Head and Body/Tail Pancreatic Carcinomas Are Not the Same Tumors


Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Expression Data Sets
2.2. Preanalytic Gene Expression Data Processing
2.3. Gene Expression Data Analysis
2.4. Statistical Analysis
3. Results
3.1. Patients’ Characteristics
3.2. Clinicopathological and Molecular Characteristics According to Anatomic Location
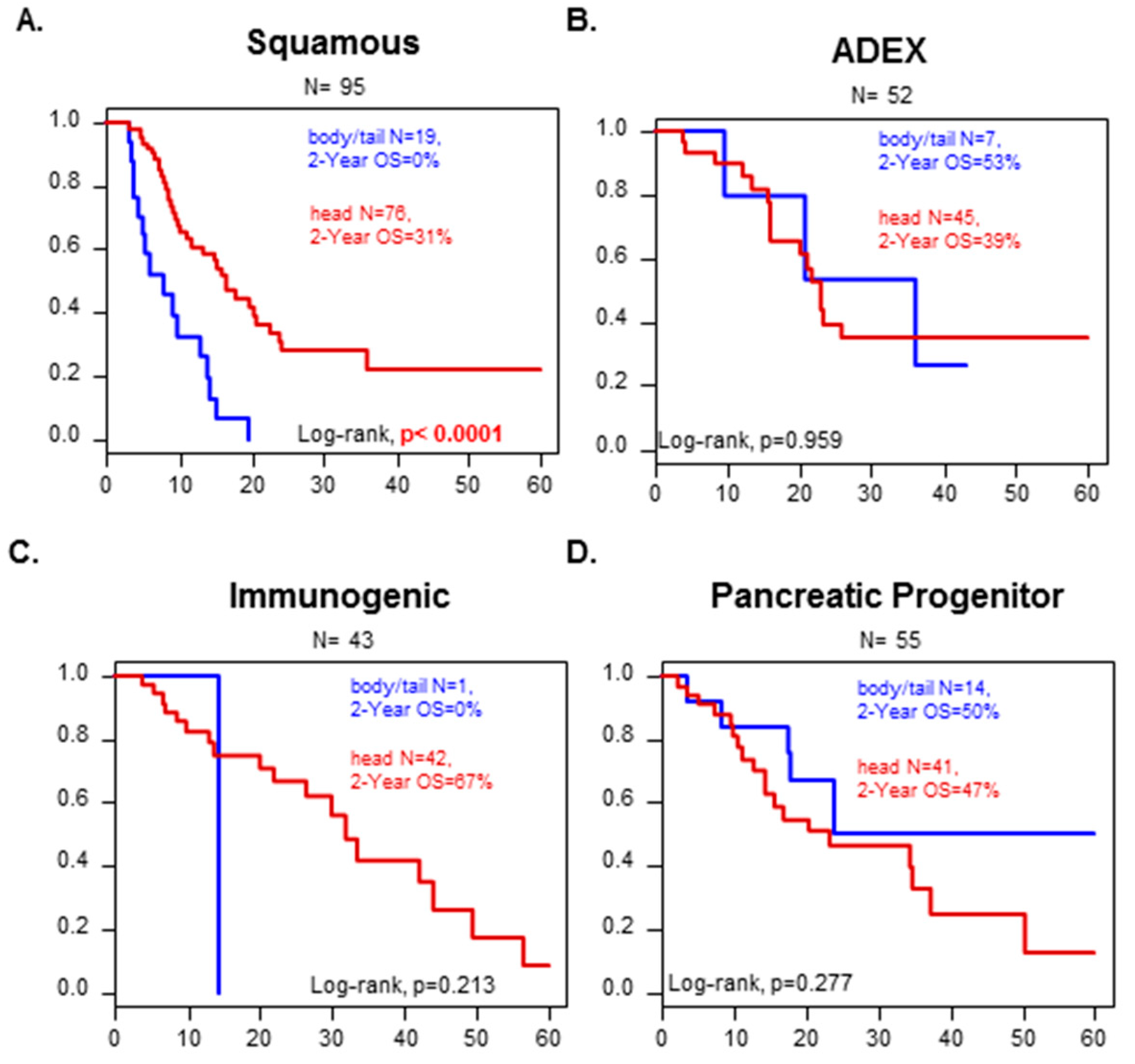
3.3. OS According to Anatomic Location and Prognostic Analysis
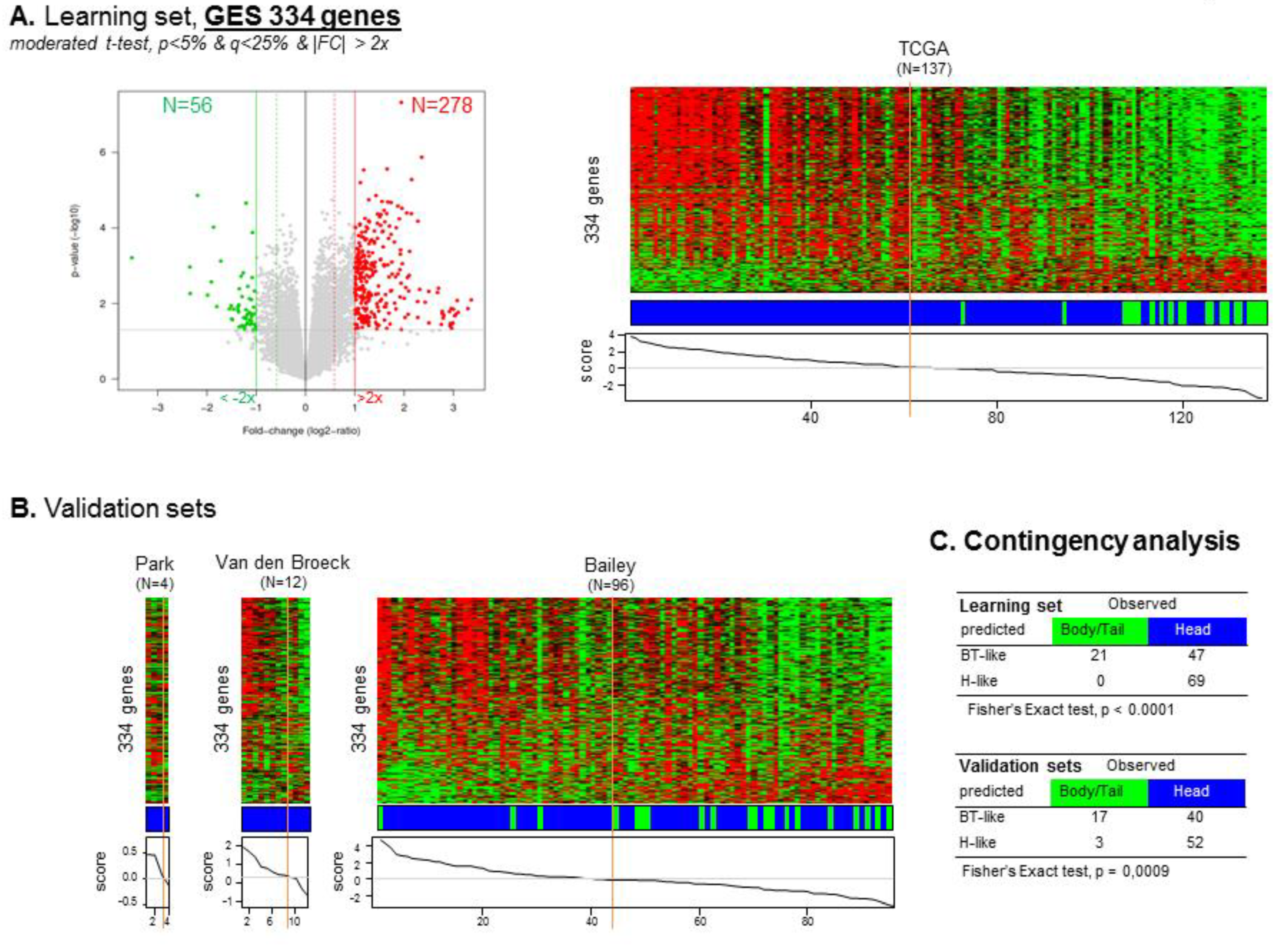
3.4. Head versus Body/Tail Gene Expression Signature
3.5. Correlation between “Head” vs. “Body/Tail” Locations and Immune and Stromal Signatures
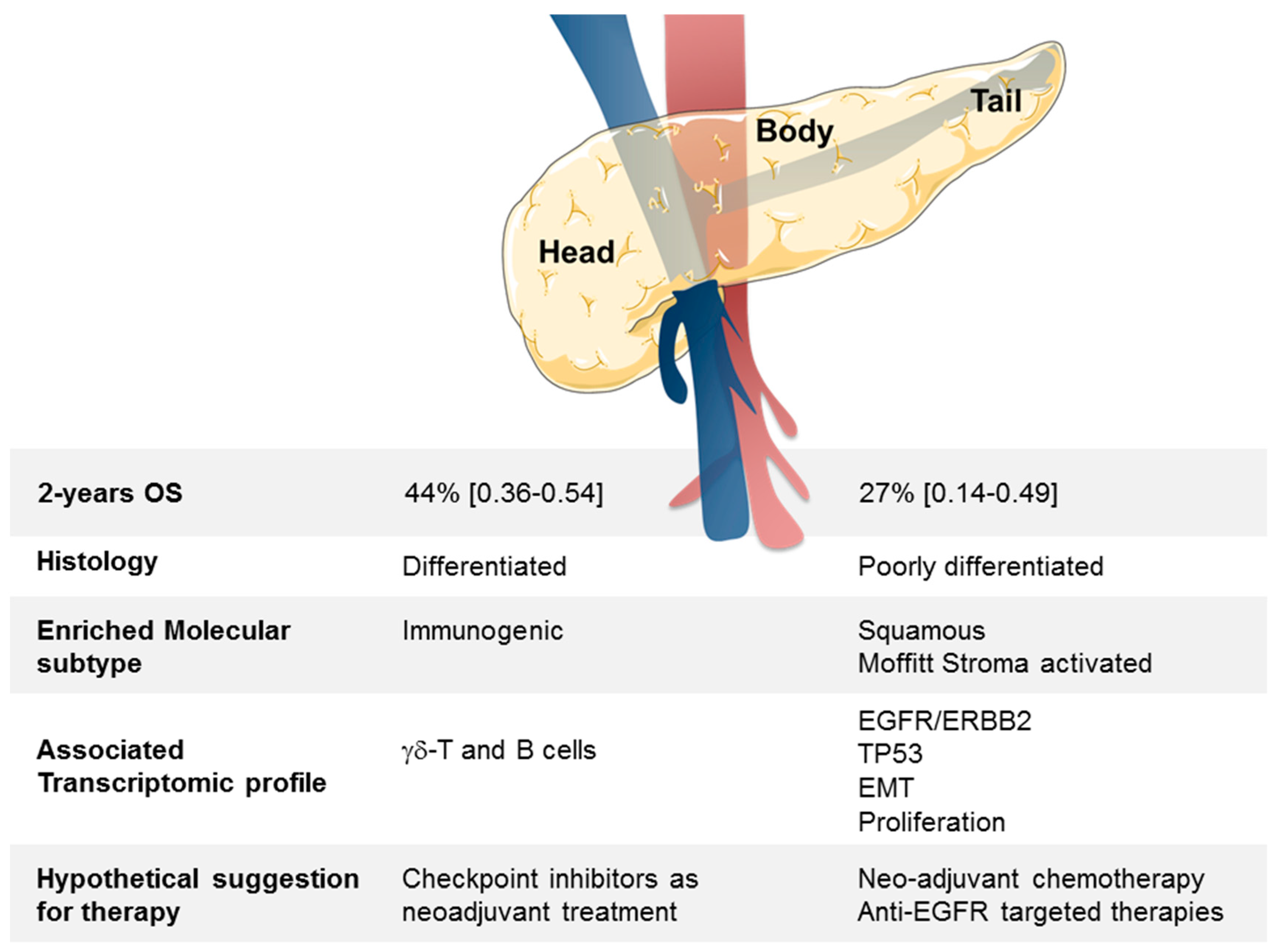
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting cancer incidence and deaths to 2030: The unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef]
- Ducreux, M.; Cuhna, A.S.; Caramella, C.; Hollebecque, A.; Burtin, P.; Goere, D.; Seufferlein, T.; Haustermans, K.; Van Laethem, J.L.; Conroy, T.; et al. Cancer of the pancreas: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2015, 26, v56–v68. [Google Scholar] [CrossRef] [PubMed]
- Neoptolemos, J.P.; Palmer, D.H.; Ghaneh, P.; Psarelli, E.E.; Valle, J.W.; Halloran, C.M.; Faluyi, O.; O’Reilly, D.A.; Cunningham, D.; Wadsley, J.; et al. Comparison of adjuvant gemcitabine and capecitabine with gemcitabine monotherapy in patients with resected pancreatic cancer (ESPAC-4): A multicentre, open-label, randomised, phase 3 trial. Lancet 2017, 389, 1011–1024. [Google Scholar] [CrossRef]
- Nowak, N.J.; Gaile, D.; Conroy, J.M.; McQuaid, D.; Cowell, J.; Carter, R.; Goggins, M.G.; Hruban, R.H.; Maitra, A. Genome-wide aberrations in pancreatic adenocarcinoma. Cancer Genet. Cytogenet. 2005, 161, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Buchler, M.; Friess, H.; Schultheiss, K.H.; Gebhardt, C.; Kubel, R.; Muhrer, K.H.; Winkelmann, M.; Wagener, T.; Klapdor, R.; Kaul, M.; et al. A randomized controlled trial of adjuvant immunotherapy (murine monoclonal antibody 494/32) in resectable pancreatic cancer. Cancer 1991, 68, 1507–1512. [Google Scholar] [CrossRef]
- Friess, H.; Buchler, M.; Kruger, M.; Beger, H.G. Treatment of duct carcinoma of the pancreas with the LH-RH analogue buserelin. Pancreas 1992, 7, 516–521. [Google Scholar]
