Immunomagnetic-Enriched Subpopulations of Melanoma Circulating Tumour Cells (CTCs) Exhibit Distinct Transcriptome Profiles


 ,
,  ,
,
Abstract
:1. Background
2. Results
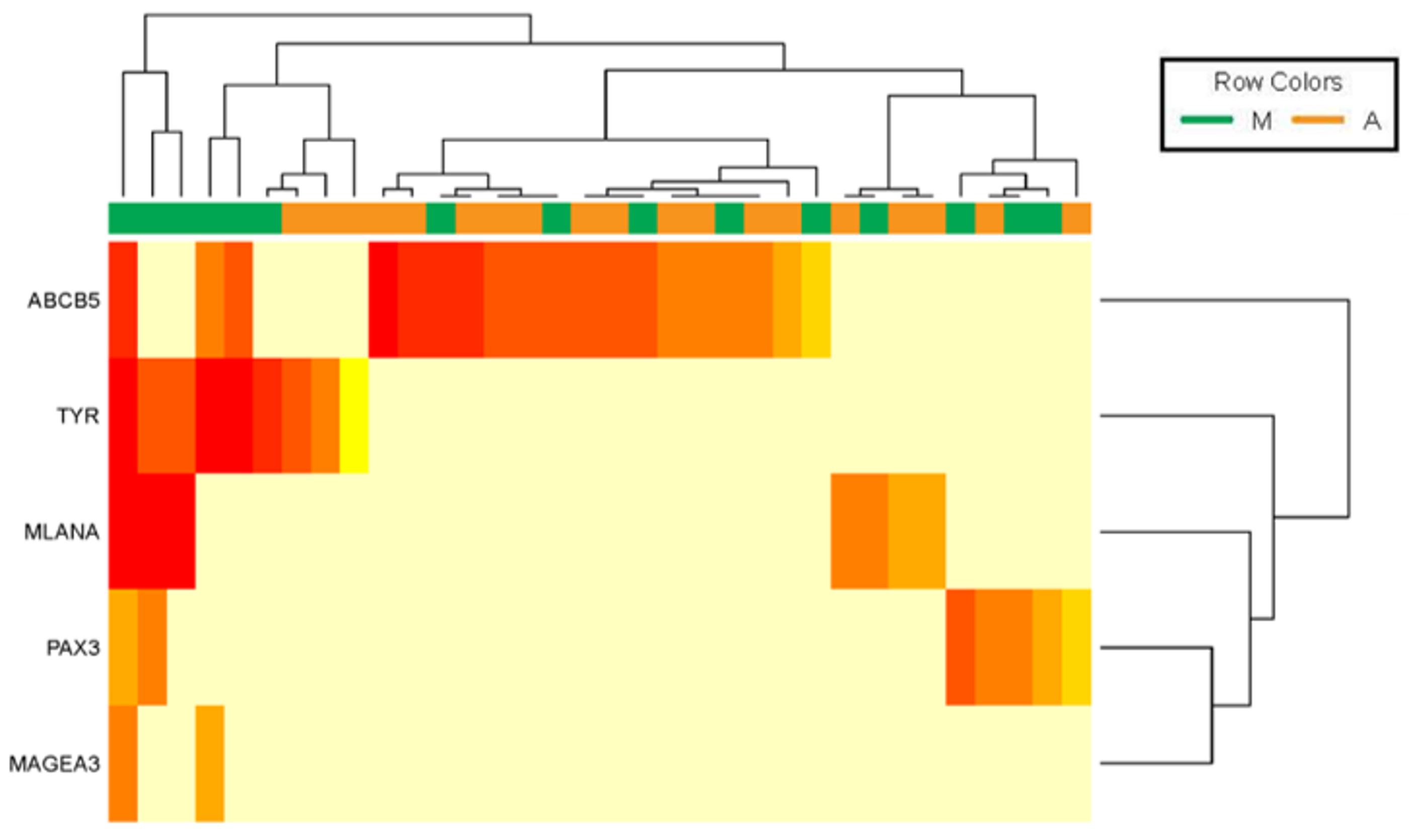
2.1. Expression of Five Melanoma-Specific Transcripts in Immunomagnetically Enriched ABCB5 and MCSP CTC Fractions from Metastatic Melanoma Patients
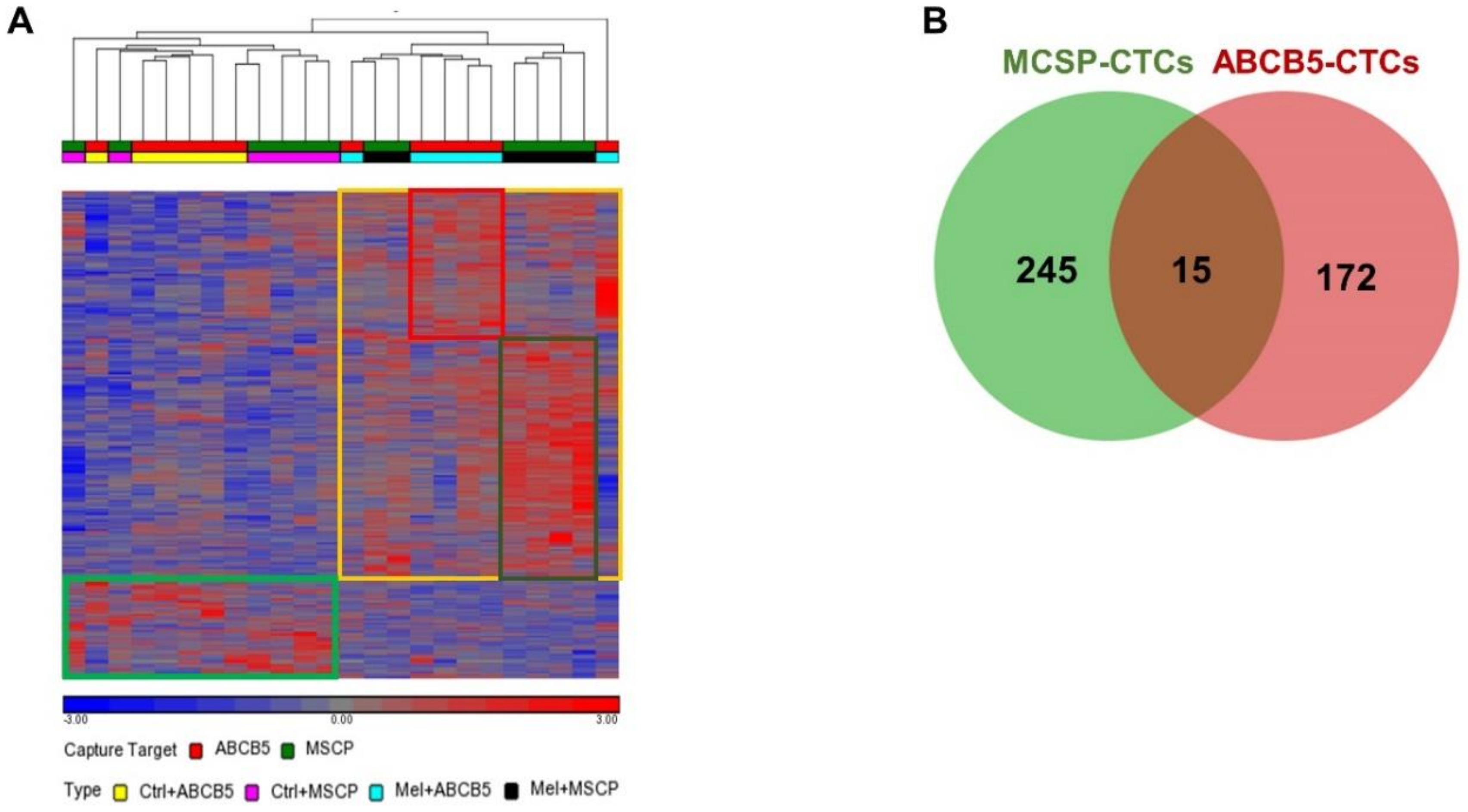
2.2. MCSP and ABCB5 CTC Subpopulations Exhibit Distinct Gene Expression Profiles
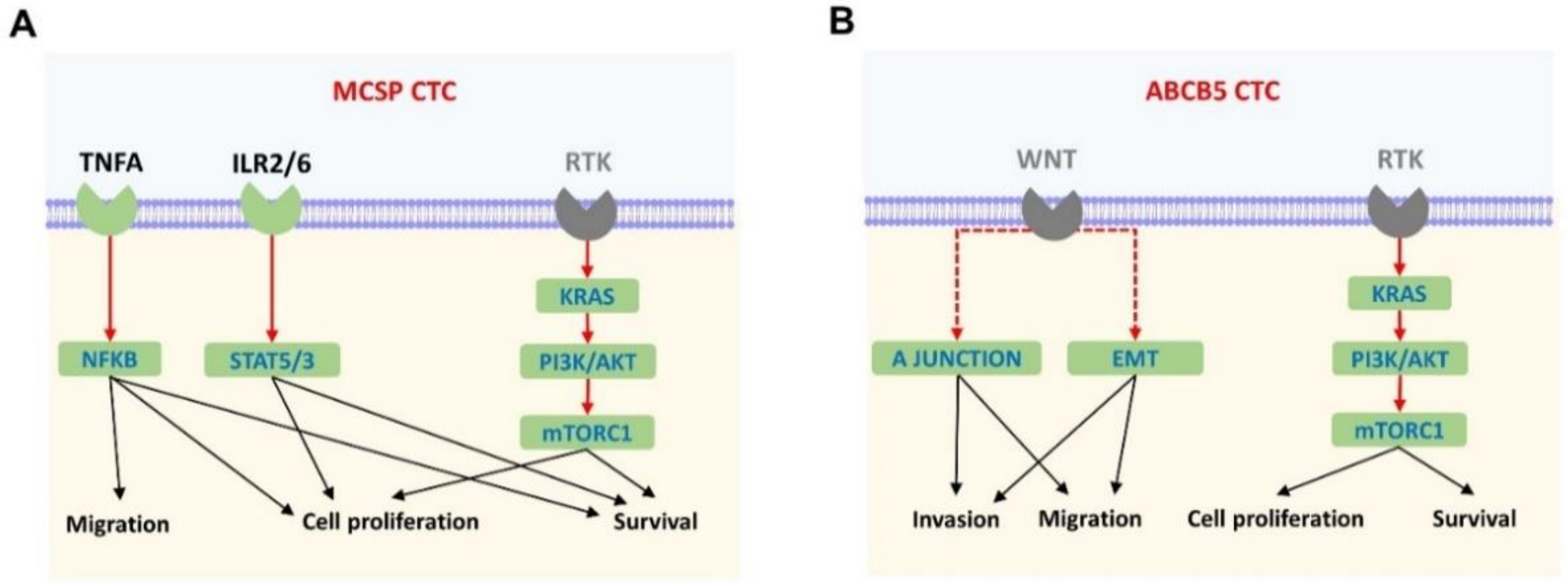
2.3. Tumour Necrosis Factor Alpha (TNFA) Signalling and Epithelial Mesenchymal Transition (EMT) Are Significantly Enriched in the MCSP and ABCB5 CTC Fractions, Respectively
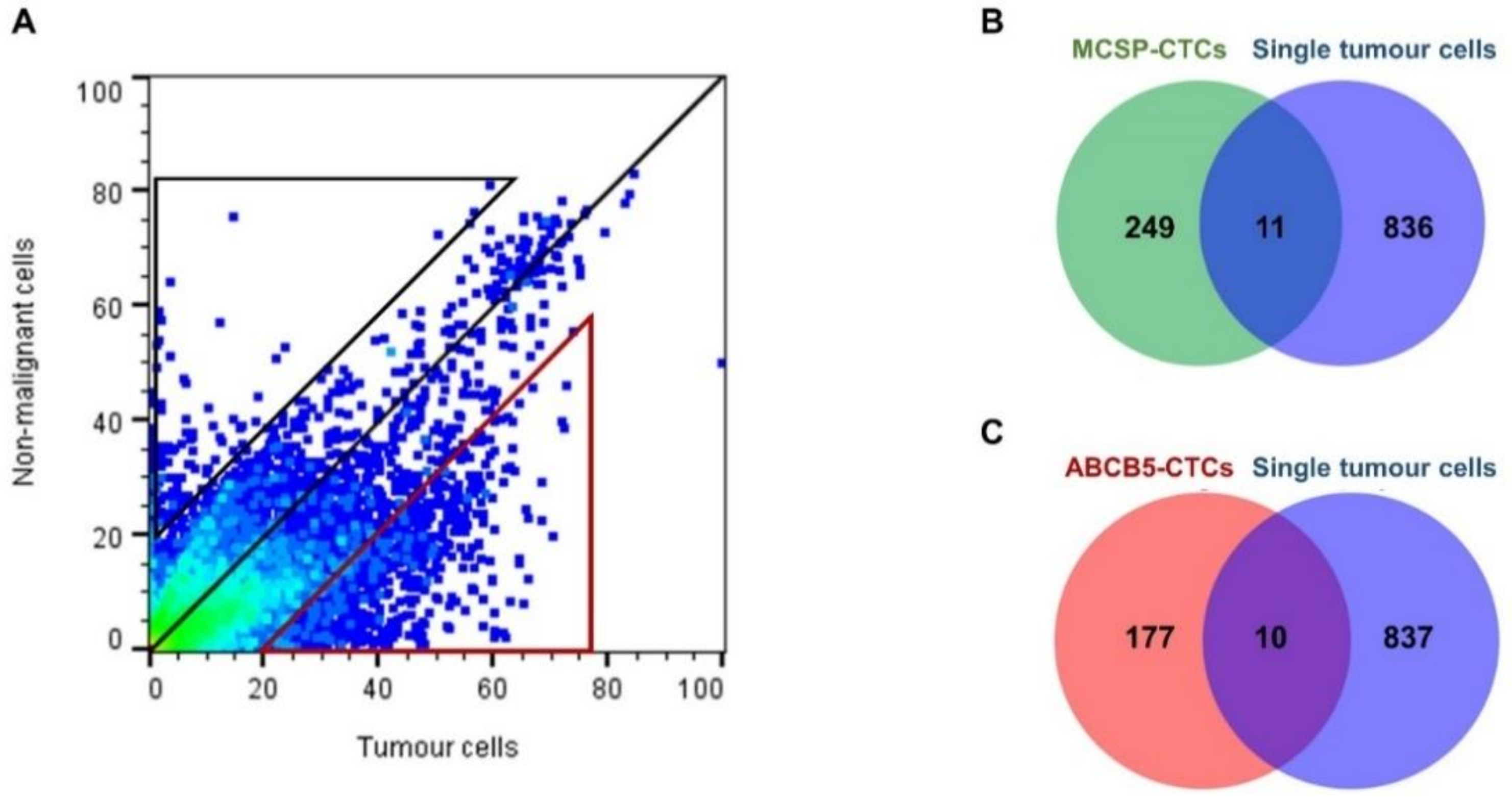
2.4. In Silico Analysis Identifies Melanoma Cell-Associated Genes in the Gene Expression Profiles of the MCSP- and ABCB5-Enriched CTC Fractions
3. Discussion
4. Methods
4.1. Patient Recruitment and Blood Collection
4.2. CTC Enrichment and Nucleic Acid Extraction
4.3. RT-PCR
4.4. Microarray
4.5. Microarray Data Analysis
4.6. Gene Set Enrichment Analysis (GSEA)
4.7. Identification of Tumour-Derived Genes in the MCSP- and ABCB5-Enriched Fractions
5. Conclusions
Supplementary Materials
Data Availability
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sharma, P.; Hu-Lieskovan, S.; Wargo, J.A.; Ribas, A. Primary, Adaptive, and Acquired Resistance to Cancer Immunotherapy. Cell 2017, 168, 707–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roesch, A. Tumor heterogeneity and plasticity as elusive drivers for resistance to MAPK pathway inhibition in melanoma. Oncogene 2015, 34, 2951–2957. [Google Scholar] [CrossRef] [PubMed]
- Dive, C.; Brady, G. SnapShot: Circulating Tumor Cells. Cell 2017, 168, 742. [Google Scholar] [CrossRef] [PubMed]
- Micalizzi, D.S.; Maheswaran, S.; Haber, D.A. A conduit to metastasis: circulating tumor cell biology. Genes Dev. 2017, 31, 1827–1840. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.S.; Reid, A.; Bowyer, S.; Calapre, L.; Siew, K.; Pearce, R.; Cowell, L.; Frank, M.H.; Millward, M.; Ziman, M. Circulating Melanoma Cell Subpopulations: Their Heterogeneity and Differential Responses to Treatment. J. Investig. Dermatol. 2015, 135, 2040–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khattak, M.; Gray, E.S.; Freeman, J.; Pereira, M.; Meniawy, T.; Siew, K.; Millward, M.; Ziman, M. PD-L1 expression in Circulating Melanoma Cells is predictive of response to Pembrolizumab. In Proceedings of the Society for Melanoma Research, Boston, MA, USA, 29 October 2016. [Google Scholar]
- Khoja, L.; Lorigan, P.; Zhou, C.; Lancashire, M.; Booth, J.; Cummings, J.; Califano, R.; Clack, G.; Hughes, A.; Dive, C. Biomarker utility of circulating tumor cells in metastatic cutaneous melanoma. J. Investig. Dermatol. 2013, 133, 1582–1590. [Google Scholar] [CrossRef] [PubMed]
