. Nothern South America in Colombia and Venezuela. Available online: https://www.worldwildlife.org/ecoregions/nt0709 (accessed on 9 June 2017).
- Argell, P.; Keller-Grein, G. Regional experience with Brachiaria: Tropical America-Humid Lowlands. In Brachiaria: Biology, Agronomy and Improvement; Miles, B.L., Maass, C.B., Eds.; CIAT-EMBRAPA: Cali, Colombia, 1996; pp. 205–224. ISBN 958-9439-57-8. [Google Scholar]
- Lozano, M.; Doncel, B.; Moreno, C. Manual de Plantas Tóxicas Para Bovinos. Región Llanos Orientales de Colombia: Meta y Casanare; Universidad Nacional de Colombia: Bogotá, Colombia, 2011; p. 60. [Google Scholar]
- Diaz, G. Plantas Tóxicas de Importancia en Salud y Producción Animal en Colombia; Editorial UN Universidad Nacional de Colombia: Bogotá, Colombia, 2010; pp. 154–177. [Google Scholar]
- Riet-Correa, B.; Castro, M.B.; De Lemos, R.A.; Riet-Correa, G.; Mustafa, V.; Riet-Correa, F.; Lemos, R. Brachiaria spp. poisoning of ruminants in Brazil. Pesq. Vet. Bras. 2011, 31, 183–192. Available online: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-736X2011000300001&lng=en&nrm=iso&tlng=en (accessed on 10 January 2014).
- Brum, K.; Haraguchi, M.; Lemos, R.; Riet-Correa, F.; Fioravanti, M. Crystal-associated cholangiopathy in sheep grazing Brachiaria decumbens containing the saponin protodioscin protodioscin. Pesq. Vet. Bras. 2000, 27, 39–42. Available online: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-736X2007000100007 (accessed on 24 April 2016).
- Cruz, C.; Driemeier, D.; Pires, V.; Colodel, E.; Daketa, A. Isolation of steroidal sapogenins implicated in experimentally induced cholangiopathy of sheep grazing Brachiaria decumbens in Brazil. Vet. Hum. Toxicol. 2000, 43, 142–145. [Google Scholar]
- Cruz, C.; Driemeier, D.; Pires, V.; Schenkel, E. Experimentally induced cholangiohepatopathy by dosing sheep with fractionated extracts from Brachiaria decumbens. J. Vet. Diagn. Investig. 2001, 13, 170–172. Available online: http://journals.sagepub.com/doi/pdf/10.1177/104063870101300215 (accessed on 15 March 2015). [CrossRef] [PubMed]
- Miles, C.; Wilkins, A.; Munday, S.; Flayoen, A.; Holland, P.; Smith, B. Identification of insoluble salts of the β-D-glucuronides of episarsasapogenin and epismilagenin in the bile of lambs with alved and examination of Narthecium ossifragum, Tribulus terrestris and Panicum miliaceum for sapogenins. J. Agric. Food Chem. 1993, 41, 914–917. [Google Scholar] [CrossRef]
- Miles, C.; Wilkins, A.; Munday, S.; Holland, P.; Smith, B.; Lancaster, M. Identification of the calcium salt of epismilagenin beta-D-glucuronide in the bile crystals of sheep affected by Panicum dichotomiflorum and Panicum schinzii toxicoses. J. Agric. Food Chem. 1992, 40, 1606–1609. [Google Scholar] [CrossRef]
- Mysterud, I.; Flåøyen, A.; Loader, J.I.; Wilkins, A.L. Sapogenin levels in Narthecium ossifragum plants and Ovis aries lamb faeces during two alveld outbreaks in Møre og Romsdal, Norway, 2001. Vet. Res. Commun. 2007, 31, 895–908. Available online: https://link.springer.com/article/10.1007%2Fs11259-007-3561-z (accessed on 13 July 2013). [CrossRef] [PubMed]
- Torres, P.; Diaz, G.J.; Cárdenas, E.; Lozano, M.C. Ethnobotanical Study of Plants Poisonous to Cattle in Eastern Colombia. IJPPR 2012, 2, 14–19. Available online: https://www.ars.usda.gov/ARSUserFiles/oc/np/PoisonousPlants/Fall2012/colombia.pdf (accessed on 10 July 2013).