- Gidekel Friedlander, S.Y.; Chu, G.C.; Snyder, E.L.; Girnius, N.; Dibelius, G.; Crowley, D.; Vasile, E.; DePinho, R.A.; Jacks, T. Context-dependent transformation of adult pancreatic cells by oncogenic K-Ras. Cancer Cell 2009, 16, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Hori, M.; Kitahashi, T.; Imai, T.; Ishigamori, R.; Takasu, S.; Mutoh, M.; Sugimura, T.; Wakabayashi, K.; Takahashi, M. Enhancement of carcinogenesis and fatty infiltration in the pancreas in N-nitrosobis(2-oxopropyl)amine-treated hamsters by high-fat diet. Pancreas 2011, 40, 1234–1240. [Google Scholar] [CrossRef]
- Kawamoto, S.; Siegelman, S.S.; Bluemke, D.A.; Hruban, R.H.; Fishman, E.K. Focal fatty infiltration in the head of the pancreas: Evaluation with multidetector computed tomography with multiplanar reformation imaging. J. Comput. Assist. Tomogr. 2009, 33, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Pitman, M.B.; Lewandrowski, K.; Shen, J.; Sahani, D.; Brugge, W.; Fernandez-del Castillo, C. Pancreatic cysts: Preoperative diagnosis and clinical management. Cancer Cytopathol. 2010, 118, 1–13. [Google Scholar] [CrossRef]
- Khashab, M.A.; Shin, E.J.; Amateau, S.; Canto, M.I.; Hruban, R.H.; Fishman, E.K.; Cameron, J.L.; Edil, B.H.; Wolfgang, C.L.; Schulick, R.D.; et al. Tumor size and location correlate with behavior of pancreatic serous cystic neoplasms. Am. J. Gastroenterol. 2011, 106, 1521–1526. [Google Scholar] [CrossRef]
- Park, J.; Lee, K.T.; Jang, T.H.; Seo, Y.W.; Lee, K.H.; Lee, J.K.; Jang, K.T.; Heo, J.S.; Choi, S.H.; Choi, D.W.; et al. Risk factors associated with the postoperative recurrence of intraductal papillary mucinous neoplasms of the pancreas. Pancreas 2011, 40, 46–51. [Google Scholar] [CrossRef]
- Oh, T.G.; Chung, M.J.; Park, J.Y.; Bang, S.M.; Park, S.W.; Chung, J.B.; Song, S.Y. Prognostic factors and characteristics of pancreatic neuroendocrine tumors: Single center experience. Yonsei Med. J. 2012, 53, 944–951. [Google Scholar] [CrossRef] [PubMed]
- Ruess, D.A.; Makowiec, F.; Chikhladze, S.; Sick, O.; Riediger, H.; Hopt, U.T.; Wittel, U.A. The prognostic influence of intrapancreatic tumor location on survival after resection of pancreatic ductal adenocarcinoma. BMC Surg. 2015, 15, 123. [Google Scholar] [CrossRef]
- Van Erning, F.N.; Mackay, T.M.; van der Geest, L.G.M.; Groot Koerkamp, B.; van Laarhoven, H.W.M.; Bonsing, B.A.; Wilmink, J.W.; van Santvoort, H.C.; de Vos-Geelen, J.; van Eijck, C.H.J.; et al. Association of the location of pancreatic ductal adenocarcinoma (head, body, tail) with tumor stage, treatment, and survival: A population-based analysis. Acta Oncol. 2018, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.T.; Schellenberg, D.; Shen, J.; Kim, J.; Goodman, K.A.; Fisher, G.A.; Ford, J.M.; Desser, T.; Quon, A.; Koong, A.C. Stereotactic radiotherapy for unresectable adenocarcinoma of the pancreas. Cancer 2009, 115, 665–672. [Google Scholar] [CrossRef]