- Klinac, D.; Gray, E.S.; Freeman, J.B.; Reid, A.; Bowyer, S.; Millward, M.; Ziman, M. Monitoring changes in circulating tumour cells as a prognostic indicator of overall survival and treatment response in patients with metastatic melanoma. BMC Cancer 2014, 14, 423. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Mitra, D.; Sullivan, R.J.; Wittner, B.S.; Kimura, A.M.; Pan, S.; Hoang, M.P.; Brannigan, B.W.; Lawrence, D.P.; Flaherty, K.T.; et al. Isolation and Molecular Characterization of Circulating Melanoma Cells. Cell Rep. 2014, 7, 645–653. [Google Scholar] [CrossRef] [Green Version]
- Freeman, J.B.; Gray, E.S.; Millward, M.; Pearce, R.; Ziman, M. Evaluation of a multi-marker immunomagnetic enrichment assay for the quantification of circulating melanoma cells. J. Transl. Med. 2012, 10, 192. [Google Scholar] [CrossRef] [Green Version]
- Khoja, L.; Shenjere, P.; Hodgson, C.; Hodgetts, J.; Clack, G.; Hughes, A.; Lorigan, P.; Dive, C. Prevalence and heterogeneity of circulating tumour cells in metastatic cutaneous melanoma. Melanoma Res. 2014, 24, 40–46. [Google Scholar] [CrossRef]
- Aya-Bonilla, C.; AG, M.; Freeman, J.; Lomme, C.; Khattak, M.; Meniawy, T.; Millward, M.; Warkiani, M.E.; Gray, E.S.; Ziman, M. Isolation and detection of circulating tumour cells from metastatic melanoma patients using a slanted spiral microfluidic device. Oncotarget 2017, 8, 67355–67368. [Google Scholar] [CrossRef]
- Marsavela, A.G.; Aya Bonilla, C.A.; Warkiani, M.E.; Gray, E.S.; Ziman, M. Melanoma Circulating Tumor Cells: Benefits and Challenges Required for Clinical Application. Cancer Lett. 2018, 424, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Trzpis, M.; McLaughlin, P.M.J.; de Leij, L.M.F.H.; Harmsen, M.C. Epithelial cell adhesion molecule: More than a carcinoma marker and adhesion molecule. Am. J. Pathol. 2007, 171, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Cristofanilli, M.; Budd, G.T.; Ellis, M.J.; Stopeck, A.; Matera, J.; Miller, M.C.; Reuben, J.M.; Doyle, G.V.; Allard, W.J.; Terstappen, L.W.M.M.; et al. Circulating Tumor Cells, Disease Progression, and Survival in Metastatic Breast Cancer. New Eng. J. Med. 2004, 351, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.; Wu, E.; Weiss, L.M. Cytokeratin 7 and Cytokeratin 20 Expression in Epithelial Neoplasms: A Survey of 435 Cases. Mod. Pathol. 2000, 13, 962. [Google Scholar] [CrossRef] [PubMed]
- Momburg, F.; Moldenhauer, G.; Hämmerling, G.J.; Möller, P. Immunohistochemical Study of the Expression of a Mr 34,000 Human Epithelium-specific Surface Glycoprotein in Normal and Malignant Tissues. Cancer Res. 1987, 47, 2883–2891. [Google Scholar] [PubMed]