- Chong Dubé, D.; Figueredo, J.M.; Percedo, M.I.; Domínguez, P.; Martínez García, Y.; Alfonso, P.; Marrero Faz, E. Toxicosis por pasto Mulato (Brachiaria ruziziensis- Brachiaria brizantha) en cabras de la provincia Artemisa. Rev. Salud Anim. 2016, 38, 19–24. Available online: http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S0253-570X2016000100004&lng=pt&nrm=iso&tlng=es (accessed on 15 November 2016).
- Brum, K.; Haraguchi, M.; Garutti, M.; Nóbrega, F.; Rosa, B.; Fioravanti, M.; Soares, J. Steroidal saponin concentrations in Brachiaria decumbens and B. brizantha at different developmental stages. Cienc. Rural 2009, 39, 279–281. Available online: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0103-84782009000100049 (accessed on 20 December 2013). [CrossRef]
- Castro, M.B.; Santos, H.L., Jr.; Mustafa, V.S.; Gracindo, C.V.; Moscardini, A.C.R.; Louvandini, H.; Paludo, G.R.; Borges, J.R.J.; Haraguchi, M.; Ferreira, M.B.; et al. Brachiaria spp. poisoning in sheep in Brazil: Experimental and epidemiological findings. In Poisoning by Plants, Mycotoxins, and Related Toxin, Proceedings of the 8th International Symposium on Poisonous Plants, João Pessoa, Para-Íba, Brazil, 2–8 May 2009; Riet-Correa, F., Pfister, J., Schild, A.L., Wierenga, T., Eds.; CABI: Oxfordshire, UK, 2011. [Google Scholar]
- Ferreira, M.; Brum, K.; Fernandes, C.; Martins, C.; Pinto, G.; Castro, V.; Rezende, K.G.; Riet-Correa, F.; Haraguchi, M.; Wysocki, H.L.; et al. Variation in saponin concentration in Brachiaria brizantha leaves as a function of maturation: Preliminary data. In Poisoning by Plants, Mycotoxins, and Related Toxin, Pcoceedings of the 8th International Symposium on Poisonous Plants, João Pessoa, Para-Íba, Brazil, 2–8 May 2009; Riet-Correa, F., Pfister, J., Schild, A.L., Wierenga, T., Eds.; CABI: Oxfordshire, UK, 2011. [Google Scholar]
- Sierra, J. Fundamentos Para el Establecimiento de Pasturas y Cultivos Forrajeros, 2nd ed.; Editorial Universidad de Antioquia: Medellín, Colombia, 2005; pp. 102–125. [Google Scholar]
- Lee, S.; Mitchell, R.; Gardner, D.; Tokarnia, C.; Riet-Correa, F. Measurement of steroidal saponins in Panicum and Brachiaria grasses in the USA and Brazil. In Poisoning by Plants, Mycotoxins, and Related Toxin, Proceedings of the 8th International Symposium on Poisonous Plants, João Pessoa, Para-Íba, Brazil, 2–8 May 2009; Riet-Correa, F., Pfister, J., Schild, A.L., Wierenga, T., Eds.; CABI: Oxfordshire, UK, 2011. [Google Scholar]
- Meagher, L.; Wilkins, A.; Miles, C.; Fagliari, J. Hepatogenous photosensitization of ruminants by Brachiaria decumbens and Panicum dichotomiflorum in the absence of sporidesmin: Lithogenic saponins may be responsible. Vet. Hum. Toxicol. 1996, 38, 271–274. [Google Scholar] [PubMed]
- Pires, V.S.; Taketa, A.T.C.; Gosmann, G.; Schenkel, E.P. Saponins and sapogenins from Brachiaria decumbens Stapf. J. Braz. Chem. Soc. 2002, 13, 135–139. [Google Scholar] [CrossRef]
- Low, S. Signal Grass (Brachiaria decumbens) Toxicity in Grazing Ruminants. Agriculture 2015, 5, 971–990. [Google Scholar] [CrossRef]
- Szakiel, A.; Pa̧czkowski, C.; Henry, M. Influence of environmental biotic factors on the content of saponins in plants. Phytochem. Rev. 2011, 10, 493–502. Available online: https://link.springer.com/article/10.1007%2Fs11101-010-9164-2 (accessed on 22 September 2016). [CrossRef]
- Giraldo, C.; Reyes, L.; Molina, J. Manejo Integrado de Artrópodos y Parásitos en Sistemas Silvopastoriles Intensivos; Manual 2, Proyecto Ganadería Colombiana Sostenible; GEF, BANCO MUNDIAL, FEDEGAN, CIPAV, FONDO ACCION, TNC: Bogotá, Colombia, 2011; pp. 8–30. Available online: http://www.cipav.org.co/pdf/2.Manejo.Integrado.de.Plagas.pdf (accessed on 10 September 2016).