- Marechal, R.; Demols, A.; Gay, F.; De Maertelaere, V.; Arvanitaki, M.; Hendlisz, A.; Van Laethem, J.L. Prognostic factors and prognostic index for chemonaive and gemcitabine-refractory patients with advanced pancreatic cancer. Oncology 2007, 73, 41–51. [Google Scholar] [PubMed]
- Tanaka, T.; Ikeda, M.; Okusaka, T.; Ueno, H.; Morizane, C.; Hagihara, A.; Iwasa, S.; Kojima, Y. Prognostic factors in japanese patients with advanced pancreatic cancer treated with single-agent gemcitabine as first-line therapy. Jpn. J. Clin. Oncol. 2008, 38, 755–761. [Google Scholar] [CrossRef]
- Lau, M.K.; Davila, J.A.; Shaib, Y.H. Incidence and survival of pancreatic head and body and tail cancers: A population-based study in the United States. Pancreas 2010, 39, 458–462. [Google Scholar] [CrossRef]
- Sener, S.F.; Fremgen, A.; Menck, H.R.; Winchester, D.P. Pancreatic cancer: A report of treatment and survival trends for 100,313 patients diagnosed from 1985–1995, using the National Cancer Database. J. Am. Coll. Surg. 1999, 189, 1–7. [Google Scholar] [CrossRef]
- Sohn, T.A.; Yeo, C.J.; Cameron, J.L.; Koniaris, L.; Kaushal, S.; Abrams, R.A.; Sauter, P.K.; Coleman, J.; Hruban, R.H.; Lillemoe, K.D. Resected adenocarcinoma of the pancreas-616 patients: Results, outcomes, and prognostic indicators. J. Gastrointest. Surg. 2000, 4, 567–579. [Google Scholar] [CrossRef]
- Toomey, P.; Hernandez, J.; Golkar, F.; Ross, S.; Luberice, K.; Rosemurgy, A. Pancreatic adenocarcinoma: Complete tumor extirpation improves survival benefit despite larger tumors for patients who undergo distal pancreatectomy and splenectomy. J. Gastrointest. Surg. 2012, 16, 376–381. [Google Scholar] [CrossRef]
- Dalton, R.R.; Sarr, M.G.; van Heerden, J.A.; Colby, T.V. Carcinoma of the body and tail of the pancreas: Is curative resection justified? Surgery 1992, 111, 489–494. [Google Scholar] [PubMed]
- Brennan, M.F.; Moccia, R.D.; Klimstra, D. Management of adenocarcinoma of the body and tail of the pancreas. Ann. Surg. 1996, 223, 506–511; discussion 511–512. [Google Scholar] [CrossRef] [PubMed]