- Korabiowska, M.; Fischer, G.; Steinacker, A.; Stachura, J.; Cordon-Cardo, C.; Brinck, U. Cytokeratin Positivity in Paraffin-embedded Malignant Melanomas: Comparative Study of KL1, A4 and Lu5 Antibodies. Anticancer Res. 2004, 24, 3203–3208. [Google Scholar]
- Maguire, L.H.; Thomas, A.R.; Goldstein, A.M. Tumors of the neural crest: Common themes in development and cancer. Dev. Dyn. 2015, 244, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Mort, R.L.; Jackson, I.J.; Patton, E.E. The melanocyte lineage in development and disease. Development 2015, 142, 620–632. [Google Scholar] [CrossRef] [Green Version]
- Yanagita, M.; Luke, J.J.; Hodi, F.S.; Janne, P.A.; Paweletz, C.P. Isolation and characterization of circulating melanoma cells by size filtration and fluorescent in situ hybridization. Melanoma Res. 2018, 28, 89–95. [Google Scholar] [CrossRef]
- Ulmer, A.; Schmidt-Kittler, O.; Fischer, J.; Ellwanger, U.; Rassner, G.; Riethmuller, G.; Fierlbeck, G.; Klein, C.A. Immunomagnetic enrichment, genomic characterization, and prognostic impact of circulating melanoma cells. Clin. Cancer Res. 2004, 10, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Faye, R.S.; Aamdal, S.; Hoifodt, H.K.; Jacobsen, E.; Holstad, L.; Skovlund, E.; Fodstad, O. Immunomagnetic detection and clinical significance of micrometastatic tumor cells in malignant melanoma patients. Clin. Cancer Res. 2004, 10, 4134–4139. [Google Scholar] [CrossRef] [PubMed]
- Ramsköld, D.; Luo, S.; Wang, Y.-C.; Li, R.; Deng, Q.; Faridani, O.R.; Daniels, G.A.; Khrebtukova, I.; Loring, J.F.; Laurent, L.C.; et al. Full-Length mRNA-Seq from single cell levels of RNA and individual circulating tumor cells. Nat. Biotechnol. 2012, 30, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Campoli, M.R.; Chang, C.-C.; Kageshita, T.; Wang, X.; McCarthy, J.B.; Ferrone, S. Human high molecular weight-melanoma-associated antigen (HMW-MAA): A melanoma cell surface chondroitin sulfate proteoglycan (MSCP) with biological and clinical significance. Crit. Rev. Immunol. 2004, 24, 267. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, M.; Rybczynska, A.A.; Wei, Y.; Schwenkert, M.; Fey, G.H.; Dierckx, R.A.; van Waarde, A.; Helfrich, W.; Bremer, E. Melanoma-associated Chondroitin Sulfate Proteoglycan (MCSP)-targeted delivery of soluble TRAIL potently inhibits melanoma outgrowth in vitro and in vivo. Mol. Cancer 2010, 9, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chartrain, M.; Riond, J.; Stennevin, A.; Vandenberghe, I.; Gomes, B.; Lamant, L.; Meyer, N.; Gairin, J.E.; Guilbaud, N.; Annereau, J.P. Melanoma Chemotherapy Leads to the Selection of ABCB5-Expressing Cells. PLoS ONE 2012, 7, e36762. [Google Scholar] [CrossRef] [PubMed]
