Assembly and Connection of Micropatterned Single Neurons for Neuronal Network Formation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
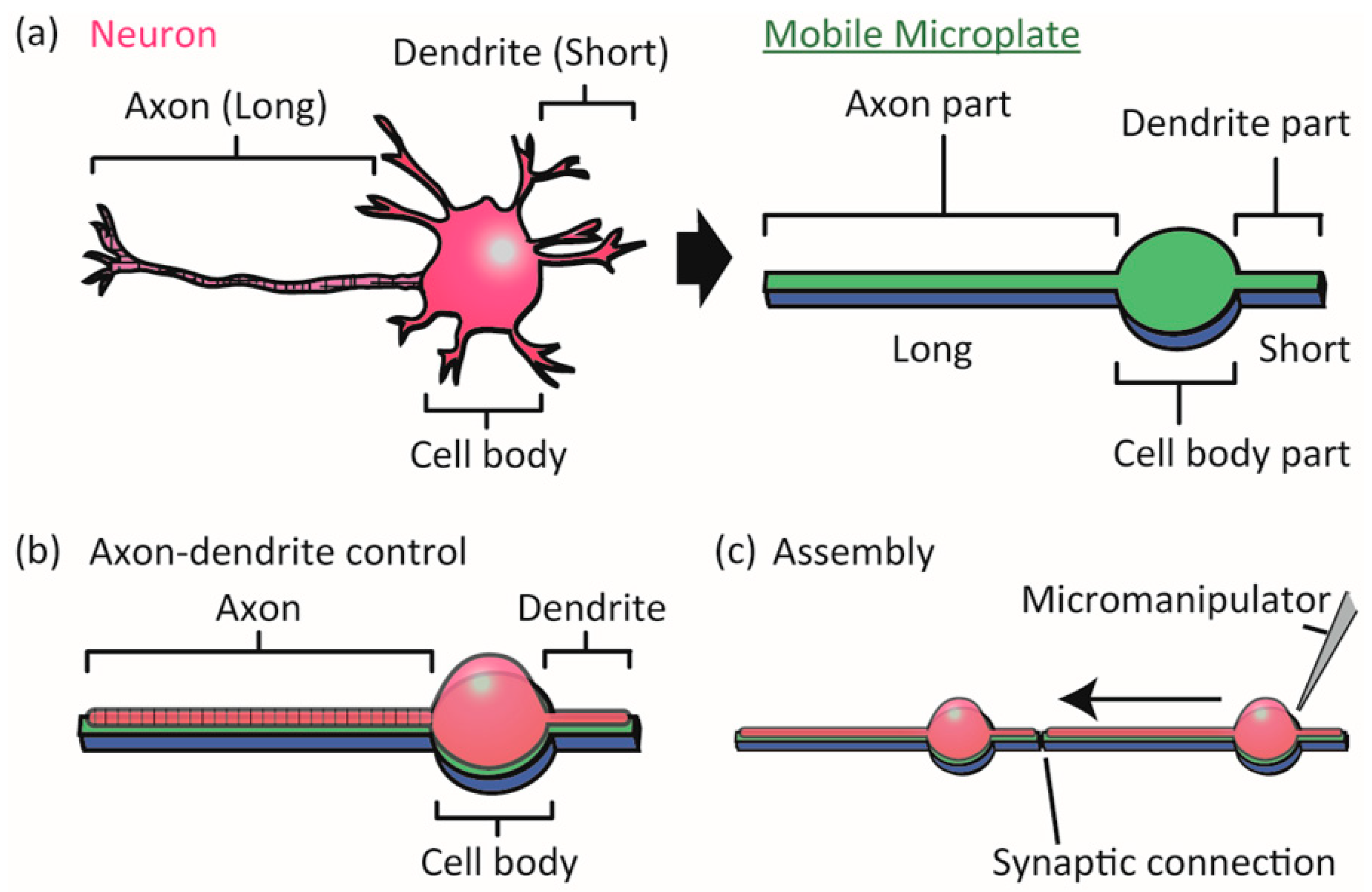
2.1. Design of Mobile Microplates
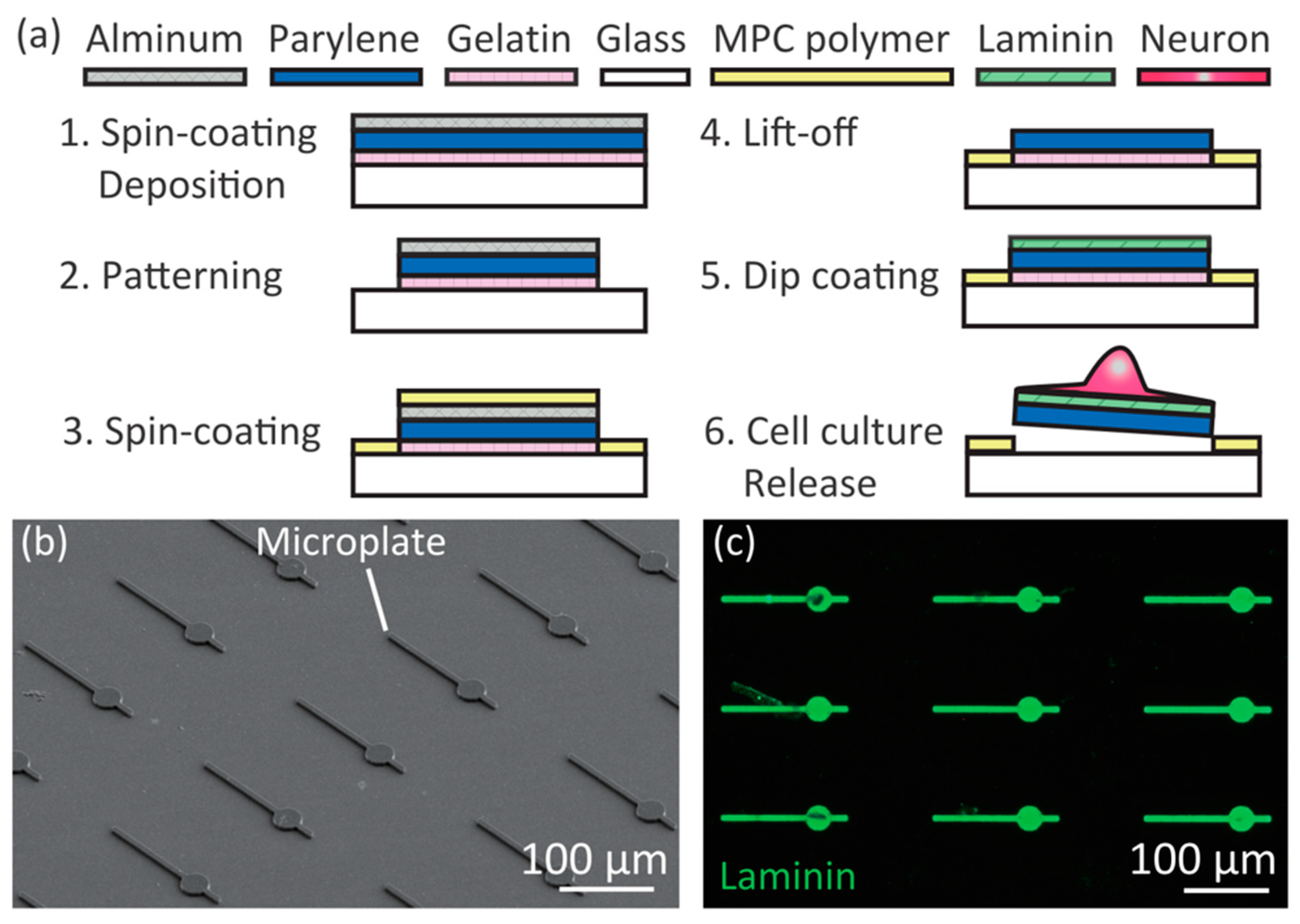
2.2. Fabrication of Mobile Microplates
2.3. Cell Culture
2.4. Gene Manipulation
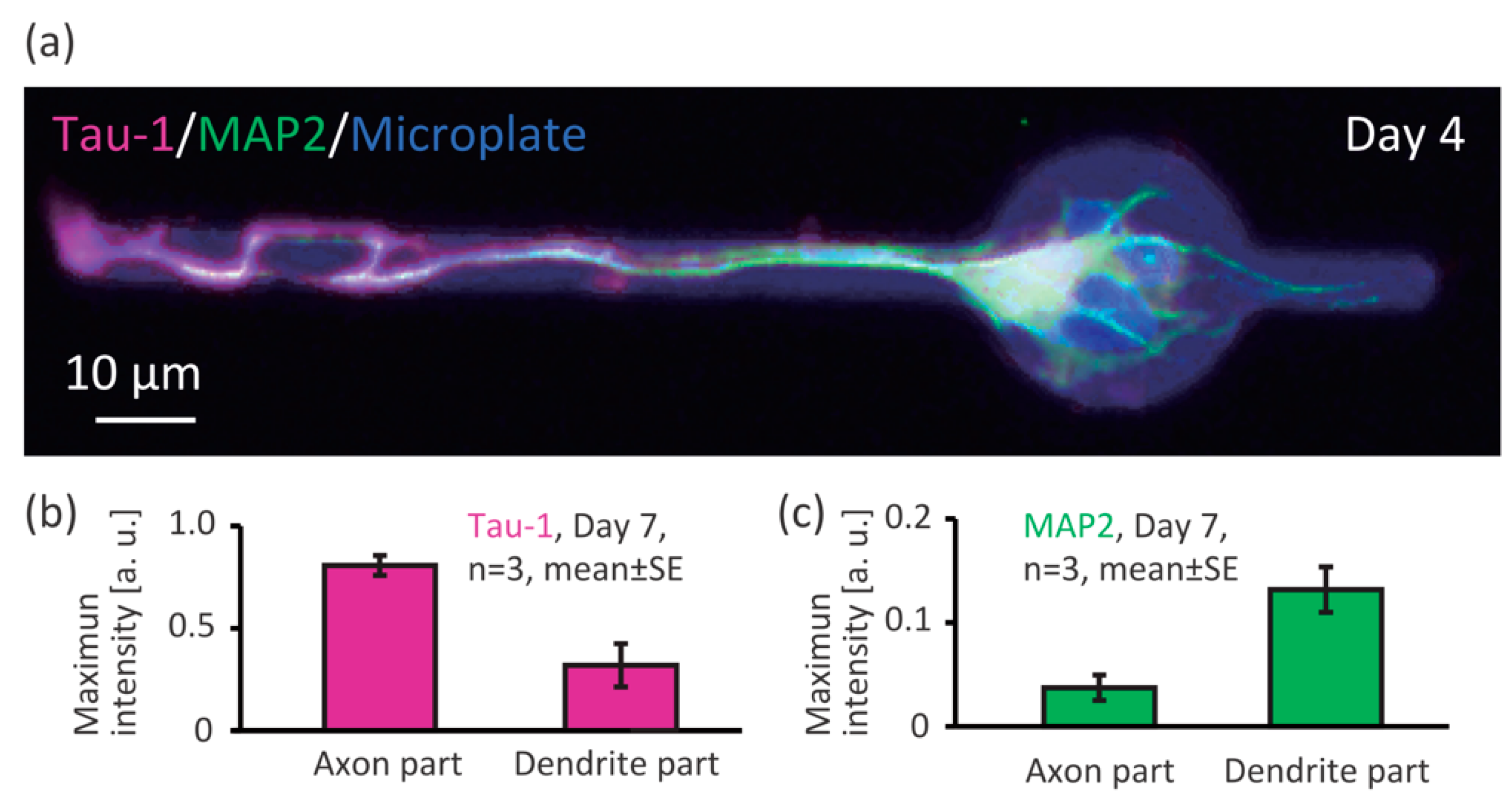
2.5. Immunostaining of Neurons on Microplates
2.6. SEM Imaging
2.7. Image Acquisition and Processing
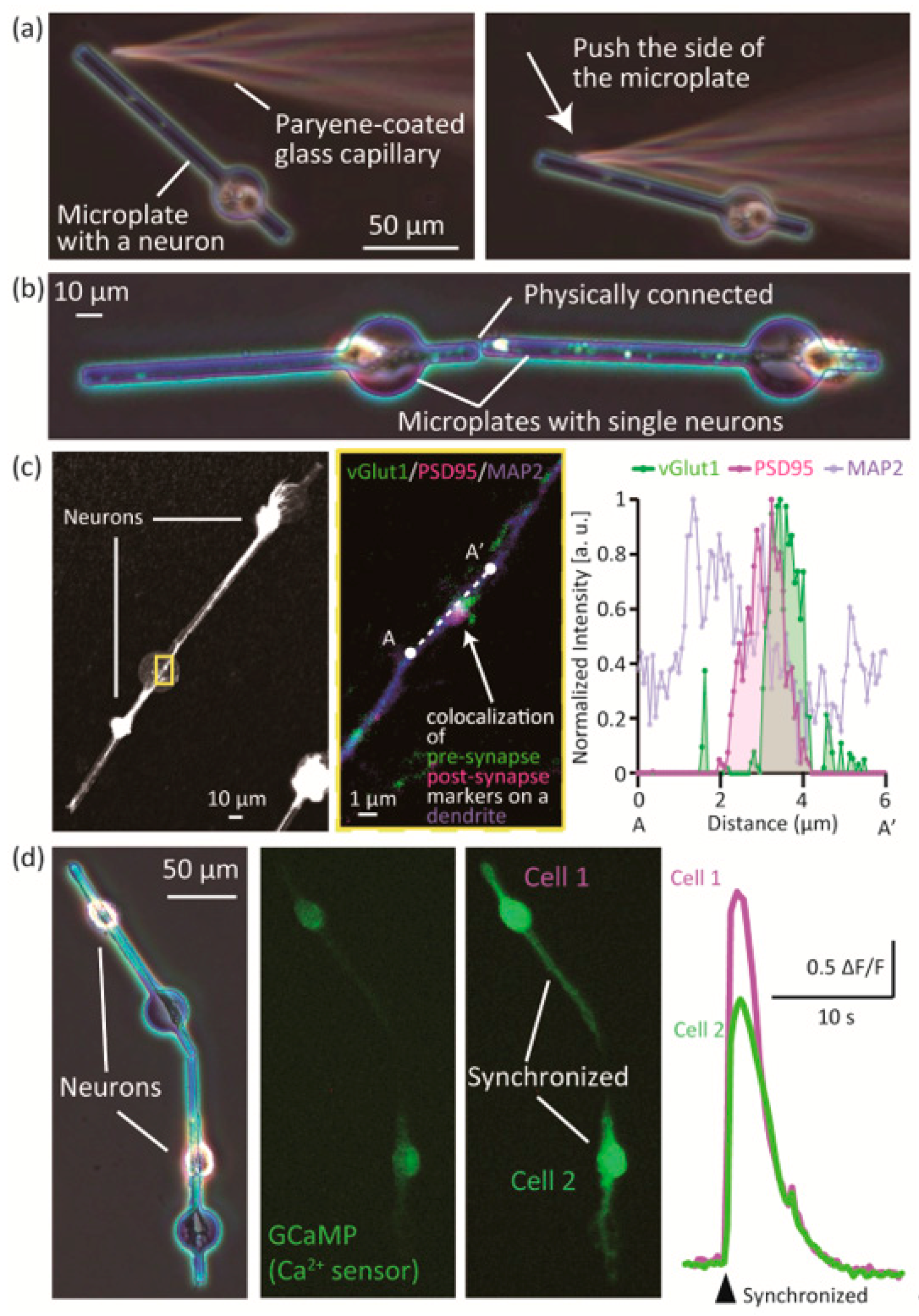
2.8. Handling of Single Neurons on Microplates
3. Results
3.1. Controllability of Position of Cell Body/Axon/Dendrites on Microplates
3.2. Assembly and Connection of Neurons on the Microplates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kandel, E.R.; Schwartz, J.H.; Jessell, T.M.; Siegelbaum, S.A.; Hudspeth, A.J. Principles of Neural Science, 5th ed.; McGraw-Hill Education/Medical: New York, NY, USA, 2012. [Google Scholar]
- Bock, D.D.; Lee, W.C.; Kerlin, A.M.; Andermann, M.L.; Hood, G.; Wetzel, A.W.; Yurgenson, S.; Soucy, E.R.; Kim, H.S.; Reid, R.C. Network anatomy and in vivo physiology of visual cortical neurons. Nature 2011, 471, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Dan, Y.; Poo, M.M. Spike timing-dependent plasticity: From synapse to perception. Physiol. Rev. 2006, 86, 1033–1048. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, B.C.; Brewer, G.J. Designing Neural Network in Culture. Proc. IEEE Inst. Electr. Electron. Eng. 2010, 98, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Simitzi, C.; Ranella, A.; Stratakis, E. Controlling the morphology and outgrowth of nerve and neuroglial cells: The effect of surface topography. Acta Biomater. 2017, 51, 21–52. [Google Scholar] [CrossRef] [PubMed]
- Simitzi, C.; Karali, K.; Ranella, A.; Stratakis, E. Controlling the outgrowth and functions of neural stem cells: The effect of surface topography. ChemPhysChem 2018. [Google Scholar] [CrossRef] [PubMed]
- Corey, J.M.; Wheeler, B.C.; Brewer, G.J. Compliance of hippocampal neurons to patterned substrate networks. J. Neurosci. Res. 1991, 30, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.J.; Nam, Y. Geometric effect of cell adhesive polygonal micropatterns on neuritogenesis and axon guidance. J. Neural Eng. 2012, 9, 046019. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.R.; Jang, M.J.; Joo, S.; Sun, W.; Nam, Y. Surface-printed microdot array chips for the quantification of axonal collateral branching of a single neuron in vitro. Lab Chip 2014, 14, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Albers, J.; Toma, K.; Offenhäusser, A. Engineering connectivity by multiscale micropatterning of individual populations of neurons. Biotechnol. J. 2015, 10, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Kunze, A.; Tseng, P.; Godzich, C.; Murray, C.; Caputo, A.; Schweizer, F.E.; Di Carlo, D. Engineering cortical neuron polarity with nanomagnets on a chip. ACS Nano 2015, 9, 3664–3676. [Google Scholar] [CrossRef] [PubMed]
- Su, W.-T.; Liao, Y.-F.; Wu, T.-W.; Wang, B.-J.; Shih, Y.-Y. Microgrooved patterns enhanced PC12 cell growth, orientation, neurite elongation, and neuritogenesis. J. Biomed. Mater. Res. Part A 2013, 101, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Unsworth, C.P.; Holloway, H.; Delivopoulos, E.; Murray, A.F.; Simpson, M.C.; Dickinson, M.E.; Graham, E.S. Biomaterials Patterning and detailed study of human hNT astrocytes on parylene-C/silicon dioxide substrates to the single cell level. Biomaterials 2011, 32, 6541–6550. [Google Scholar] [CrossRef] [PubMed]