- Sperti, C.; Pasquali, C.; Piccoli, A.; Pedrazzoli, S. Survival after resection for ductal adenocarcinoma of the pancreas. Br. J. Surg. 1996, 83, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Artinyan, A.; Soriano, P.A.; Prendergast, C.; Low, T.; Ellenhorn, J.D.; Kim, J. The anatomic location of pancreatic cancer is a prognostic factor for survival. HPB (Oxford) 2008, 10, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, S.B.; Jamieson, N.B.; Upstill-Goddard, R.; Bailey, P.J.; McKay, C.J.; Biankin, A.V.; Chang, D.K. Defining the molecular pathology of pancreatic body and tail adenocarcinoma. Br. J. Surg. 2018, 105, e183–e191. [Google Scholar] [CrossRef] [PubMed]
- Irizarry, R.A.; Bolstad, B.M.; Collin, F.; Cope, L.M.; Hobbs, B.; Speed, T.P. Summaries of Affymetrix GeneChip probe level data. Nucleic Acids Res. 2003, 31, e15. [Google Scholar] [CrossRef] [PubMed]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [PubMed]
- Collisson, E.A.; Sadanandam, A.; Olson, P.; Gibb, W.J.; Truitt, M.; Gu, S.; Cooc, J.; Weinkle, J.; Kim, G.E.; Jakkula, L.; et al. Subtypes of pancreatic ductal adenocarcinoma and their differing responses to therapy. Nat. Med. 2011, 17, 500–503. [Google Scholar] [CrossRef] [PubMed]
- Moffitt, R.A.; Marayati, R.; Flate, E.L.; Volmar, K.E.; Loeza, S.G.; Hoadley, K.A.; Rashid, N.U.; Williams, L.A.; Eaton, S.C.; Chung, A.H.; et al. Virtual microdissection identifies distinct tumor- and stroma-specific subtypes of pancreatic ductal adenocarcinoma. Nat. Genet. 2015, 47, 1168–1178. [Google Scholar] [CrossRef]
- Bailey, P.; Chang, D.K.; Nones, K.; Johns, A.L.; Patch, A.M.; Gingras, M.C.; Miller, D.K.; Christ, A.N.; Bruxner, T.J.; Quinn, M.C.; et al. Genomic analyses identify molecular subtypes of pancreatic cancer. Nature 2016, 531, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Gatza, M.L.; Lucas, J.E.; Barry, W.T.; Kim, J.W.; Wang, Q.; Crawford, M.D.; Datto, M.B.; Kelley, M.; Mathey-Prevot, B.; Potti, A.; et al. A pathway-based classification of human breast cancer. Proc. Natl. Acad. Sci. USA 2010, 107, 6994–6999. [Google Scholar] [CrossRef] [PubMed]
- Aran, D.; Hu, Z.; Butte, A.J. xCell: Digitally portraying the tissue cellular heterogeneity landscape. Genome Biol. 2017, 18, 220. [Google Scholar] [CrossRef]
- Smyth, G.K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Da Huang, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- McShane, L.M.; Altman, D.G.; Sauerbrei, W.; Taube, S.E.; Gion, M.; Clark, G.M. REporting recommendations for tumour MARKer prognostic studies (REMARK). Br. J. Cancer 2005, 93, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Gatza, M.L.; Kung, H.N.; Blackwell, K.L.; Dewhirst, M.W.; Marks, J.R.; Chi, J.T. Analysis of tumor environmental response and oncogenic pathway activation identifies distinct basal and luminal features in HER2-related breast tumor subtypes. Breast Cancer Res. 2011, 13, R62. [Google Scholar] [CrossRef]