- Dinchev, D.; Janda, B.; Evstatieva, L.; Oleszek, W.; Aslani, M.R.; Kostova, I. Distribution of steroidal saponins in Tribulus terrestris from different geographical regions. Phytochemistry 2008, 69, 176–186. Available online: http://www.sciencedirect.com/science/article/pii/S0031942207004657 (accessed on 5 October 2012). [CrossRef] [PubMed]
- Pavarini, D.P.; Pavarini, S.P.; Niehues, M.; Lopes, N.P. Exogenous influences on plant secondary metabolite levels. Anim. Feed Sci. Technol. 2012, 176, 5–16. Available online: http://www.sciencedirect.com/science/article/pii/S0377840112002350 (accessed on 10 September 2015). [CrossRef]
- Viloria de la Hoz, J. Geografía Económica de la Orinoquia; Banco de La República: Cartagena de Indias, Colombia, 2009; Available online: http://www.banrep.gov.co/publicaciones/pub_ec_reg4.htm (accessed on 8 October 2013).
- Gobbo-Neto, L.; Lopes, N.P. Plantas medicinais: Fatores de influência no conteúdo de metabólitos secundários fatores que influenciam o conteúdo de metabólitos secundários. Quim. Nova 2007, 31, 374–381. [Google Scholar] [CrossRef]
- Lima, F.G.; Lee, S.T.; Pfister, J.A.; Miyagi, E.S.; Costa, G.L.; Dias, R.; Fioravanti, M.C. The effect of ensiling and haymaking on the concentrations of steroidal saponin in two Brachiaria grass species. Cienc. Rural 2015, 45, 858–863. Available online: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0103-84782015000500858 (accessed on 30 July 2016). [CrossRef]
- Pulido, J.I.; Romero, M.; Rivero, S.T.; Duarte, O.A.; Gómez, P.J.; Vanegas, E.; Jaime, W.E.; Parra, J.L.; Pérez, R.A.; Cipagauta, M.; et al. Atlas de los Sistemas de Producción Bovina. Modulo Orinoquía y Amazonía. Plan de la Modernización Tecnológica de la Ganadería Bovina Colombiana; CORPOICA: Bogotá, Colombia, 2002; pp. 5–35. [Google Scholar]
- Rippstein, G.; Escobar, G.; Motta, F. Agroecología y Biodiversidad de las Sabanas en los Llanos Orientales de Colombia. Meta; CIAT: Cali, Colombia, 2001; pp. 1–22. [Google Scholar]
- Lima, F.G.; Haraguchi, M.; Pfister, J.A.; Guimaraes, V.Y.; Diogo, D.F. Weather and plant age affect the levels of steroidal saponin and Pithomyces chartarum spores in Brachiaria grass main forage source for ruminants. IJPPR 2012, 2, 45–53. Available online: https://www.ars.usda.gov/ARSUserFiles/oc/np/PoisonousPlants/Fall2012/grass.pdf (accessed on 15 April 2017).