- Schatton, T.; Murphy, G.F.; Frank, N.Y.; Yamaura, K.; Waaga-Gasser, A.M.; Gasser, M.; Zhan, Q.; Jordan, S.; Duncan, L.M.; Weishaupt, C.; et al. Identification of cells initiating human melanomas. Nature 2008, 451, 345–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, B.J.; Saab, K.R.; Ma, J.; Schatton, T.; Pütz, P.; Zhan, Q.; Murphy, G.F.; Gasser, M.; Waaga-Gasser, A.M.; Frank, N.Y.; et al. ABCB5 Maintains Melanoma-Initiating Cells through a Proinflammatory Cytokine Signaling Circuit. Cancer Res. 2014, 74, 4196–4207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupas, V.; Weishaupt, C.; Siepmann, D.; Kaserer, M.-L.; Eickelmann, M.; Metze, D.; Luger, T.A.; Beissert, S.; Loser, K. RANK Is Expressed in Metastatic Melanoma and Highly Upregulated on Melanoma-Initiating Cells. J. Investig. Dermatol. 2010, 131, 944–955. [Google Scholar] [CrossRef] [PubMed]
- Tirosh, I.; Izar, B.; Prakadan, S.M.; Wadsworth, M.H., 2nd; Treacy, D.; Trombetta, J.J.; Rotem, A.; Rodman, C.; Lian, C.; Murphy, G.; et al. Dissecting the multicellular ecosystem of metastatic melanoma by single-cell RNA-seq. Science 2016, 352, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Reid, A.L.; Freeman, J.B.; Millward, M.; Ziman, M.; Gray, E.S. Detection of BRAF-V600E and V600K in melanoma circulating tumour cells by droplet digital PCR. Clin. Bio. 2015, 48, 999–1002. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Sullivan, R.J.; Kalinich, M.; Kwan, T.T.; Giobbie-Hurder, A.; Pan, S.; LiCausi, J.A.; Milner, J.D.; Nieman, L.T.; Wittner, B.S.; et al. Molecular signatures of circulating melanoma cells for monitoring early response to immune checkpoint therapy. PNAS 2018. [Google Scholar] [CrossRef]
- Eisenmann, K.M.; McCarthy, J.B.; Simpson, M.A.; Keely, P.J.; Guan, J.-L.; Tachibana, K.; Lim, L.; Manser, E.; Furcht, L.T.; Iida, J. Melanoma chondroitin sulphate proteoglycan regulates cell spreading through Cdc42, Ack-1 and p130cas. Nat. Cell Biol. 1999, 1, 507. [Google Scholar] [CrossRef] [PubMed]
- Iida, J.; Wilhelmson, K.L.; Ng, J.; Lee, P.; Morrison, C.; Tam, E.; Overall, C.M.; McCarthy, J.B. Cell surface chondroitin sulfate glycosaminoglycan in melanoma: role in the activation of pro-MMP-2 (pro-gelatinase A). Biochem. J. 2007, 403, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Price, M.A.; Li, G.Y.; Bar-Eli, M.; Salgia, R.; Jagedeeswaran, R.; Carlson, J.H.; Ferrone, S.; Turley, E.A.; McCarthy, J.B. Melanoma Proteoglycan Modifies Gene Expression to Stimulate Tumor Cell Motility, Growth, and Epithelial-to-Mesenchymal Transition. Cancer Res. 2009, 69, 7538–7547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Price, M.A.; Neudauer, C.L.; Wilson, C.; Ferrone, S.; Xia, H.; Iida, J.; Simpson, M.A.; McCarthy, J.B. Melanoma chondroitin sulfate proteoglycan enhances FAK and ERK activation by distinct mechanisms. J. Cell Biol. 2004, 165, 881–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royer, C.; Lu, X. Epithelial cell polarity: A major gatekeeper against cancer? Cell Death Differ. 2011, 18, 1470–1477. [Google Scholar] [CrossRef]