- Watanabe, I.; Sasaki, S.; Konishi, M.; Nakagohri, T.; Inoue, K.; Oda, T.; Kinoshita, T. Onset symptoms and tumor locations as prognostic factors of pancreatic cancer. Pancreas 2004, 28, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Bilimoria, K.Y.; Bentrem, D.J.; Ko, C.Y.; Stewart, A.K.; Winchester, D.P.; Talamonti, M.S. National failure to operate on early stage pancreatic cancer. Ann. Surg. 2007, 246, 173–180. [Google Scholar] [CrossRef]
- Matsuno, S.; Egawa, S.; Fukuyama, S.; Motoi, F.; Sunamura, M.; Isaji, S.; Imaizumi, T.; Okada, S.; Kato, H.; Suda, K.; et al. Pancreatic Cancer Registry in Japan: 20 years of experience. Pancreas 2004, 28, 219–230. [Google Scholar] [CrossRef]
- Van Oost, F.J.; Luiten, E.J.; van de Poll-Franse, L.V.; Coebergh, J.W.; van den Eijnden-van Raaij, A.J. Outcome of surgical treatment of pancreatic, peri-ampullary and ampullary cancer diagnosed in the south of The Netherlands: A cancer registry based study. Eur. J. Surg. Oncol. 2006, 32, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.C.; Shao, Y.F.; Shan, Y.; Wu, J.X.; Zhao, P. Surgical effect of malignant tumor of body and tail of the pancreas: Compare with pancreatic head cancer. Zhonghua Wai Ke Za Zhi 2007, 45, 30–33. [Google Scholar] [PubMed]
- Notta, F.; Chan-Seng-Yue, M.; Lemire, M.; Li, Y.; Wilson, G.W.; Connor, A.A.; Denroche, R.E.; Liang, S.B.; Brown, A.M.; Kim, J.C.; et al. A renewed model of pancreatic cancer evolution based on genomic rearrangement patterns. Nature 2016, 538, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Zhang, X.; Parsons, D.W.; Lin, J.C.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Kamiyama, H.; Jimeno, A.; et al. Core signaling pathways in human pancreatic cancers revealed by global genomic analyses. Science 2008, 321, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- Talbott, V.A.; Yeo, C.J.; Brody, J.R.; Witkiewicz, A.K. Molecular profiling of synchronous and metachronous cancers of the pancreas reveal molecular mimicry between samples from the same patient. J. Surg. Res. 2012, 176, 154–158. [Google Scholar] [CrossRef]
- Eyigor, C.; Karaca, B.; Kuzeyli-Yildirim, Y.; Uslu, R.; Uyar, M.; Coker, A. Does the tumor localization in advanced pancreatic cancer have an influence on the management of symptoms and pain? J. Balk. Union Oncol. 2010, 15, 543–546. [Google Scholar]