- Aebersold, M.J.; Dermutz, H.; Forró, C.; Weydert, S.; Thompson-Stecke, G.; Vörös, J.; Demkó, L. “Brains on a chip”: Towards engineered neural networks. TrAC-Trends Anal. Chem. 2016, 78, 60–69. [Google Scholar] [CrossRef]
- Fricke, R.; Zentis, P.D.; Rajappa, L.T.; Hofmann, B.; Banzet, M.; Offenhausser, A.; Meffert, S.H. Axon guidance of rat cortical neurons by microcontact printed gradients. Biomaterials 2011, 32, 2070–2076. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.; Bisbal, M.; Brocard, J.; Bugnicourt, G.; Saoudi, Y.; Andrieux, A.; Gory-Faure, S.; Villard, C. How morphological constraints affect axonal polarity in mouse neurons. PLoS ONE 2012, 7, e33623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, A.K.; Wrobel, G.; Meyer, W.; Knoll, W.; Offenhausser, A. Synaptic plasticity in micropatterned neuronal networks. Biomaterials 2005, 26, 2549–2557. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, I.; Sugio, Y.; Jimbo, Y.; Yasuda, K. Stepwise pattern modification of neuronal network in photo-thermally-etched agarose architecture on multi-electrode array chip for individual-cell-based electrophysiological measurement. Lab Chip 2005, 5, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.; Bugnicourt, G.; Bisbal, M.; Gory-Faure, S.; Brocard, J.; Villard, C. Neuronal architectures with axo-dendritic