- Pearlman, R.L.; Montes de Oca, M.K.; Pal, H.C.; Afaq, F. Potential therapeutic targets of epithelial–mesenchymal transition in melanoma. Cancer Lett. 2017, 391, 125–140. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Yao, X.; Tian, T.; Fu, X.; Wang, W.; Li, S.; Shi, T.; Suo, A.; Ruan, Z.; Guo, H.; et al. ABCB5-ZEB1 Axis Promotes Invasion and Metastasis in Breast Cancer Cells. Oncol. Res. 2017, 25, 305–316. [Google Scholar] [CrossRef]
- Touil, Y.; Zuliani, T.; Wolowczuk, I.; Kuranda, K.; Prochazkova, J.; Andrieux, J.; Roy, H.L.; Mortier, L.; Vandomme, J.; Jouy, N.; et al. The PI3K/AKT signaling pathway controls the quiescence of the low-rhodamine123-retention cell compartment enriched for melanoma stem cell activity. Stem Cells 2013, 31, 641–651. [Google Scholar] [CrossRef]
- Ho, H.; Kapadia, R.; Al-Tahan, S.; Ahmad, S.; Ganesan, A.K. WIPI1 Coordinates Melanogenic Gene Transcription and Melanosome Formation via TORC1 Inhibition. JBC 2011, 286, 12509–12523. [Google Scholar] [CrossRef] [PubMed]
- Metri, R.; Mohan, A.; Nsengimana, J.; Pozniak, J.; Molina-Paris, C.; Newton-Bishop, J.; Bishop, D.; Chandra, N. Identification of a gene signature for discriminating metastatic from primary melanoma using a molecular interaction network approach. Sci. Rep. 2017, 7, 17314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Su, L.; Lei, Y.; Liu, X.; Zhang, Y.; Liu, X. DDIT3 and KAT2A Proteins Regulate TNFRSF10A and TNFRSF10B Expression in Endoplasmic Reticulum Stress-mediated Apoptosis in Human Lung Cancer Cells. JBC 2015, 290, 11108–11118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerezo, M.; Lehraiki, A.; Millet, A.; Rouaud, F.; Plaisant, M.; Jaune, E.; Botton, T.; Ronco, C.; Abbe, P.; Amdouni, H.; et al. Compounds Triggering ER Stress Exert Anti-Melanoma Effects and Overcome BRAF Inhibitor Resistance. Cancer Cell 2016, 29, 805–819. [Google Scholar] [CrossRef] [PubMed]
- Murugaesu, N.; Iravani, M.; van Weverwijk, A.; Ivetic, A.; Johnson, D.A.; Antonopoulos, A.; Fearns, A.; Jamal-Hanjani, M.; Sims, D.; Fenwick, K.; et al. An In Vivo Functional Screen Identifies ST6GalNAc2 Sialyltransferase as a Breast Cancer Metastasis Suppressor. Cancer Discov. 2014, 4, 304–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erler, J.T.; Giaccia, A.J. Lysyl Oxidase Mediates Hypoxic Control of Metastasis. Cancer Research 2006, 66, 10238–10241. [Google Scholar] [CrossRef] [PubMed]