- Jamal-Hanjani, M.; Wilson, G.A.; McGranahan, N.; Birkbak, N.J.; Watkins, T.B.K.; Veeriah, S.; Shafi, S.; Johnson, D.H.; Mitter, R.; Rosenthal, R.; et al. Tracking the Evolution of Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2017, 376, 2109–2121. [Google Scholar] [CrossRef]
- Al-Ghamdi, S.; Cachat, J.; Albasri, A.; Ahmed, M.; Jackson, D.; Zaitoun, A.; Guppy, N.; Otto, W.R.; Alison, M.R.; Kindle, K.B.; et al. C-terminal tensin-like gene functions as an oncogene and promotes cell motility in pancreatic cancer. Pancreas 2013, 42, 135–140. [Google Scholar] [CrossRef]
- Yin, X.; Wang, M.; Wang, H.; Deng, H.; He, T.; Tan, Y.; Zhu, Z.; Wu, Z.; Hu, S.; Li, Z. Evaluation of neurotensin receptor 1 as a potential imaging target in pancreatic ductal adenocarcinoma. Amino Acids 2017, 49, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Deb, T.B.; Zuo, A.H.; Barndt, R.J.; Sengupta, S.; Jankovic, R.; Johnson, M.D. Pnck overexpression in HER-2 gene-amplified breast cancer causes Trastuzumab resistance through a paradoxical PTEN-mediated process. Breast Cancer Res. Treat. 2015, 150, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Deb, T.B.; Zuo, A.H.; Wang, Y.; Barndt, R.J.; Cheema, A.K.; Sengupta, S.; Coticchia, C.M.; Johnson, M.D. Pnck induces ligand-independent EGFR degradation by probable perturbation of the Hsp90 chaperone complex. Am. J. Physiol. Cell Physiol. 2011, 300, C1139–C1154. [Google Scholar] [CrossRef]
- Deb, T.B.; Coticchia, C.M.; Barndt, R.; Zuo, H.; Dickson, R.B.; Johnson, M.D. Pregnancy-upregulated nonubiquitous calmodulin kinase induces ligand-independent EGFR degradation. Am. J. Physiol. Cell Physiol. 2008, 295, C365–C377. [Google Scholar] [CrossRef] [PubMed]
- Malka, D.; Hammel, P.; Sauvanet, A.; Rufat, P.; O’Toole, D.; Bardet, P.; Belghiti, J.; Bernades, P.; Ruszniewski, P.; Levy, P. Risk factors for diabetes mellitus in chronic pancreatitis. Gastroenterology 2000, 119, 1324–1332. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | N | All | Pancreas Cancer Site | p-Value | Statistic | |
|---|---|---|---|---|---|---|
| Body/Tail | Head | |||||
| Age at diagnosis, years | 0.198 | 0.56 | ||||
| ≤60 | 79 | 79 (32%) | 9 (22%) | 70 (34%) | (0.22–1.27) | |
| >60 | 170 | 170 (68%) | 32 (78%) | 138 (66%) | ||
| Sex | 0.733 | 0.86 | ||||
| Female | 117 | 117 (47%) | 18 (44%) | 99 (48%) | (0.41–1.78) | |
| Male | 132 | 132 (53%) | 23 (56%) | 109 (52%) | ||
| Pathological tumor size (pT) | 0.049 | |||||