polarity above silicon nanowires. Small 2012, 8, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Demura, T.; Morita, M.; Banker, G.A.; Tanii, T.; Nakamura, S. Differential neurite outgrowth is required for axon specification by cultured hippocampal neurons. J. Neurochem. 2012, 123, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Boehler, M.D.; Leondopulos, S.S.; Wheeler, B.C.; Brewer, G.J. Hippocampal networks on reliable patterned substrates. J. Neurosci. Methods 2012, 203, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Teshima, T.; Kuribayashi-Shigetomi, K.; Takeuchi, S. Mobile microplates for morphological control and assembly of individual neural cells. Adv. Healthc. Mater. 2016, 5, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Sibarani, J.; Takai, M.; Ishihara, K. Surface modification on microfluidic devices with 2-methacryloyloxyethyl phosphorylcholine polymers for reducing unfavorable protein adsorption. Coll. Surf. B 2007, 54, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Kato-Negishi, M.; Onoe, H.; Takeuchi, S. Rod-shaped Neural Units for Aligned 3D Neural Network Connection. Adv. Healthc. Mater. 2017, 6, 1700143. [Google Scholar] [CrossRef] [PubMed]
- Hümmer, D.; Kurth, F.; Naredi-Rainer, N.; Dittrich, P.S. Single cells in confined volumes: Microchambers and microdroplets. Lab Chip 2016, 16, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Le Roux, P.D.; Reh, T.A. Regional differences in glial-derived factors that promote dendritic outgrowth from mouse cortical neurons in vitro. J. Neurosci. 1994, 14, 4639–4655. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshida, S.; Kato-Negishi, M.; Takeuchi, S. Assembly and Connection of Micropatterned Single Neurons for Neuronal Network Formation. Micromachines 2018, 9, 235. https://doi.org/10.3390/mi9050235
Yoshida S, Kato-Negishi M, Takeuchi S. Assembly and Connection of Micropatterned Single Neurons for Neuronal Network Formation. Micromachines. 2018; 9(5):235. https://doi.org/10.3390/mi9050235
Chicago/Turabian StyleYoshida, Shotaro, Midori Kato-Negishi, and Shoji Takeuchi. 2018. "Assembly and Connection of Micropatterned Single Neurons for Neuronal Network Formation" Micromachines 9, no. 5: 235. https://doi.org/10.3390/mi9050235
APA StyleYoshida, S., Kato-Negishi, M., & Takeuchi, S. (2018). Assembly and Connection of Micropatterned Single Neurons for Neuronal Network Formation. Micromachines, 9(5), 235. https://doi.org/10.3390/mi9050235