- Avery-Kiejda, K.A.; Bowden, N.A.; Croft, A.J.; Scurr, L.L.; Kairupan, C.F.; Ashton, K.A.; Talseth-Palmer, B.A.; Rizos, H.; Zhang, X.D.; Scott, R.J.; et al. P53 in human melanoma fails to regulate target genes associated with apoptosis and the cell cycle and may contribute to proliferation. BMC Cancer 2011, 11, 203. [Google Scholar] [CrossRef]
- Aida, S.; Sonobe, Y.; Yuhki, M.; Sakata, K.; Fujii, T.; Sakamoto, H.; Mizuno, T. MITF suppression by CH5552074 inhibits cell growth in melanoma cells. Cancer Chemother. Pharmacol. 2017, 79, 1187–1193. [Google Scholar] [CrossRef]
- Zois, C.E.; Harris, A.L. Glycogen metabolism has a key role in the cancer microenvironment and provides new targets for cancer therapy. J. Mol. Med. 2016, 94, 137–154. [Google Scholar] [CrossRef] [Green Version]
- Mandruzzato, S.; Callegaro, A.; Turcatel, G.; Francescato, S.; Montesco, M.C.; Chiarion-Sileni, V.; Mocellin, S.; Rossi, C.R.; Bicciato, S.; Wang, E.; et al. A gene expression signature associated with survival in metastatic melanoma. J. Transl. Med. 2006, 4, 50. [Google Scholar] [CrossRef]
- Yang, L.; Moss, T.; Mangala, L.S.; Marini, J.; Zhao, H.; Wahlig, S.; Armaiz-Pena, G.; Jiang, D.; Achreja, A.; Win, J.; et al. Metabolic shifts toward glutamine regulate tumor growth, invasion and bioenergetics in ovarian cancer. Mol. Syst. Biol. 2014, 10, 728. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.-M.; Wei, W.; Sun, Y. Role of SKP1-CUL1-F-Box-Protein (SCF) E3 Ubiquitin Ligases in Skin Cancer. J. Genet. Genomics. 2013, 40. [Google Scholar] [CrossRef] [PubMed]
- Nagano, K.; Imai, S.; Zhao, X.; Yamashita, T.; Yoshioka, Y.; Abe, Y.; Mukai, Y.; Kamada, H.; Nakagawa, S.; Tsutsumi, Y.; et al. Identification and evaluation of metastasis-related proteins, oxysterol binding protein-like 5 and calumenin, in lung tumors. Int. J. Oncol. 2015, 47, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Welinder, C.; Jönsson, G.B.; Ingvar, C.; Lundgren, L.; Baldetorp, B.; Olsson, H.; Breslin, T.; Rezeli, M.; Jansson, B.; Fehniger, T.E.; et al. Analysis of Alpha-Synuclein in Malignant Melanoma—Development of a SRM Quantification Assay. PLOS ONE 2014, 9, e110804. [Google Scholar] [CrossRef] [PubMed]
- Woischke, C.; Blaj, C.; Schmidt, E.M.; Lamprecht, S.; Engel, J.; Hermeking, H.; Kirchner, T.; Horst, D. CYB5R1 links epithelial-mesenchymal transition and poor prognosis in colorectal cancer. Oncotarget 2016, 7, 31350–31360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, C.M.; Biran, A.; Burney, M.J.; Patel, H.; Henser-Brownhill, T.; Cohen, A.-H.S.; Li, Y.; Ben-Hamo, R.; Nye, E.; Spencer-Dene, B.; et al. The linker histone H1.0 generates epigenetic and functional intratumor heterogeneity. Science 2016, 353. [Google Scholar] [CrossRef] [PubMed]
- Vandereyken, M.; Jacques, S.; Van Overmeire, E.; Amand, M.; Rocks, N.; Delierneux, C.; Singh, P.; Singh, M.; Ghuysen, C.; Wathieu, C.; et al. Dusp3 deletion in mice promotes experimental lung tumour metastasis in a macrophage dependent manner. PLoS ONE 2017, 12, e0185786. [Google Scholar] [CrossRef]