| pT1 | 10 | 10 (5%) | 5 (16%) | 5 (3%) | ||
| pT2 | 24 | 24 (13%) | 4 (13%) | 20 (13%) | ||
| pT3 | 148 | 148 (79%) | 22 (71%) | 126 (81%) | ||
| pT4 | 5 | 5 (3%) | 0 (0%) | 5 (3%) | ||
| Pathological lymph node status (pN) | 0.001 | 3.3 | ||||
| Negative | 79 | 79 (33%) | 22 (56%) | 57 (28%) | (1.53–7.1) | |
| Positive | 162 | 162 (67%) | 17 (44%) | 145 (72%) | ||
| Metastases | 0.063 | 0.28 | ||||
| Negative | 239 | 239 (96%) | 37 (90%) | 202 (97%) | (0.06–1.4) | |
| Positive | 10 | 10 (4%) | 4 (10%) | 6 (3%) | ||
| AJCC stage | 0.048 | |||||
| 1 | 25 | 25 (10%) | 7 (17%) | 18 (9%) | ||
| 2 | 209 | 209 (84%) | 30 (73%) | 179 (86%) | ||
| 3 | 5 | 5 (2%) | 0 (0%) | 5 (2%) | ||
| 4 | 10 | 10 (4%) | 4 (10%) | 6 (3%) | ||
| Pathological grade | 0.808 | |||||
| 1 | 21 | 21 (9%) | 2 (5%) | 19 (9%) | ||
| 2 | 137 | 137 (57%) | 23 (57%) | 114 (57%) | ||
| 3 | 81 | 81 (34%) | 15 (38%) | 66 (33%) | ||
| 4 | 2 | 2 (1%) | 0 (0%) | 2 (1%) | ||
| Bailey subtypes | 0.006 | |||||
| ADEX | 52 | 52 (21%) | 7 (17%) | 45 (22%) | ||
| Immunogenic | 45 | 45 (18%) | 1 (2%) | 44 (21%) | ||
| Pancreatic progenitor | 56 | 56 (22%) | 14 (34%) | 42 (20%) | ||
| Squamous | 96 | 96 (39%) | 19 (46%) | 77 (37%) | ||
| Collisson subtypes | 0.065 | |||||
| Classical | 118 | 118 (47%) | 24 (59%) | 94 (45%) | ||
| Exocrine-like | 87 | 87 (35%) | 8 (20%) | 79 (38%) | ||
| Quasi-mesenchymal | 44 | 44 (18%) | 9 (22%) | 35 (17%) | ||
| Moffitt subtypes, “Tumor” | 0.395 | 1.4 | ||||
| Basal-like | 112 | 112 (45%) | 21 (51%) | 91 (44%) | (0.65–2.8) | |
| Classical | 137 | 137 (55%) | 20 (49%) | 117 (56%) | ||
| Moffitt subtypes, “Stroma” | 0.007 | 3.1 | ||||
| Activated | 154 | 154 (63%) | 32 (82%) | 122 (60%) | (1.26–8.72) | |
| Normal | 90 | 90 (37%) | 7 (18%) | 83 (40%) | ||
| Follow-up median, months (min-max) | 245 | 11.17 (0–156.4) | 9.63 (0.03–60.25) | 10.97 (0–156.4) | ||
| 2-Year OS (95% CI) | 245 | 41% (0.33–0.49) | 27% (0.14–0.49) | 44% (0.36–0.54) | 0.044 | |
| Characteristics | Univariate | Multivariate | |||||
|---|---|---|---|---|---|---|---|
| N | HR (95%CI) | p-Value | N | HR (95%CI) | p-Value | ||
| Age at diagnosis | >60 vs. ≤60 | 245 | 0.93 (0.63–1.37) | 0.708 | |||
| Sex | male vs. female | 245 | 1.13 (0.78–1.62) | 0.516 | |||
| Pathological type | other vs. ductal | 245 | 1.12 (0.62–1.99) | 0.711 | |||
| Pathological tumor size (pT) | 2 vs. 1 | 183 | 1.78 (0.37–8.66) | 0.290 | |||
| 3 vs. 1 | 2.49 (0.61–10.24) | ||||||
| 4 vs. 1 | 5.88 (0.81–42.62) | ||||||