- Lee, E.J.; Yoo, K.S.; Patil, B.S. Development of a Rapid HPLC-UV Method for Simultaneous Quantification of Protodioscin and Rutin in White and Green Asparagus Spears. J. Food Sci. 2010, 75, 703–709. Available online: http://onlinelibrary.wiley.com/doi/10.1111/j.1750-3841.2010.01824.x/pdf (accessed on 18 July 2016). [CrossRef] [PubMed]
- EPA (Environmental Protection Agency). 40 CFR Appendix B to Part 136—Definition and Procedure for the Determination of the Method Detection Limit-Revision 1.11; In EPA Code of Federal Regulations; USA Government Printing Office: WA, USA, 2011; pp. 345–347.
- Martínez, R.; Martínez, N.; Martínez, M.V. Diseño de Experimentos en Ciencias Agropecuarias y Biológicas con SAS, SPSS, R Y STATISTIX; Fondo Nacional Universitario: Bogotá, Colombia, 2011; Volume 1, pp. 73–125. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protodioscin Concentration (%) | Brachiaria decumbens | Hybrid * | Brachiaria brizantha | Total | |||
|---|---|---|---|---|---|---|---|
| V | P | V | P | V | P | ||
| >0 to ≤1 | 0% n = 0 | 4% n = 6 | 1% n = 2 | 1% n = 2 | 8% n = 11 | 4% n = 5 | 23% n = 33 |
| >1 to ≤2 | 4% n = 6 | 5% n = 7 | 8% n = 12 | 14% n = 20 | 8% n = 12 | 12% n = 17 | 51% n = 74 |
| >2 to ≤3 | 11% n = 16 | 6% n = 8 | 3% n = 5 | 1% n = 1 | 0% n = 0 | 0% n = 0 | 21% n = 30 |
| >3 | 0% n = 0 | 1% n = 2 | 4% n = 5 | 0% n = 0 | 0% n = 0 | 0% n = 0 | 5% n = 7 |
| Sampling | Day | P (mm) | T (°C) | RH(%) | SB(hours) | ||||
|---|---|---|---|---|---|---|---|---|---|
| P | V | P | V | P | V | P | V | ||
| April 2012 Initial rains | 14 | 130.4 | 183.8 | 25.7 | 24.5 | 85 | 90 | 76.0 | 37.1 |
| 28 | 367.0 | 276.9 | 25.3 | 24.9 | 84 | 88 | 133.5 | 91.8 | |
| June 2012 Advanced rains | 14 | 176.0 | 112.7 | 24.9 | 24.5 | 84 | 87 | 55.5 | 58.7 |
| 28 | 364.9 | 203.5 | 24.7 | 24.5 | 83 | 87 | 136.4 | 128.5 | |
| October 2012 Light rains | 14 | 129.5 | 159.0 | 26.0 | 25.2 | 82 | 87 | 51.3 | 60.2 |
| 28 | 286.6 | 244.8 | 26.3 | 25.5 | 80 | 85 | 159.1 | 131.6 | |
| February 2013 Droughty | 14 | 9.4 | 23.9 | 28.2 | 25.2 | 72 | 87 | 70.8 | 74.8 |
| 28 | 35.6 | 57.2 | 27.7 | 25.5 | 73 | 85 | 116.8 | 122.9 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozano, M.C.; Martinez, N.M.; Diaz, G.J. Content of the Saponin Protodioscin in Brachiaria spp. from the Eastern Plains of Colombia. Toxins 2017, 9, 220. https://doi.org/10.3390/toxins9070220
Lozano MC, Martinez NM, Diaz GJ. Content of the Saponin Protodioscin in Brachiaria spp. from the Eastern Plains of Colombia. Toxins. 2017; 9(7):220. https://doi.org/10.3390/toxins9070220
Chicago/Turabian StyleLozano, Maria C., Nhora M. Martinez, and Gonzalo J. Diaz. 2017. "Content of the Saponin Protodioscin in Brachiaria spp. from the Eastern Plains of Colombia" Toxins 9, no. 7: 220. https://doi.org/10.3390/toxins9070220
APA StyleLozano, M. C., Martinez, N. M., & Diaz, G. J. (2017). Content of the Saponin Protodioscin in Brachiaria spp. from the Eastern Plains of Colombia. Toxins, 9(7), 220. https://doi.org/10.3390/toxins9070220