- Qiu, J.; Peng, S.; Si-Tu, J.; Hu, C.; Huang, W.; Mao, Y.; Qiu, W.; Li, K.; Wang, D. Identification of endonuclease domain-containing 1 as a novel tumor suppressor in prostate cancer. BMC Cancer 2017, 17, 360. [Google Scholar]
- Vazquez, F.; Lim, J.-H.; Chim, H.; Bhalla, K.; Girnun, G.; Pierce, K.; Clish, C.B.; Granter, S.R.; Widlund, H.R.; Spiegelman, B.M.; et al. PGC1α Expression Defines a Subset of Human Melanoma Tumors with Increased Mitochondrial Capacity and Resistance to Oxidative Stress. Cancer Cell 2013, 23, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. PNAS 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database Hallmark Gene Set Collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | n | % of Total |
|---|---|---|
| Total patients enrolled | 39 | |
| Age at enrolment (years) | ||
| Median (60) | ||
| Range (30–84) | ||
| Gender | ||
| Male | 16 | 59% |
| Female | 23 | 41% |
| Stage of disease at baseline | ||
| Stage IV | ||
| M1a | 5 | 13% |
| M1b | 5 | 13% |
| M1c | 19 | 49% |
| M1d | 10 | 25% |
| Mutational status of tumour | ||
| BRAF Mut | 26 | 67% |
| V600E * | 20 | 51% |
| V600K | 5 | 13% |
| V600R | 1 | 3% |
| NRAS Mut | 3 | 7% |
| Q61K | 2 | 5% |
| Q61L | 1 | 2% |
| BRAF/NRAS WT | 10 | 26% |
| Gene Set Name (Hallmark) | Number of Genes in Overlap | k/K | p-Value | FDR q-Value | Overlapped Genes |
|---|---|---|---|---|---|
| TNFA_SIGNALING_VIA_NFKB | 20 | 0.1 | 3 × 10−21 | 1.5 × 10−19 | IL1B, NAMPT, NFKBIA, TNFAIP6, ABCA1, TNFAIP3, G0S2, TRIB1, NFIL3, SLC2A3, IRS2, SOCS3, GADD45B, SAT1, PLK2, BCL6, NR4A2, CCNL1, BCL2A1, B4GALT5 |
| INFLAMMATORY_RESPONSE | 16 | 0.08 | 1.1 × 10−15 | 2.8 × 10−14 | IL1B, NAMPT, NFKBIA, TNFAIP6, ABCA1, RAF1, ADM, HIF1A, CSF3R, AQP9, IL18RAP, MEFV, SLC31A2, FAR2, KCNJ2, PROK2 |
| KRAS_SIGNALING_UP | 9 | 0.045 | 3.3 × 10−7 | 5.5 × 10−6 | IL1B, TNFAIP3, G0S2, TRIB1, CSF2RA, ARG1, LY96, CLEC4A, TLR8 |
| COMPLEMENT | 8 | 0.04 | 3.6 × 10−6 | 3.0 × 10−5 | TNFAIP3, RAF1, PLSCR1, CDA, CR1, F5, GCA, ITGAM |
| HYPOXIA | 8 | 0.04 | 3.6 × 10−6 | 3.0 × 10−5 | TNFAIP3, NFIL3, SLC2A3, ADM, IRS2, DDIT3, EXT1, FOXO3 |
| INTERFERON_GAMMA_RESPONSE | 8 | 0.04 | 3.6 × 10−6 | 3.0 × 10−5 | NAMPT, NFKBIA, TNFAIP6, TNFAIP3, HIF1A, EIF4E3, PLSCR1, SOCS3 |
| IL2_STAT5_SIGNALING | 7 | 0.035 | 3.5 × 10−5 | 2.2 × 10−4 | NFIL3, SLC2A3, GADD45B, PLSCR1, IL1R2, NFKBIZ, WLS |
| MTORC1_SIGNALING | 7 | 0.035 | 3.5 × 10−5 | 2.2 × 10−4 | NAMPT, NFIL3, SLC2A3, DDIT3, IDI1, FAM129A, IFRD1 |
| IL6_JAK_STAT3_SIGNALING | 5 | 0.0575 | 4.6 × 10−5 | 2.6 × 10−4 | IL1B, SOCS3, CSF3R, CSF2RA, IL1R2 |
| BILE_ACID_METABOLISM | 5 | 0.0446 | 1.5 × 10−4 | 7.7 × 10−4 | ABCA1, AQP9, IDI1, ACSL1, SULT1B1 |