| Pathological lymph node status (pN) | 1 vs. 0 | 237 | 1.77 (1.16–2.7) | 0.008 | 230 | 2.04 (1.28–3.27) | 0.003 |
| Metastases | 1 vs. 0 | 245 | 1.71 (0.74–3.92) | 0.207 | |||
| AJCC stage | 2 vs. 1 | 245 | 2.14 (0.99–4.63) | 0.083 | |||
| 3 vs. 1 | 4.98 (1.01–24.50) | ||||||
| 4 vs. 1 | 3.50 (1.16–10.61) | ||||||
| Pathological grade | 2 vs. 1 | 241 | 1.47 (0.59–3.69) | 0.056 | 230 | 1.38 (0.52–3.61) | 0.516 |
| 3 vs. 1 | 2.33 (0.92–5.89) | 230 | 1.86 (0.71–4.89) | 0.209 | |||
| 4 vs. 1 | 2.52 (0.49–13.13) | 230 | 3.61 (0.66–19.82) | 0.140 | |||
| Pancreatic tumor site | head vs. body/tail | 245 | 0.63 (0.41–0.99) | 0.045 | 230 | 0.46 (0.27–0.79) | 0.005 |
| KRAS mutation | MT vs. WT | 137 | 1.2 (0.68–2.13) | 0.534 | |||
| KRAS mutation, exon12 | G12D vs. WT | 137 | 1.53 (0.79–2.98) | 0.317 | |||
| G12R vs. WT | 1.03 (0.39–2.72) | ||||||
| G12V vs. WT | 0.63 (0.26–1.57) | ||||||
| Bailey subtypes | Immunogenic vs. ADEX | 245 | 0.92 (0.50–1.70) | 0.006 | 230 | 1.64 (0.75–3.6) | 0.216 |
| Pancreatic progenitor vs. ADEX | 0.97 (0.54–1.73) | 230 | 1.43 (0.68–3) | 0.345 | |||
| Squamous vs. ADEX | 1.87 (1.13–3.10) | 230 | 1.81 (0.8–4.1) | 0.152 | |||
| Collisson subtypes | Exocrine-like vs. Classical | 245 | 1.13 (0.74–1.70) | 0.131 | 230 | 1.75 (0.98–3.12) | 0.057 |
| Quasi-mesenchymal vs. Classical | 1.64 (1.01–2.66) | 230 | 1.12 (0.63–2.01) | 0.698 | |||
| Moffitt subtypes, “Tumor” | Classical vs. Basal-like | 245 | 0.53 (0.37–0.77) | 0.001 | 230 | 0.55 (0.31–1) | 0.050 |
| Moffitt subtypes, “Stroma” | Normal vs. Activated | 240 | 0.46 (0.3–0.69) | 0.0002 | 230 | 0.58 (0.36–0.93) | 0.023 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Birnbaum, D.J.; Bertucci, F.; Finetti, P.; Birnbaum, D.; Mamessier, E. Head and Body/Tail Pancreatic Carcinomas Are Not the Same Tumors. Cancers 2019, 11, 497. https://doi.org/10.3390/cancers11040497
Birnbaum DJ, Bertucci F, Finetti P, Birnbaum D, Mamessier E. Head and Body/Tail Pancreatic Carcinomas Are Not the Same Tumors. Cancers. 2019; 11(4):497. https://doi.org/10.3390/cancers11040497
Chicago/Turabian StyleBirnbaum, David Jérémie, François Bertucci, Pascal Finetti, Daniel Birnbaum, and Emilie Mamessier. 2019. "Head and Body/Tail Pancreatic Carcinomas Are Not the Same Tumors" Cancers 11, no. 4: 497. https://doi.org/10.3390/cancers11040497
APA StyleBirnbaum, D. J., Bertucci, F., Finetti, P., Birnbaum, D., & Mamessier, E. (2019). Head and Body/Tail Pancreatic Carcinomas Are Not the Same Tumors. Cancers, 11(4), 497. https://doi.org/10.3390/cancers11040497