| Gene Set Name (Hallmark) | Number of Genes in Overlap | k/K | p-Value | FDR q-Value | Overlapped Genes |
|---|---|---|---|---|---|
| EPITHELIAL_MESENCHYMAL_TRANSITION | 8 | 0.04 | 4.7 × 10−7 | 2.3 × 10−5 | VCAN, MYL9, PLOD2, LRP1, THBS1, FGF2, PTX3, CALU |
| APICAL_JUNCTION | 7 | 0.035 | 6 × 10−6 | 1 × 10−6 | VCAN, MYL9, INSIG1, ADAM9, SRC, VCL, RRAS |
| HEME_METABOLISM | 7 | 0.035 | 6 × 10−6 | 1 × 10−6 | H1F0, ENDOD1, BLVRA, CTNS, HEBP1, MARCH2, SNCA |
| ANDROGEN_RESPONSE | 4 | 0.04 | 4 × 10−4 | 4.5 × 10−3 | INSIG1, H1F0, GUCY1A3, ABCC4 |
| ESTROGEN_RESPONSE_EARLY | 5 | 0.025 | 6.3 × 10−4 | 4.5 × 10−3 | ENDOD1, KLF10, MYOF, AMFR, ZNF185 |
| KRAS_SIGNALING_UP0 | 5 | 0.025 | 6.3 × 10−4 | 4.5 × 10−3 | GUCY1A3, TSPAN13, YRDC, EPB41L3, MMD |
| MTORC1_SIGNALING | 5 | 0.025 | 6.3 × 10−4 | 4.5 × 10−3 | PLOD2, INSIG1, DDIT3, STARD4, MLLT11 |
| COAGULATION | 4 | 0.029 | 1.3 × 10−3 | 8.1 × 10−3 | LRP1, THBS1, ADAM9, SERPING1 |
| COMPLEMENT | 4 | 0.02 | 4.9 × 10−3 | 2.7 × 10−2 | LRP1, ADAM9, SRC, SERPING1 |
| PI3K_AKT_MTOR_SIGNALING | 3 | 0.029 | 5.6 × 10−3 | 2.7 × 10−9 | DDIT3, DUSP3, MAP2K6 |
| CTC Fraction | Gene Symbols |
|---|---|
| MCSP CTC | ST6GALNAC2, DDIT3, PLK2, LOXL4, CYSTM1, WIPI1, PYGL, GLUL, SAT1, SKP1, SEC31A |
| ABCB5 CTC | CYB5R1, H1F0, DUSP3, DDIT3, CREG1, ENDOD1, CALU, WIPI1, TXN2, SNCA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aya-Bonilla, C.; Gray, E.S.; Manikandan, J.; Freeman, J.B.; Zaenker, P.; Reid, A.L.; Khattak, M.A.; Frank, M.H.; Millward, M.; Ziman, M. Immunomagnetic-Enriched Subpopulations of Melanoma Circulating Tumour Cells (CTCs) Exhibit Distinct Transcriptome Profiles. Cancers 2019, 11, 157. https://doi.org/10.3390/cancers11020157
Aya-Bonilla C, Gray ES, Manikandan J, Freeman JB, Zaenker P, Reid AL, Khattak MA, Frank MH, Millward M, Ziman M. Immunomagnetic-Enriched Subpopulations of Melanoma Circulating Tumour Cells (CTCs) Exhibit Distinct Transcriptome Profiles. Cancers. 2019; 11(2):157. https://doi.org/10.3390/cancers11020157
Chicago/Turabian StyleAya-Bonilla, Carlos, Elin S. Gray, Jayapal Manikandan, James B. Freeman, Pauline Zaenker, Anna L. Reid, Muhammad A. Khattak, Markus H. Frank, Michael Millward, and Mel Ziman. 2019. "Immunomagnetic-Enriched Subpopulations of Melanoma Circulating Tumour Cells (CTCs) Exhibit Distinct Transcriptome Profiles" Cancers 11, no. 2: 157. https://doi.org/10.3390/cancers11020157
APA StyleAya-Bonilla, C., Gray, E. S., Manikandan, J., Freeman, J. B., Zaenker, P., Reid, A. L., Khattak, M. A., Frank, M. H., Millward, M., & Ziman, M. (2019). Immunomagnetic-Enriched Subpopulations of Melanoma Circulating Tumour Cells (CTCs) Exhibit Distinct Transcriptome Profiles. Cancers, 11(2), 157. https://doi.org/10.3390/cancers11020157